


 Figure 1: Chromosome translocations.
Figure 1: Chromosome translocations.















« Prev Next »
Translocations generate novel chromosomes. In a translocation, a segment from one chromosome is transferred to a nonhomologous chromosome or to a new site on the same chromosome. Translocations place genes in new linkage relationships and generate chromosomes without normal pairing partners. When scientists compare the genomes of closely related species, they can see that translocations have occurred many times during the course of evolution. However, translocations that give an organism an adaptive advantage are very rare. Translocations are more often associated with negative consequences such as aneuploidy, infertility, or cancer. Scientists are now able to use molecular probes to analyze translocations, and the results are providing new insights into the molecular origins of various diseases, including cancer.
Translocations Generate Novel Chromosomes
Translocations were first detected cytologically in the late nineteenth and early twentieth centuries as novel chromosomes that appeared prominently in tumor cells. Some of the earliest, most complete descriptions of tumor cell chromosomes were provided by the great German cytologist Theodor Boveri, considered by many to be first cancer geneticist. Based on his observations of tumors, Boveri postulated that tumor cells possessed "growth-stimulatory chromosomes" that played a role in malignancy (Balmain, 2001). At the time, cytological markers were not available for human chromosomes, so Boveri was not able to identify specific chromosomal changes in tumors. Today, we know that Boveri's insights were correct. Translocations are indeed common in cancer cells, and some translocations produce oncogenes that are responsible for malignant transformation.
With the development of genetic and cytological models in the early twentieth century, the existence of translocations became firmly established. Geneticists had discovered that genes were physically linked on chromosomes, and that the strength of genetic linkage could be used to provide a rough map of each chromosome. Occasionally, however, geneticists discovered mutations in which genes from two different chromosomes behaved as if they were physically linked. Furthermore, the number of these mutations could be increased by orders of magnitude when organisms were treated with X-rays. Examination of these organisms' chromosomes provided an explanation for the appearance of novel linkage groups; specifically, new chromosomes had appeared, and existing chromosomes had been altered. Moreover, in organisms like Drosophila and maize, for which cytological markers were available, scientists observed that the novel chromosomes contained parts of normal chromosomes. Thus, they were able to infer that breaks had occurred in normal chromosomes, and that the broken ends of nonhomologous chromosomes had fused together, producing translocations.
These early observations provided the foundation for our modern understanding of translocations. We now appreciate that translocations require double-strand breaks (DSBs) in DNA at two locations. The frequency of these DSBs is greatly increased by ionizing radiation, which is still used experimentally to generate translocations. Cells are not tolerant of DSBs; these breaks cause cells to arrest in mitosis or to undergo apoptosis. Therefore, the appearance of DSBs activates the cellular DNA repair machinery that catalyzes the joining of broken chromosome ends (Lieber et al., 2003). A variety of rearrangements can result from this joining. For instance, precise joining of broken ends can regenerate a normal chromosome. Deletions, duplications, and inversions can occur when joining involves two broken ends on the same chromosome. Furthermore, translocations may occur when the broken ends of two nonhomologous chromosomes are joined together. Nonhomologous end joining is often imprecise, so some nucleotides may be lost altogether during the joining process.
Karyotypes Are Used to Classify Translocations
© 2002 Nature Publishing Group Braude, P. et al. Preimplantation genetic diagnosis. Nature Reviews Genetics 3:(12) 941-953, doi:10.1038/nrg953 (2002). All rights reserved. 
Translocations involving human chromosomes are of great clinical interest because they have been linked to a number of disorders, including mental retardation, infertility, and cancer. Translocations are usually detected when a cytogeneticist examines a karyotype, which is an ordered arrangement of an individual's metaphase chromosomes. In standard karyotypes, chromosomes that have been stained with Giemsa dye after a special treatment reveal a characteristic set of bands along their length. Translocations are thus manifested as a change in the length or the banding pattern of a chromosome arm. Over time, cytogeneticists have discovered hundreds, if not thousands, of different human translocations. To describe these rearrangements, cytogeneticists have developed a standardized nomenclature that allows investigators to narrow in on chromosomal regions that are affected by translocations.
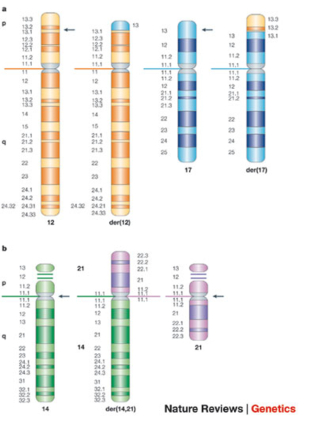
Nonreciprocal translocations are one-way translocations in which a chromosomal segment is transferred to a nonhomologous chromosome. Reciprocal translocations, on the other hand, involve the exchange of segments from two nonhomologous chromosomes. If no genetic material is lost during the exchange, the translocation is considered to be a balanced translocation. An example of a balanced reciprocal translocation is shown in the idiograms in Figure 1a, which depicts a reciprocal translocation that has occurred between chromosome 12 (orange) and chromosome 17 (blue). The arrows in the figure indicate the position of the breakpoints on each chromosome. This particular translocation would be designated as t(12;17)(p13.1;p13), where "t" indicates that a translocation has occurred. The first set of parentheses identifies the two chromosomes involved in the translocation, and the second set of parentheses indicates the breakpoints on the chromosome arms. Thus, this particular translocation involved region p13.1 of chromosome 12 and region p13 of chromosome 17. The rearranged chromosomes that result from a translocation are referred to as derivative (der) chromosomes. In this particular translocation, the rearranged chromosome is referred to as der(12) t(12;17), because the centromere of the derivative chromosome is derived from chromosome 12.
Yet another category of translocations is Robertsonian translocations (Figure 1b), in which the long q arms of two acrocentric chromosomes become joined at a single centromere. Chromosome p arms are lost during Robertsonian translocations, but because acrocentric chromosomes have very short p arms that are repetitive, there are no phenotypic consequences. Figure 1b depicts a Robertsonian translocation between chromosomes 14 and 21 that has generated the derivative chromosome der(14; 21). This particular translocation is interesting because it is commonly observed in patients with the familial form of Down syndrome. Familial Down syndrome is much less common than the form in which patients have 47 chromosomes due to the presence of an extra full copy of chromosome 21. Patients with familial Down syndrome have 46 chromosomes, including two normal copies of chromosome 21 and a Robertsonian translocation with chromosome 21 material. Familial Down patients usually inherit the translocation chromosome from an unaffected parent, who has only 45 chromosomes, including the Robertsonian chromosome and one normal copy of chromosome 21.
Investigators Use Multi-Color FISH to Scan for Translocations
Today, multi-color fluorescence in situ hybridization (FISH), or spectral karyotyping, is often used to screen patient samples for translocations because this method provides a sensitive genome-wide overview of an individual's chromosomes. Spectral karyotyping is particularly useful for cancer diagnoses, because transformed cells often have multiple chromosomal rearrangements. In spectral karyotyping, chromosomes are hybridized with a mixture of fluorescent DNA probes that generate a discrete spectral signal for each chromosome. When metaphase cells are hybridized with these probes, normal chromosomes appear uniformly colored along their lengths, while translocation chromosomes show more than one color along their lengths.
Figure 2 shows an example of a spectral karyotype prepared from acute myeloid leukemia cells with multiple translocations, which are indicated by arrows. The image obtained in the microscope (Figure 2a) was processed by a computer to generate a false-color image (Figure 2b) that depicts each chromosome in a distinctive color. The chromosomes were then arranged into a karyotype. Figure 2c shows portions of the karyotype in which translocation chromosomes were placed next to their normal homologues. At least 30 rearrangements were observed in this cell, and some of the chromosomes underwent multiple translocations.
Spectral karyotypes lack the spatial detail of Giemsa-stained karyotypes, so it is not possible to define the translocation breakpoints. Nonetheless, FISH is useful because it can reveal small translocations that would not be possible to detect with banding.
Translocation Carriers Face Reproductive Issues
Balanced translocations may be undetected in individuals who carry them as long as the total amount of genetic material has not been changed by the translocations and the translocations have not altered normal gene function. Problems arise during meiosis, however, because the translocation chromosomes are partially homologous to two different chromosomes. For example, Figure 3 diagrams the meiotic divisions in an individual who carries a balanced reciprocal translocation. During prophase I, a cross-like structure forms to allow synapsis and crossing over. Then, when the cells enter anaphase I, there are three possible ways that the chromosomes can segregate. (Each of the daughter cells must receive a "red" centromere and a "blue" centromere.) If the chromosomes separate by adjacent-1 or adjacent-2 segregation, the daughter cells receive two chromosomes that were "adjacent" to each other in the prophase configuration. These gametes are aneuploid, with an excess of some genes and deficiencies in others. Only alternate segregation produces euploid gametes, which contain either both translocation products or two normal chromosomes. Statistically, aneuploid gametes should outnumber euploid gametes, so translocation carriers have an elevated risk of having children with mental retardation and other birth defects linked to aneuploidy. If the aneuploidies are severe enough to cause embryonic lethality, translocation carriers may also appear to have problems with fertility.
Translocations Can Produce Oncogenes
© 2001 Nature Publishing Group Rowley, J. D. Chromosome translocations: dangerous liaisons revisited. Nature Reviews Cancer 1, 247 (2001). All rights reserved.
Depending on the chromosome breakpoints, a translocation can result in the disruption or misregulation of normal gene function. These molecular rearrangements, in many cases, are considered to be the primary cause of various cancers. Indeed, over the past few decades, clinical cytogeneticists have been able to link specific chromosome breakpoints to clinically defined cancers, including subtypes of leukemias, lymphomas, and sarcomas. Virtually all of the translocations observed in tumors have arisen through somatic mutations, so these are not inherited in families.
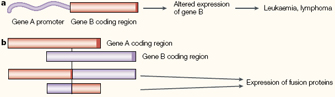
Analyses of the DNA sequences surrounding many of these translocation breakpoints have provided important mechanistic insights into cancer. Figure 4 outlines two kinds of molecular rearrangements that have frequently been linked to malignant transformation. In Figure 4a, the translocation places the coding sequence of one gene (Gene B) in proximity to the regulatory sequence for a different gene (Gene A). The first translocation of this kind to be described was a translocation involving chromosomes 8 and 14 in patients with Burkitt's lymphoma. This particular translocation places the MYC proto-oncogene from chromosome 8 under the control of the powerful immunoglobin heavy chain gene (IGH) promoter on chromosome 14. The MYC protein normally signals for cell proliferation, and the translocation causes high levels of MYC overexpression in lymphoid cells, where the IGH promoter is normally active.
In other cancers, translocations fuse the coding sequences of two genes together to generate potent oncogenes (Figure 4b). An example of historic interest is the Philadelphia chromosome, which was initially identified as a minute, or unusually small, chromosome in patients with chronic myelogenous leukemia (CML). With the advent of chromosome banding techniques, Janet Rowley was able to show that the Philadelphia chromosome was actually a product of a reciprocal translocation involving small segments at the ends of the q arms of chromosomes 9 and 22 (Rowley, 1973). Subsequent molecular analyses involving multiple laboratories revealed that the translocation fused the coding sequence of the BCR (breakpoint cluster region) gene on chromosome 22 with the coding sequence of the ABL gene on chromosome 9. The BCR-ABL fusion protein encoded by the chimeric gene is a protein tyrosine kinase that constitutively activates signaling pathways involved in cell growth and proliferation. Knowledge of this particular breakpoint has led to a successful treatment for CML, because investigators were able to use the sequence information to overexpress and crystallize the BCR-ABL protein, which in turn led to the development of drugs that inhibit this protein's activity.
Hundreds of translocations have now been linked to cancer, and this information has been catalogued and is freely available online to cytogeneticists. The database continues to grow as investigators uncover new links between genes, chromosomes, and cancer. It is interesting to consider that approximately one century has elapsed since Theodor Boveri predicted that growth-stimulatory chromosomes were present in cancer cells. Scientists continue to identify new molecular rearrangements that occur in cancer, with the hope that this information will translate into new treatments for a devastating disease.
References and Recommended Reading
Balmain, A. Cancer genetics: From Boveri and Mendel to microarrays. Nature Reviews Cancer 1, 77–82 (2001) doi:10.1038/35094086 (link to article)
Lieber, M. R., et al. Mechanism and regulation of human non-homologous end-joining. Nature Reviews Molecular Cell Biology 4, 712–720 (2003) doi:10.1038/nrm1202 (link to article)
Rowley, J. D. A new consistent chromosomal abnormality in chronic myelogeneous leukemia identified by quinacrine fluorescence and Giemsa staining. Nature 243, 290–293 (1973) doi:10.1038/243290a0 (link to article)
———. Chromosome translocations: Dangerous liaisons revisited. Nature Reviews Cancer 1, 245–250 (2001) doi:10.1038/35106108 (link to article)










