


















« Prev Next »

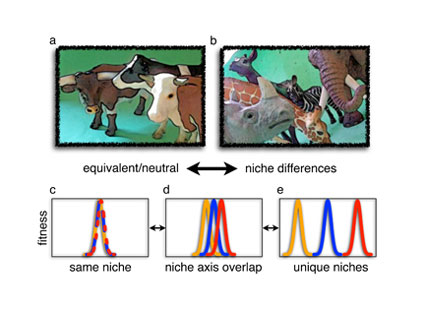
Figure 1: Neutral and niche
(A) Very similar herbivores might be considered to be equivalent and essentially neutral. (B) Herbivores that eat different plants, or even the same plants but at different times or at different heights, would have different or overlapping niches. In contrast, neutral theory suggests that even species that look very different from each other might be considered to be ecologically equivalent. (C-E) Species have greatest fitness at some point along an environmental gradient or niche axis. Neutral species have completely overlapping niches; they share the same niche and their fitness changes identically along an environmental gradient or niche axis. Greater niche differences correspond to less niche overlap between species; species differ in their fitness at different points along an environmental gradient or niche axis.
Building on foundational ideas described in island biogeography and the neutral theory of molecular evolution, the neutral theory of species diversity makes the provocative assumption that all individuals are ecologically identical, and that niche differences are not needed to explain biodiversity patterns. Individuals of certain species may all share characteristics that make them look or function different from other species, but those differences do not influence diversity. An individual in a community interacts with and experiences its neighbors as though they were exactly the same, regardless of species. This assumption of equivalence is the essential feature of neutrality, which differs from typical niche-based assumptions that an individual’s fitness depends on who its neighbors are: are they stronger or weaker competitors or do they belong to different species with different niche requirements (Figure 1)? Neutral theory predicts that species have perfectly overlapping niches — at the other extreme would be species with unique, non-overlapping niches. Real communities, of course, likely represent neither of these extremes but are somewhere in the middle (Figure 1). What neutral theory forces us to ask is how ecologically different are species, and how important are those differences for determining biodiversity?
How Neutral Theory Works
Under the neutral theory, highly diverse communities of equivalent species arise because chance extinctions are balanced by speciation. Specifically, the assumption of fitness equivalence combined with stochastic or random processes that include death, immigration from a regional pool of species, and speciation can lead to species-rich communities (Figure 2). As an example, imagine a field, which will be the local community, made up of many species of plants. Plants in the surroundings will represent the regional species pool. The field is thus part of a larger collection of plant communities called a meta-community. Individuals in the local community die at random and create openings for seeds to grow. Individuals from the meta-community, and from the local community, randomly disperse their seeds into the field (Figure 2). If there are more individuals of some species than others they will contribute more seeds than will rare species, but each individual seed has an equal chance of establishing. Individuals are in that sense competing for open sites. If dispersal from the meta-community is strong, the local community will look like a small version of the region; if dispersal from outside is weak, random deaths and extinctions combined with random mutations and speciation will cause the local community to drift and differ more and more from other communities over time, but in an unpredictable way. This random change in species abundance over time is termed ecological drift. There can be a limit to the number of individuals in the local community — this is known as a zero sum assumption: if the community is full, a new individual can only establish if another one dies and makes space (Figure 2).

Figure 2: Modeling a neutral process
A local community (a) represents a subset of all of the species found in the region or meta-community. Some individuals die at random in a local community (16 identical individuals from 6 species, in this example), which creates open sites (b). Open sites are then filled by random dispersal from both the meta-community and from the local community (c), including the addition of a new species from the meta-community. Random mutation (not shown) can also lead to the formation of new species. The sequence repeats leading to ecological drift where species abundances change randomly over time.
What Neutral Theory Is; What It Isn’t
Random death, dispersal and speciation are all important features of the neutral theory of biodiversity, but its one key, essential feature is the assumption of identical individuals (species may have differences, but those differences do not matter because all individuals have the same fitness and experience each other identically). The other features are assumptions about the processes that determine community assembly, or how species are added to and lost from communities, and how communities change over time — their dynamics. Another aspect of neutral theory is that it only applies to groups of similarly functioning species: trees in a forest or corals in a reef, and not to species of different size or trophic position, like microbes and elephants, or plants and herbivores. Also, there is no single neutral theory model, and different neutral models make different assumptions about these other processes. For example, one could imagine a neutral community — made up of identical individuals of multiple species — without immigration. Neutral models don’t need to follow the zero sum assumption: a local community could remain partly empty, or alternatively the numbers of individuals might be allowed increase in a model continuously. Stochastic processes are important in neutral models for communities to change over time, but some parameters might be random while others might be non-random: annual plants always die at the end of the year. Niche models, where individuals of different species do differ from each other and those differences matter for their fitness, can also be stochastic, and some purely deterministic or truly non-random processes can also be unpredictable (see chaos theory) and only apparently random. Stochasticity or randomness is often just a simplification we make when we can’t predict exactly the outcome of an event: assuming something is random allows us to conveniently describe a process in terms of probability or chance.
Although dispersal, stochasticity and speciation are not unique to the neutral theory, neutral models are more interesting and useful when we add in things like dispersal and stochasticity. For this reason, neutral theory is often described as a “dispersal-assembly” theory or a “stochastic” theory, even though neither dispersal nor stochasticity is a feature uniquely or necessarily neutral. Dispersal and reproductive traits are in fact another way that species can differ in their fitness, and how organisms move through space can be an important component of a species’ niche. Consider the enormous variety shown by flowering plants in their fruits (coconuts to dandelion seeds) and the diverse ways plants can disperse by wind, water or animals, as well as their different mating strategies, germination and pollination mechanisms. Dispersal may often be highly unpredictable, but it is not necessarily neutral.
Dispersal and speciation processes in neutral models can lead to highly diverse communities. But the coexistence of species in neutral communities is unstable — there are no mechanisms that cause one species to remain dominant or prevent rare species from going extinct. Because neutral processes are driven by random events, and because all individuals are competitively identical, their abundances either increase or decrease purely by chance. In a closed system, stable coexistence, or long-term persisting species, can only occur when there are niche differences that cause individuals to compete most strongly with individuals of their own species compared to those of other species. Niche-based stabilizing mechanisms limit the growth of species when they become very abundant, while at the same time stabilizing mechanisms allow rare species to increase because they enjoy higher fitness when surrounded by neighbors of different species with whom they compete less strongly.
Although dispersal, stochasticity and speciation are not unique to the neutral theory, neutral models are more interesting and useful when we add in things like dispersal and stochasticity. For this reason, neutral theory is often described as a “dispersal-assembly” theory or a “stochastic” theory, even though neither dispersal nor stochasticity is a feature uniquely or necessarily neutral. Dispersal and reproductive traits are in fact another way that species can differ in their fitness, and how organisms move through space can be an important component of a species’ niche. Consider the enormous variety shown by flowering plants in their fruits (coconuts to dandelion seeds) and the diverse ways plants can disperse by wind, water or animals, as well as their different mating strategies, germination and pollination mechanisms. Dispersal may often be highly unpredictable, but it is not necessarily neutral.
Dispersal and speciation processes in neutral models can lead to highly diverse communities. But the coexistence of species in neutral communities is unstable — there are no mechanisms that cause one species to remain dominant or prevent rare species from going extinct. Because neutral processes are driven by random events, and because all individuals are competitively identical, their abundances either increase or decrease purely by chance. In a closed system, stable coexistence, or long-term persisting species, can only occur when there are niche differences that cause individuals to compete most strongly with individuals of their own species compared to those of other species. Niche-based stabilizing mechanisms limit the growth of species when they become very abundant, while at the same time stabilizing mechanisms allow rare species to increase because they enjoy higher fitness when surrounded by neighbors of different species with whom they compete less strongly.
Pattern versus Process
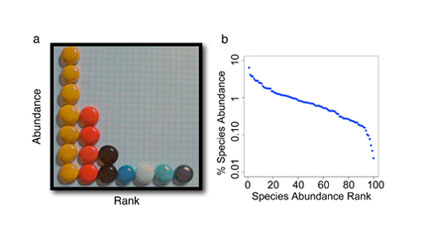
Figure 3: Species abundance patterns
(A) The individuals from the example in Figure 2c are ranked 1 through 7, from most to least abundant species. (B) An example of a simulated community of 100 species showing a characteristic pattern of species rank abundance.
The Utility of Neutral Theory
Neutral theory is still a powerful and useful concept for several reasons. Stephen Hubbell’s 2001 monograph, The Unified Neutral Theory of Biodiversity and Biogeography, has sparked vigorous debate among ecologists, and has led to more rigorous, and much needed, tests of niche mechanisms and explanations of biodiversity. Ironically, neutral theory has reinvigorated niche theory. As with all models, the neutral model is a simplification, albeit a drastic one, of processes we think important in the natural world (e.g., competition, dispersal). Thus, the strength of the neutral model is that it provides us with a logical place to start: an elegant and simple null model, with clear and testable assumptions and predictions. We can then ask if other mechanisms, and their added complexity, are necessary to explain what we observe.
Summary
The neutral theory of species diversity starts with the key assumption that all individuals in a community of trophically similar species are ecologically identical. Neutral models that additionally include random death, speciation, extinction, and dispersal from the meta-community can lead to highly diverse communities that have similar species abundance patterns to what we observe in real communities. The abundances of species in neutral models fluctuate randomly over time leading to ecological drift, where diversity is due to unstable coexistence and the balance between extinction and speciation, in contrast to niche models that assume the importance of species’ niche differences and stabilizing mechanisms. Neutral theory provides a null model, or a starting point, from which we can test niche-based hypotheses for how species’ evolutionary adaptations and niches maintain biodiversity.
References and Recommended Reading
Adler, P. B., HilleRisLambers, J. et al. A niche for neutrality. Ecology Letters 10, 95-104 (2007).
Alonso, D., Etienne, R. S. et al. The merits of neutral theory. Trends in Ecology and Evolution 21, 451-457 (2006).
Bell, G. Neutral macroecology. Science 293, 2413-2418 (2001).
Chase, J. M. & Leibold, M. A. Ecological Niches: Linking Classical and Contemporary Approaches. Chicago, IL: University of Chicago Press, 2003.
Hubbell, S. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press, 2001.
McGill, B. J. A test of the unified neutral theory of biodiversity. Nature 422, 881-885 (2003).
McGill, B. J., Maurer, B. A. et al. Empirical evaluation of neutral theory. Ecology 87, 1411-1423 (2006).
Silvertown, J. Plant coexistence and the niche. Trends in Ecology and Evolution 19, 605-611 (2004).
Tilman, D. Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences, USA. 101, 10854-10861 (2004).










