


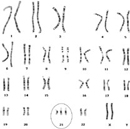
 Figure 2
Figure 2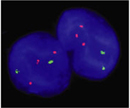















« Prev Next »
Aneuploidies disturb the delicate balance of gene products in cells. By definition, aneuploid cells have an abnormal number of chromosomes. Because each chromosome contains hundreds of genes, the addition or loss of even a single chromosome disrupts the existing equilibrium in cells, and in most cases, is not compatible with life.
Using the tools of modern cytogenetics, scientists have recently provided new insights into the origins of aneuploidy. Researchers now appreciate that aneuploid gametes are produced at surprisingly high rates in human meioses, and that very few aneuploid embryos are able to survive. Much attention is currently focused on determining how specific imbalances in gene expression lead to the profound phenotypes associated with aneuploid conditions, such as Down syndrome, with the ultimate goal of developing therapeutic interventions.
Most Aneuploidies Are Lethal
In humans, the most common aneuploidies are trisomies, which represent about 0.3% of all live births. Trisomies are characterized by the presence of one additional chromosome, bringing the total chromosome number to 47. With few exceptions, trisomies do not appear to be compatible with life. In fact, trisomies represent about 35% of spontaneous abortions (Figure 1; Hassold & Hunt, 2001).

Figure 1: Incidence of aneuploidy during development
The observed level of aneuploidy in humans varies depending on the developmental time point being examined.
© 2001 Nature Publishing Group Hassold, T. et al. To err (meiotically) is human: The genesis of human aneuploidy. Nature Reviews Genetics 2, 283. All rights reserved. 
Viable trisomies are restricted to only a few human chromosomes. The most common human trisomy involves chromosome 21 and is known as Down syndrome (DS), named for John Langdon Down, the physician who first described the condition in 1866. Interestingly, almost a century elapsed between the clinical description of DS and its association with trisomy 21. When Jerome Lejeune associated DS with trisomy 21 in 1959, human cytogenetics was still maturing as a science; thus, Lejeune's discovery represents a landmark in the field (Patterson & Costa, 2005). Joe Hin Tjio and Albert Levan had established the human chromosome number as 46 only a few years earlier, and chromosome-staining techniques were still under development at the time.
Today, DS (Figure 2) is routinely identified in karyotypes or by using fluorescent probes for chromosome 21 through fluorescence in situ hybridization (FISH) (Figure 3; Antonarakis et al., 2004). Down syndrome is a relatively common birth defect, affecting about 1 in every 750 births. Individuals with DS all demonstrate signs of cognitive impairment, along with a more variable range of other symptoms, but patients routinely live to adulthood. By contrast, trisomies of other autosomes appear to have more severe effects, because they are rarely observed in live births. Indeed, the only other autosomal trisomies that are detected in any appreciable numbers among newborns involve chromosomes 13 and 18, but affected babies rarely survive beyond the first few months of life (Hassold & Hunt, 2001).
Humans are much more able to tolerate extra sex chromosomes than extra autosomes. Thus, after DS, the most common human aneuploidy is the condition known as Klinefelter's syndrome (Jacobs & Strong, 1959). Klinefelter's males have a total chromosome number of 47, which includes two X chromosomes and one Y chromosome. According to convention, these males are designated as 47,XXY individuals. Other configurations of the sex chromosomes have been observed in 47,XXX females and 47,XYY males. Compared to autosomal trisomies, these sorts of sex chromosome trisomies are fairly benign. Affected individuals generally show reduced sexual development and fertility, but they often have normal life spans, and many of their symptoms can be treated by hormone supplementation. The ability of humans to tolerate supernumerary sex chromosomes is quite remarkable, as individuals can survive with as many as four sex chromosomes. This tolerance most likely relates to both X inactivation and to the small number of genes on the Y chromosome. In fact, when cells from individuals with more than one copy of the X chromosome are analyzed under a microscope, all but one of the X chromosomes appear as condensed Barr bodies, the cytological manifestations of X-chromosome inactivation. Supernumerary copies of the Y chromosome may be tolerated because the few gene products of the Y chromosome are not required for survival.
Monosomies are the opposite of trisomies, in that affected individuals are missing one chromosome, reducing their total chromosome number to 45. Cells seem to be particularly sensitive to the loss of a chromosome, because the only viable human monosomy involves the X chromosome. Females with a single copy of the X chromosome have the condition known as Turner's syndrome. Interestingly, the frequency of Turner's syndrome is significantly lower than that of sex chromosome trisomies, suggesting that a single X chromosome is insufficient for optimum cell function. The viable Turner's (45,X) females display a wide range of symptoms, which include infertility and impaired sexual development, and these individuals are usually mosaics. Most Aneuploidies Arise from Errors in Meiosis, Especially in Maternal Meiosis I
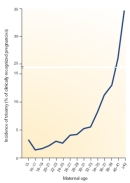
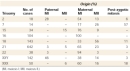
For some time, researchers have known that most aneuploidies result from the nondisjunction of chromosomes during meiosis. In recent years, scientists have been able to more precisely pinpoint the causes of aneuploidy by using polymorphic DNA markers to follow individual chromosomes. Large collections of polymorphic markers are now available for all of the human chromosomes, and investigators can use these markers to distinguish between maternally derived chromosomes and paternally derived chromosomes. Because we know how chromosomes segregate in meiosis, polymorphic DNA markers can also be used to determine whether an aneuploid gamete arose from an error in meiosis I or meiosis II. A recent analysis of trisomic fetuses and live births with these kinds of markers (Hassold & Hunt, 2001) revealed considerable variation with respect to both the parental origin of the extra chromosome and the meiotic division in which the error occurred (Table 1). The results showed that trisomies can arise from segregation errors in either parent and in either meiotic division, but the overwhelming number of trisomies arise from errors during maternal meiosis I. Intuitively, this observation makes sense when considering that human oocytes can be arrested in prophase I for several decades. The medical community is well aware that the risk of trisomy also increases sharply with maternal age, particularly as women near the end of their reproductive life span (Figure 4). Consequently, pregnant women over the age of 35 are routinely offered testing for fetal chromosome abnormalities. It is difficult to estimate the true frequency of human aneuploidies, because the most seriously affected embryos probably do not survive to the developmental stage at which they can be scored, and many pregnancies may terminate before they are diagnosed. Nonetheless, rough rates of aneuploidy can be calculated by analyzing chromosomes in gametes and preimplantation embryos, and then making corrections for undetected chromosome compositions that could be lethal. With these reservations, the results from several studies suggest that the frequency of human aneuploidy is at least 5%, although it is probably higher than 20%. Interestingly, the spectrum of chromosomal aneuploidies in gametes and preimplantation embryos is broader than that in more advanced pregnancies, suggesting that many aneuploidies have lethal effects during early development (Hassold & Hunt, 2001).
Widespread Upregulation of Chromosome 21 Genes Is Observed in Down Syndrome
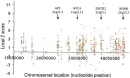
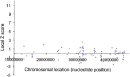
Scientists hypothesize that the severe phenotypes associated with aneuploidy are caused by imbalances in gene expression resulting from altered gene dosage (Antonarakis et al., 2004; FitzPatrick, 2005). In the absence of any mechanism for dosage compensation, the presence of an additional chromosome would be expected to bring about the simultaneous overexpression of hundreds of genes. In humans, the overexpression of X-linked genes is prevented by X inactivation, but no similar mechanism has been identified for autosomes. It is therefore interesting that trisomy 21 is the only viable autosomal trisomy, because the number of protein-coding sequences predicted for chromosome 21 is the smallest of any human chromosome, with the exception of the Y chromosome. Thus, an additional copy of chromosome 21 would be predicted to perturb the normal equilibrium in cells less than an extra copy of any other autosome. With the advent of DNA microarray technology, investigators are now able to measure the effects of trisomy 21 on the simultaneous expression of thousands of genes from multiple chromosomes. Using microarrays, several studies have documented widespread upregulation of chromosome 21 gene expression in brain samples obtained from aborted Down syndrome fetuses, compared to brain samples from unaffected fetuses of the same developmental age (Mao et al., 2003). In the data shown in Figure 5, a Z score is used to provide a rough estimate of the difference in gene expression between DS samples and normal samples. (Z scores are not simply ratios of expression levels in DS and normal tissues.) The Z scores are plotted for multiple genes along the length of chromosome 21. The positions of the genes involved in hereditary forms of Alzheimer's disease (APP) and amyotrophic lateral sclerosis (SOD1, superoxide dismutase) are also indicated. Colored symbols in the plot represent four different fetal DS brain samples and four astrocyte cell lines obtained from DS brain samples. The results show that the expression of many genes along the entire length of chromosome 21 is increased in DS. The increases observed for APP and SOD1 transcription are fairly typical of many genes, suggesting that their overexpression is not the major cause of DS symptoms. Importantly, control experiments (Figure 6) comparing two normal brain samples showed much lower levels of variation, indicating that the changes in gene expression observed in DS fetal brain samples were not due to random fluctuations associated with the microarray technique. Overall, the microarray results paint a complex picture of the molecular events that underlie DS.
An ongoing challenge for DS research is to sort through the complex set of transcriptional changes associated with DS to identify those genes whose expression is most closely linked to DS phenotypes. To this end, investigators have already constructed mouse models of chromosome 21 overexpression that reproduce some of the symptoms of DS (Antonarakis et al., 2004; FitzPatrick, 2005; Patterson & Costa, 2005). Researchers hope that these kinds of models will provide useful experimental systems for developing therapeutic interventions for this debilitating condition, which affects millions of people around the world.
References and Recommended Reading
Antonarakis, S. E., et al. Chromosome 21 and Down syndrome: From genomics to pathophysiology. Nature Reviews Genetics 5, 725–738 (2004) doi:10.1038/nrg1448 (link to article)
FitzPatrick, D. R. Transcriptional consequences of autosomal trisomy: Primary gene dosage with complex downstream effects. Trends in Genetics 21, 249–253 (2005) doi:10.1016/j.tig.2005.02.012
Hassold, T., & Hunt, P. To err (meiotically) is human: The genesis of human aneuploidy. Nature Reviews Genetics 2, 280–291 (2001) doi:10.1038/35066065 (link to article)
Jacobs, P. A., & Strong, J. A. A case of human intersexuality having a possible XXY sex-determining mechanism. Nature 183, 302–303 (1959) doi:10.1038/183302a0 (link to article)
Mao, M., et al. Global up-regulation of chromosome 21 gene expression in the developing Down syndrome brain. Genomics 81, 457–467 (2003) doi:10.1016/S0888-7543(03)00035-1
Patterson, D., & Costa, A. C. S. Down syndrome and genetics—A case of linked histories. Nature Reviews Genetics 6, 137–147 (2005) doi:10.1038/nrg1525 (link to article)










