


 Figure 1: Drawing of mitosis by Walther Flemming.
Figure 1: Drawing of mitosis by Walther Flemming.














« Prev Next »
The second half of the nineteenth century was a time of remarkable advances in genetic research. As early as the 1860s, Gregor Mendel and Charles Darwin began to explore possible mechanisms of heredity. Then, over the next few decades, Walther Flemming, Theodor Boveri, and Walter Sutton made a series of significant discoveries involving chromosomes, including what these structures looked like, how they moved during mitosis, and what role they likely played in the transmission of genetic characteristics. However, it wasn't until the work of Thomas Hunt Morgan in the early twentieth century that researchers were finally able to directly link the inheritance of genetic traits to the behavior of chromosomes, thereby providing concrete evidence for what became known as the chromosome theory of heredity.
Early Breakthroughs: Mendel and Darwin Propose Mechanisms of Heredity
The year 1865 was marked by two profound biological breakthroughs: Mendel's publication of his experiments in plant hybridization, and Darwin's provisional hypothesis of pangenesis. Although neither man had any direct evidence for the theories he proposed, Mendel speculated that cells contained some type of factor that carried traits from one generation to the next, while Darwin proposed that traits could be passed down via units he termed "gemmules," which he believed traveled from every body part to the sexual organs, where they were stored (Benson, 2001). Because these first attempts to explain the mechanisms of heredity lacked any scientific support, their profound importance went unrecognized by the scientific community for decades. Nonetheless, Mendel and Darwin's work laid the foundation for formulating a testable, research-based theory of heredity.
Flemming, Boveri, and Sutton Connect Chromosomes to Heredity
Meanwhile, several independent advances-namely, discovery of chromosomes and their behavior during mitosis as described by , and discovery of the link between chromosomes and heredity as described by Theodor Boveri-began to shed light on the events that take place in the nucleus during cell division and their implications. Then, some thirty-five years later, the significance of Mendel's work was emphasized by Walter Sutton, whose observations of chromosome behavior during cell division and gamete formation were consistent with Mendel's findings. Thus, the basis for chromosome theory, and the field of cytogenetics, was created.
Walther Flemming Describes Chromosomes

Flemming, W. Zellsubstanz, Kern und Zelltheilung (F. C. W. Vogel, Leipzig, 1882).
By the middle of the nineteenth century, scientists understood that cells derived from other cells and that the hereditary information was located in the nucleus, but the physical nature of the hereditary material remained unknown. The microscopes of the time provided very poor resolution of living cellular structures, making it necessary for investigators to treat fixed cells with various stains to enhance the contrast of their contents. Thus, using innovative microscopy techniques and painstaking precision, German anatomist Walther Flemming recognized and explored the fibrous network within the nucleus, which he termed chromatin, or "stainable material." (Flemming had actually discovered the chromosome, although the term would be coined a few years later by Heinrich Waldeyer.) Flemming noted that during cell division, the chromatin formed threadlike bodies, which he termed mitosen, from the Greek word for thread. Based on many observations of cells in various stages of division (Figure 1), Flemming correctly deduced the sequence of chromosome movements during mitosis, which would be confirmed decades later by microscopy of live dividing cells. With his characteristic attention to detail, Flemming also made the important observation that chromosomes split along their length during mitosis, and he correctly hypothesized that the split chromosomes were partitioned into different daughter cells at the end of mitosis.
Thus, Flemming recognized that chromosomal movement during mitosis offered a mechanism for the precise distribution of nuclear material during cell division. His work provided an invaluable description of the initial mechanisms underlying the process of cell division, and it helped paved the way for the discovery of hereditary mechanisms.
Theodor Boveri Links Chromosomes and Heredity
The end of the nineteenth century was marked by advancements in cytological techniques and microscopy. During this period, German embryologist Theodor Boveri took Flemming's findings to the next level by providing the first evidence that chromosomes of germ cell lineage provide continuity between generations. Boveri found evidence for this hypothesis through his research of early development in the roundworm species Ascaris megalocephala (now known as Parascaris equorum). Ascaris embryos provided an excellent experimental model for Boveri's observations; this is because the large, clear cells of the Ascaris embryo have only two pairs of chromosomes, and the embryos develop distinct somatic cell and germ cell lineages during the first few cleavage divisions. Boveri was thus able to trace the fate of the chromosomes in individual cell lineages with great precision, and he made the surprising observation that the full complement of chromosomes was retained only in the Ascaris germ cell lineage, the source of future gametes. By contrast, the chromosomes in somatic cells, the source of all other adult tissues, underwent a curious process of fragmentation and elimination known as chromosome diminution. (Chromosome diminution does not occur in mammals.)
Boveri also recognized that chromosome number was reduced in the gametes. In Ascaris eggs, meiosis does not occur until fertilization is complete, so Boveri was able to observe the behavior of egg and sperm chromosomes following fertilization. He noted that Ascaris eggs retained only two chromosomes after the polar body formed, and that the normal number of four chromosomes was restored following fusion of the sperm and egg pronuclei. Boveri's work provided one of the first descriptions of meiosis.
Walter Sutton Finds Evidence for Mendel's Principles
Working independently near the dawn of the twentieth century, American graduate student Walter Sutton confirmed and expanded upon Boveri's observations using a superior cytological model. Sutton, a Kansas farm boy, had made the serendipitous discovery that it was possible to distinguish individual chromosomes in cells undergoing meiosis in the testes of the lubber grasshopper, Brachystola magna. Thus, in his classic paper published in 1902, Sutton described the configurations of individual chromosomes in cells at various stages of meiosis. Sutton was able to identify 11 pairs of chromosomes that could be distinguished by their sizes, as well as an accessory singleton that he correctly presumed to be a sex chromosome. Due to its apparent lack of a partner, this chromosome sorted into only half of the sperm cells, while the remaining half never received a copy. Though the accessory chromosome was unpaired, it still replicated and entered stages of mitosis in the same manner as all other chromosomes, which prompted Sutton to declare it a "true chromosome" and not simply an accessory.
Sutton postulated that all chromosomes have a stable structure, or "individuality," that is maintained between generations, and he used this property to follow the behavior of individual chromosomes through the various stages of meiosis, including synapsis. Most notably, Sutton recognized that his observations were consistent with the , whose findings had only recently been rediscovered. In fact, Sutton closed his 1902 paper with the statement, "I may finally call attention to the probability that the association of paternal and maternal chromosomes in pairs and their subsequent separation during the reducing division as indicated above may constitute the physical basis of the Mendelian law of heredity." With these words, Sutton articulated the chromosomal theory of inheritance.
Sutton subsequently went on to explain the basis for the ongoing variation among heritable traits. He noted that the position of each chromosome at the midline during metaphase was random, and that there was never a consistent maternal or paternal side of the cell division. Each chromosome was, therefore, independent of the others. When they separated into gametes, the set of chromosomes in each daughter cell could contain a mixture of the parental traits, but not necessarily the same mixture as that of other daughter cells. The newly discovered chromosomal independence during meiosis meant that the number of possible chromosomal combinations for each gamete could be calculated based on the number of chromosomes in the organism: specifically, there are 2n possible combinations of chromosomes in gametes, with "n" representing the number of chromosomes in the gamete. Furthermore, considering all the possible pairings of one gamete with another, the variation in zygotes is (2n)2, which results in some fairly large numbers. Indeed, Sutton provided examples of the potential variation among hypothetical organisms with gamete chromosome numbers ranging from 1 to 19 (Table 1).He also correctly assumed that there was more than one trait present on each chromosome, so the actual total variation was even higher than any of those included in the table.
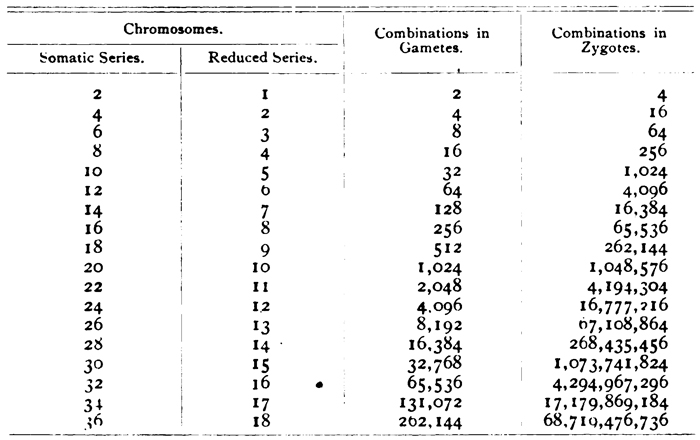
Table 1: Number of possible combinations in orgnaisms with 1 to 19 chromosome pairs in pre-synaptic cells.
Humans aren't even on this chart, because we have more than 36 chromosomes. But Sutton made this table before anyone knew that. If there were an entry in this table for 46 human somatic chromosomes, can you calculate the possible combinations of chromosomes Sutton would predict in human gametes?
© 1903 Marine Biological Laboratory Sutton, W. S., The chromosomes in heredity. Biological Bulletin 4, 231-251 (1903). All rights reserved. 
This observation of potential variation added strength to Sutton's assertion that he had discovered the physical basis for Mendel's principle of independent assortment. Still lacking, however, was definite proof. Scientists thus needed an experimental system in which the inheritance of genetic traits could be linked directly to the behavior of chromosomes. Such an opportunity presented itself soon thereafter, with a distinct mutation in the fruit fly Drosophila melanogaster.
Thomas Hunt Morgan Experimentally Demonstrates Chromosome Theory
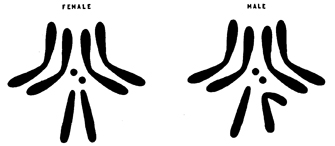
Figure 2: The characteristic pairing, size relations, and shapes of the chromosomes of Drosophila ampelophila.
In the male, an X and Y chromosome corresponds to the X pair of the female. The relative lengths of the chromosomes of this diagram are based upon averages of the measurements taken in other experiments; on the basis of X=100 the length of each long autosome is 159, of each small autosome 12, of the whole Y 112, of the long arm of the Y 71, and of the short arm 41.
© 1916 Genetics Society of America Bridges, C. B., Non-disjunction as proof of the chromosome theory of heredity, Genetics 1, 1-52 (1916). All rights reserved.
During the early years of the twentieth century, Thomas Hunt Morgan and his colleagues at Columbia University identified hundreds of Drosophila genes and made many pivotal discoveries about genetic transmission. Cytological examination showed that Drosophila possesses four pairs of chromosomes, including a pair of sex chromosomes. Female flies normally have two identical X chromosomes, whereas males have a single X chromosome and a smaller, gene-poor Y chromosome (Figure 2). Unlike humans, however, sex in fruit flies is determined by the number of X chromosomes, rather than by the presence of the Y chromosome.
One day, Morgan's associates discovered a male fly with unusual white eyes in their cultures. Breeding experiments quickly established that the white eye color was caused by a recessive mutation in the gene responsible for normal red eye color, and furthermore, that the gene was probably located on the X chromosome. The chromosome theory predicted that male flies would always display the eye color encoded on their single X chromosome, but that female flies would develop white eye color only when they inherited two mutant versions of the eye color gene. Thousands of matings confirmed this prediction, but on rare occasions, the group discovered "exceptional" white-eyed females among the progeny of a heterozygous female fly and a normal male, in apparent contradiction of the chromosome theory.
Lilian Vaughn Morgan, Thomas Morgan's wife, had the insight to recognize that these "exceptional" females might have an unusual chromosome composition. Indeed, these females had two X chromosomes as well as a Y chromosome; the XXY females resulted from defects in meiosis that caused a high frequency of nondisjunction, which is the failure of chromatids to separate during the second meiotic division. When eggs containing two nondisjoined X chromosomes, each of which carried the mutant white gene, were fertilized by a Y-bearing sperm, the product was an XXY female with white eyes. Rather than disproving the chromosome theory, these "exceptional" females actually provided strong experimental support that genes were in fact located on chromosomes.
Summary
Thus, the chromosome theory of inheritance was not the work of a single scientist, but rather the collaborative result of multiple researchers working over multiple decades. The seeds of this theory were first planted in the 1860s, when Gregor Mendel and Charles Darwin each proposed possible systems of heredity. It wasn't until several decades later, following Walther Flemming's discovery of chromosomes and description of their behavior during mitosis, that a probable mechanism for the transmission of traits was identified. The idea of a connection between chromosomes and heredity was subsequently strengthened by research conducted by Theodor Boveri and Walter Sutton, but direct evidence in support of chromosome theory didn't come until Thomas Hunt Morgan's experiments with fruit flies at dawn of the twentieth century. Thus, after nearly 50 years of speculation, scientists were finally able to confirm what they had long suspected: that chromosomes were indeed the physical carriers of hereditary information.
References and Recommended Reading
Benson, K. R. T. H. Morgan's resistance to the chromosome theory. Nature Reviews Genetics 2, 469–474 (2001) doi:10.1038/35076532 (link to article)
Bridges, C. B. Non-disjunction as proof of the chromosome theory of heredity. Genetics 1, 1–52 (1916) (link to article)
Brown, S. J. Entomological contributions to genetics: Studies on insect germ cells linked genes to chromosomes and chromosomes to Mendelian inheritance. Archives of Insect Biochemistry and Physiology 53, 115–118 (2003)
Cannon, W. A. Cytological basis for the Mendelian laws. Bulletin of the Torrey Botanical Club 29, 657–661 (1902)
---. Studies in plant hybrids: The spermatogenesis of hybrid cotton. Bulletin of the Torrey Botanical Club 30, 133–172 (1903)
Mendel, G. Gregor Mendel's letters to Carl Nägeli. Genetics 35, 1–29 (1950)
Sturtevant, A. H. A History of Genetics. (New York, Harper & Row, 1965)
Sutton, W. S. On the morphology of the chromosome group in Brachystola magna. Biological Bulletin 4, 24–39 (1902)
---.The chromosomes in heredity. Biological Bulletin 4, 231–251 (1903) (link to article)
Wilson, E. B. Mendel's principles of heredity and the maturation of the germ cells. Science 16, 991–993 (1902)
Winkelmann, A. Wilhelm von Waldeyer-Hartz (1836-1921): An anatomist who left his mark. Clinical Anatomy 20, 231–234 (2007)










