


















« Prev Next »

The unparalleled diversity of tropical ecosystems has drawn the attention of naturalists since the earliest voyages of discovery in the eighteenth and nineteenth centuries. The fundamental question then, as now, is: Why so many species? One influential perspective seeks to explain species richness at the community level in terms of mechanisms for ecological coexistence (Gause 1934, Hutchinson 1957). The central idea is that each species occupies a unique ecological niche, a distinct functional role within the ecosystem (Darwin 1859). Under this view species richness arises from the actions of natural selection to reduce competition or increase adaptive specialization.
Another view stresses the importance of historical or phylogenetic aspects of species diversification (Whittaker 1972). For example, the "species-pool" hypothesis describes local diversity by reference to the size (species richness) of the regional or global pool of species from which the local assemblage may be recruited (Taylor et al. 1990). From a macroevolutionary perspective, net rates of diversification within a geographic region arise from differential rates of speciation, extinction, and immigration (Rosensweig 1995, Stanley 1998, Jablonski 2000). Although the mechanisms of these processes remain incompletely understood for most biotas, we have known for at least a century and a half that evolutionary diversification takes place both in space and time. Biodiversity, biogeography, and paleontology are therefore all intimately related subjects.
Exceptional Amazonia
Amazonia exhibits all the principle features associated with aquatic species richness globally (Albert & Reis 2011). It is the greatest interconnected fluvial system on the planet, extending over more than seven million km2. The Amazon River alone carries about a sixth of the all world's flowing freshwater. Its tropical location, straddling the equator, means it receives high levels of solar radiation and precipitation, and the region is largely covered by humid lowland rainforests. From a purely ecological perspective, therefore, Amazonia as it exists today appears to be an excellent environment for diversification in fishes.
Yet the species-rich Amazonian ichthyofauna is ancient and did not arise under the geological conditions of the modern Amazon Basin (Lundberg et al. 2010). Direct fossil evidence shows that many phenotypes and lineages date to the early Neogene or Paleogene (16 to 65 million years ago; Ma), before the Amazon assumed its modern courses. For example, the Amazon River only adopted its current east-flowing course after about 11 Ma; prior to this it flowed west and north into the Caribbean (Figueiredo et al. 2009).
Furthermore, data from species-level phylogenetic and biogeographic studies tell us that Amazonian ichthyofaunas accumulated incrementally over a period of tens of millions of years, principally by means of allopatric speciation, and in an arena extending over most of the area of tropical South America (Albert & Reis 2011). In other words, unlike some of the well-known insular faunas (Galapagos finches, Hawaiian drosophilid flies, African rift lake cichlids), the species-rich Amazonian ichthyofauna is not the result of recent adaptive radiations.
Vicariance and Geodispersal
The geological history of landscapes is tightly linked with the evolution of their resident biotas. For freshwater organisms, landscapes are divided naturally into discrete drainage basins by watersheds, episodically isolated and reunited by erosional hydrodynamics. In regions like the Amazon Basin with an exceptionally low (flat) topographic relief, the many waterways have had a highly reticulated history over geological time.
Stream capture is an important geographic factor affecting the evolution and distribution of freshwater organisms. Stream capture occurs when an upstream portion of one river drainage is diverted to the downstream portion of an adjacent basin. This can happen because of geophysical uplift (or subsidence), natural damming as a result of a landslide, or by headward or lateral erosion of the watershed between adjacent basins.
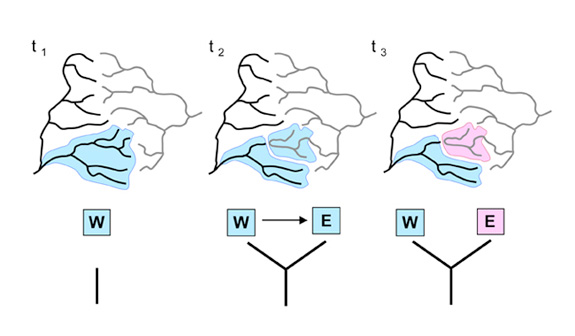
Figure 1: Effects of stream capture on species-richness of adjacent drainage basins based on dispersal, vicariance, and extinction
Stream capture event at t2 simultaneously separates and permits dispersal of populations between adjacent basins. In this hypothetical example the newly diverged species at t3 may subsequently disperse throughout the Eastern basin (E). Stream capture also changes the total area of each basin, and by means of the species-area relationship, the rates of speciation and extinction. The area of the Western basin (W) decreases in area, and may therefore maintain fewer species over evolutionary time.
© 2010 Nature Education All rights reserved. 
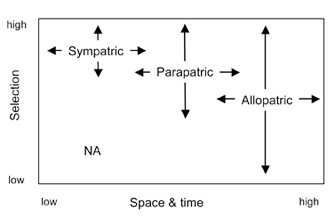
Figure 2: Modes of speciation differ based on the relative roles of space (geography) and ecology (selection) in the formation of barriers to gene flow
NA indicates that speciation is unlikely in sympatry when selection is very weak.
© 2010 Nature Education All rights reserved.
Geodispersal of a geographic area is different from the physical movements of organisms (dispersal), although the erosion of geographic barriers can and does facilitate organismal dispersal. Geodispersal, like vicariance, is an Earth history process while dispersal is a biotic process. As Earth history events, both vicariance and geodispersal may be spatially and temporally extensive, affecting most or all of the evolving lineages in a regional biota.
In tropical South America groups of closely related fish species usually exhibit non-overlapping geographic ranges, a pattern suggesting divergence in allopatry (Albert & Reis 2010). Allopatric speciation occurs when two or more populations of a species become genetically isolated following the formation of barriers to dispersal and gene flow. However, species-level phylogenies and distributions cannot be used to unambiguously prove a particular geographic mode of speciation (e.g., sympatric v. allopatric) because geographical ranges may change following speciation.
Permeability of Geographical Barriers
The emergence of large tectonic structures, like ocean basins and mountain chains, usually forms impermeable barriers to dispersal and gene flow. Many groups of plants and animals, including freshwater fishes, were permanently isolated on either side of the newly-formed Atlantic Ocean during the final separation of Africa from South America in the Upper Cretaceous (around 110 Ma). Similarly, the early Neogene (around 12 Ma) rise of the Eastern Cordillera of the northern Andes permanently separated lowland northern South America into taxonomically distinct cis- and trans-Andean biotas (Albert et al. 2006). Impermeable barriers such as these are often spatially expansive, affecting broad geographic areas and multiple phylogenetically independent clades. Impermeable barriers are therefore excellent systems to study the role of vicariance in the formation of regional biotas.
Yet tectonic activity is highly punctuated in time and space. Most mountain building in the Andes occurred in relatively discrete pulses lasting just 5–10 million years each, interrupted by much longer periods of tectonic quiescence. By contrast, most of lowland tropical South America has changed gradually through the perennial action of erosion. Stream capture results in subtle and complex patterns in the geographic separation and merging of river basins and freshwater biotas. As noted above, a single stream capture event has both vicariant and geodispersal effects, simultaneously isolating a headwater tributary from one basin and connecting it to that of an adjacent basin.
In tropical South America, with its flat topography and low-altitude watersheds, many basins boundaries have been semipermeable over geological time scales (Lovejoy et al. 2010a). Semipermeable watersheds are environmental filters in the sense that they allow differential dispersal among adjacent basins. Fish species with high vagility between basins are those able to inhabit small streams and seasonal wetlands of the intervening areas and to tolerate suboptimal environmental conditions (e.g., dysoxia, variable temperatures). Species that are ecologically restricted to large lowland rivers and floodplains have far less vagility across watersheds.
Stream capture also differentially affects taxa depending on their demographic susceptibilities to extinction. Populations isolated on either side of a newly-formed watershed are likely to have reduced population sizes, which may accelerate genetic divergence and speciation or lead to local extinction. Taxa with high riverine vagility may use the new connections to expand their ranges and avoid extinction. Such newly arrived exotics may also adversely affect members of the resident fauna. Although the responses of individual taxa are varied, the response of a fauna as a whole to stream capture is likely to be increased rates of both speciation and extinction; in other words, increased rates of net diversification.
Phylogenetic and biogeographic patterns in the electric fish Gymnotus (Gymnotidae) illustrate how stream capture across low-elevation watersheds on the relatively flat South American platform contribute to elevated species richness of regional assemblages (Figure 3). The Neogene rise of the northern Andes resulted in relatively impermeable barriers to fish dispersal, isolating cis- and trans-Andean lineages in both the G. carapo and G. tigre groups. By contrast, most watersheds in lowland Amazonia are relatively more permeable, several of which are indicated by dashed gray bars in Figure 3. Due to post-speciation dispersal, the Gymnotus species of the Amazon basin do not form a monophyletic group but rather represent an assemblage of species with origins in several basins (Albert et al. 2004).
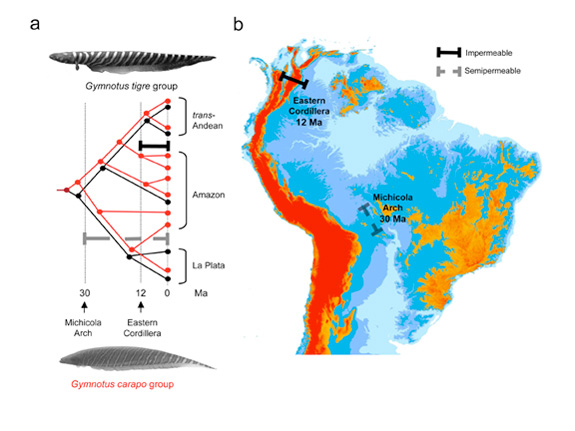
Figure 3: Effects of vicariance and geodispersal on the formation of regional assemblages
(A) Schematic phylogenies of two clades of the electric fish Gymnotus (Gymnotidae). Some species of the G. carapo group (in red) are omitted for clarity. Thick horizontal bars represent geographic barriers. Note polyphyletic origin of the Amazonian assemblage. (B) Paleogeographic vicariance and coupled geodispersal events used to constrain divergence times. Dates represent minimum lineage divergence time estimates in millions of years ago (Ma). Semipermeable watersheds are "leaky" and do not provide reliable minimum divergence times (i.e., dates for semipermeable barriers represent date ranges that extend to 0 Ma). Data from Lovejoy et al. (2010a, 2010b).
© 2010 Nature Education All rights reserved.
Phylogenetic Niche Conservatism
The empirically observed predominance of allopatric (versus sympatric) speciation highlights the important role of geography in the formation of species richness. However, it does not mean that diversification occurs in an ecological vacuum. Species differ substantially in how they respond to the formation and erosion of geographic barriers and exhibit different patterns of dispersal and demographic cohesion. Even under a purely allopatric model of speciation, the autecological requirements of a species help define the nature and permeability of barriers to gene flow and dispersal. Furthermore, the idiosyncratic ecological requirements of individual species may affect the ways in which populations disperse as geographic barriers form and become eroded. The very existence and strength (i.e., permeability) of geographic barriers to dispersal and gene flow emerge directly from the ecological and habitat requirements peculiar to a species.
Vicariant speciation is in this view a consequence of phylogenetic niche conservatism: a failure to adapt to the conditions of an intervening habitat. Niche conservatism is the tendency of species to retain ancestral ecological characteristics. The conditions for ecological speciation are more restrictive than for allopatric speciation (Figure 2). Because adaptive (parapatric or sympatric) speciation is so much less common than non-adaptive allopatric speciation, species tend to retain their ancestral ecological characteristics.
Formation of Regional Assemblages
Niche conservatism is also important in constraining the assembly of species into local and regional biotas (Cornell & Lawton 1992). Species become part of a local assemblage as immigrants from an adjacent area or as colonists from different habitats in the same area (Ricklefs 2008, Winemiller et al. 2008). Because of niche conservatism, immigration from ecologically similar areas is more likely than adaptive evolution (Wiens 2004). Immigration is easier if a species is already adapted to local habitat conditions. In this case, a species need only overcome the ecological or physical barriers to dispersal. If a species is already present in a region but in a different habitat, dispersal requires the evolution of new ecological or physiological traits. Hence local communities are more often assembled by recruitment of species from a wide landscape. This seems to the rule among Neotropical fishes, in which the great majority of species originated in allopatry without ecological divergence (Albert et al. 2011). For example, cases in which one sister species occupies deep river channels while the other occupies seasonally-anoxic floodplain systems are not known among electric knifefishes (Crampton 2011). In contrast, there are many cases of species in floodplains and rivers that have sister species occupying the same habitat in an area of geographical allopatry (e.g., the deep river specialist Compsaraia samueli from the Amazon and C. compsus from the Orinoco). To summarize, while adaptive transitions between very distinct ecosystems are rare, immigration by pre-adapted species is likely the dominant way in which local communities fill with species or replace species that become extinct.
Conclusion
Phylogenetic and biogeographic patterns in Neotropical freshwater fishes suggest that most speciation has occurred along geographic, not ecological lines. In most groups of Neotropical fishes, diversification occurred incrementally over large spatial and temporal scales, with speciation occurring over much of the continental platform and requiring tens of millions of years. Vicariance and geodispersal are complimentary biogeographic processes that have profoundly influenced the formation of new species and the taxonomic composition of regional biotas. Together these processes interact in a complex duet of Earth history events and biological diversification. These results from Neotropical freshwater fishes support an emerging paradigm shift within the field of ecology: Geography and phylogenetic history play an immensely important role in the formation of local species assemblages.
References and Recommended Reading
Albert, J. S., & Reis, R. E. Historical Biogeography of Neotropical Freshwater Fishes. Berkeley, CA: University of California Press, 2011 (in press).
Albert, J. S., Crampton, W. G. R., et al. Phylogenetic systematics and historical biogeography of the Neotropical electric fish Gymnotus (Teleostei: Gymnotiformes). Systematic and Biodiversity 2, 375–417 (2004).
Albert, J. S., Lovejoy, N. R., et al. Miocene tectonism and the separation of cis- and trans-Andean river basins: Evidence from Neotropical fishes. Journal of South American Earth Sciences. 21, 14–-27 (2006).
Cornell, H. V., & Lawton, J. H. Species interactions, local and regional processes, and limits to the richness of ecological communities. Journal of Animal Ecology 61, 1–12 (1992).
Crampton, W. G. R. An ecological perspective on diversity and distributions. In Historical biogeography of Neotropical Freshwater Fishes, eds. Albert, J. S & Reis, R. E. (Berkeley, CA: University of California Press, 2011 [in press]).
Darwin, C. R. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle For Life. London, UK: John Murray, 1859.
Figueiredo, J., Hoorn, C., et al. Late Miocene onset of the Amazon River and the Amazon deep-sea fan: Evidence from the Foz do Amazonas Basin. Geology 37, 619–622 (2009).
Gause, G. F. The Struggle For Existence. Baltimore, MD: Williams and Wilkins, 1934.
Hutchinson, G. E. Concluding remarks. Cold Spring Harbor Symposium Quantitative Biology 22, 415–427 (1957).
Jablonski, D. Micro- and macroevolution: scale and hierarchy in evolutionary biology and paleobiology. Paleobiology 26, 15–52 (2000).
Lovejoy, N. R., Willis, S. C., et al. Molecular signatures of Neogene biogeographic events in the Amazon fish fauna. In Amazonia, Landscape and Species Evolution: A Look into the Past, eds. Hoorn, C. M. & Wesselingh, F. P. (London, UK: Blackwell Publishing, 2010a): 405–417.
Lovejoy, N. R., Lester, K., et al. Phylogeny, biogeography, and electric signal evolution of Neotropical electric fishes of the genus Gymnotus (Osteichthyes: Gymnotidae). Molecular Phylogenetics and Evolution 54, 278–290 (2010b).
Lundberg, J. G., Sabaj Pérez, M. H., et al. The Amazonian Neogene fish fauna. In Amazonia, Landscape and Species Evolution: A Look into the Past eds. Hoorn, C. M. & Wesselingh, F. P. (London, UK: Blackwell Publishing, 2010): 281–301.
Ricklefs, R. E. Disintegration of the ecological community. American Naturalist 172, 741–750 (2008).
Rosensweig, M. L. Species Diversity in Space and Time. Cambridge, MA: Cambridge University Press, 1995.
Stanley, S. M. Macroevolution. Pattern and Process. Baltimore, MD: The Johns Hopkins University Press,1998.
Taylor, D. R., Aarssen, L. W., et al. On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies. Oikos 58, 239–250 (1990).
Whittaker, R. H. Evolution and measurement of species diversity. Taxon 21, 213–251 (1972).
Winemiller, K. O., Lopez-Fernandez, H., et al. Fish assemblages of the Casiquiare River, a corridor and zoogeographical filter for dispersal between the Orinoco and Amazon basins. Journal of Biogeography 35, 1551–1563 (2008).
Wiens, J. J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 58, 193–197 (2004).
Wilkinson, M. J., Marshall, L. G. et al. River behavior on megafans and potential influences on diversification and distribution of aquatic organisms. Journal of South American Earth Sciences 21, 151–172 (2006).









