



















« Prev Next »
Introduction
Homo sapiens is the only living representative of the tribe Hominini, but of course this was not always the case; our ancestors overlapped in time and space with other species throughout the Plio-Pleistocene. There are numerous factors that may be involved in the evolution or extinction of a species, but primary amongst them are ecological conditions and a species' ability — or inability — to adapt to them when they change. It is critical to our understanding of human evolution to make sense of the patterns of speciation, extinction, and morphological and behavioural variability observed in the fossil and archaeological record in light of past ecologies.There are many useful approaches to the reconstruction of the ecological and environmental settings of human evolution and the theory and methods used to reconstruct them overlap to a certain extent. The differences between them may be characterized as such: the science of paleoecology addresses the relationships between both plant and animal species and their environments whilst paleoenvironmental research tends to be more focused on the physical conditions of the surrounding area including terrain and vegetation structure, as well as characteristics such as temperature and rainfall. Paleoclimatology, a related discipline, is concerned with these sorts of characteristics and reconstructing broad, long-term records of them at a global or regional level, but it is also important to investigate the local effect of these variables at sites where hominins are known to have lived.
Mammals: Past and Present
Mammals provide a useful illustration of the relationships that exist between species and their environments. Every mammalian species requires two important things from its natural habitat for day-to-day survival: dietary resources for meeting their energy requirements and space in which to eat, sleep and move around. The dietary, or trophic, and spatial niches that are available for each taxon to fill in any given place will be dictated by biotic factors such as the types of plant and animal life present and abiotic factors such as soil type, rainfall, temperature and seasonality. These factors will determine which organisms can survive in a given habitat.
Let's consider two different types of African environments, a forest and an open woodland-bushland, which are created by variations in these biotic and abiotic factors. We'll focus on the different mammals that are capable of living in each of them and take specific examples from a family of ungulates, Bovidae, and the carnivores that prey upon them (species information from Kingdon 1997) (Figure 1a-d). In tropical latitudes where moisture and high annual temperatures create conditions that support forests with multistory tree canopies and reliable cycles of fruiting and flowering vegetation, there will be an abundance of species that are adapted for living in the arboreal substrate; many of these will be frugivores and browsers, subsisting on fruit and leaf-based diets, respectively. Small bovids called duikers are well-adapted for these forest conditions; Peters' duiker, Cephalophus callipygus, consumes one of the most fruit-dominated diets of any extant bovid (Fig 1a). Its natural predators are cat species in the family Felidae such as the leopard (Panthera pardus) (Fig 1b) and African golden cat (Profelis aurata), both of which are adapted for moving quickly up and down trees, which provide them with a safe place to consume their prey after catching them on the ground.

Figure 1: Photos of bovids and carnivore predators.
a) Peters’ duiker (Cephalophus callipygus) is commonly preyed upon by cat species such as b) the leopard (Panthera pardus). Both taxa can be found in African forested habitats. Where habitats in Africa are less wooded and more arid, bovids such as c) Grant’s gazelle (Nanger granti) are preyed upon by d) jackals (Canis aureus) and other species that are better adapted for killing prey in more open terrain.
© 2012 Nature Education (a), (b) and (d) courtesy of Wikipedia, (c) courtesy of K. Kovarovic. All rights reserved. 
In contrast, where conditions are more arid and temperature and rainfall fluctuates seasonally, more open woodland-bushland, scrub growth and grass cover may exist. Thus, the mammal community would comprise a proportionally greater number of species that are adapted for grazing on grasses and for terrestrial locomotion. A bovid commonly found in more open country settings is Nanger granti, Grant's gazelle, which will eat fresh grasses when they are seasonably available and herbs and leaves during the remaining times of the year (Fig 1c). This species is preyed upon by felids that stalk in open areas such as the cheetah (Acinonyx jubatus), as well as the jackal (Canis aureus) (Fig 1d), which is a member of the dog family, Canidae. Although Grant's gazelle is a speedy runner, it cannot always avoid predation by these fast and stealthy carnivores.
Looking to the dietary and locomotor behavior of multiple taxa living in one locality, we may evaluate the way in which an entire mammalian community fills spatial and trophic niches, creating a picture of the overall mammal community structure. Trophic and spatial niches are subdivided into categories indicative of the dietary and locomotor adaptations of each species and the proportions of species in each category are calculated to facilitate comparison across localities. Take a look at Figure 2. It illustrates how two modern African locations characterized by habitats like those detailed above have unique mammal community structures. The charts show locomotor and dietary traits of the mammal species that live in two different habitats, a tropical forest and a mixed woodland bushland. Try to evaluate these data and make hypotheses as to which one is which.
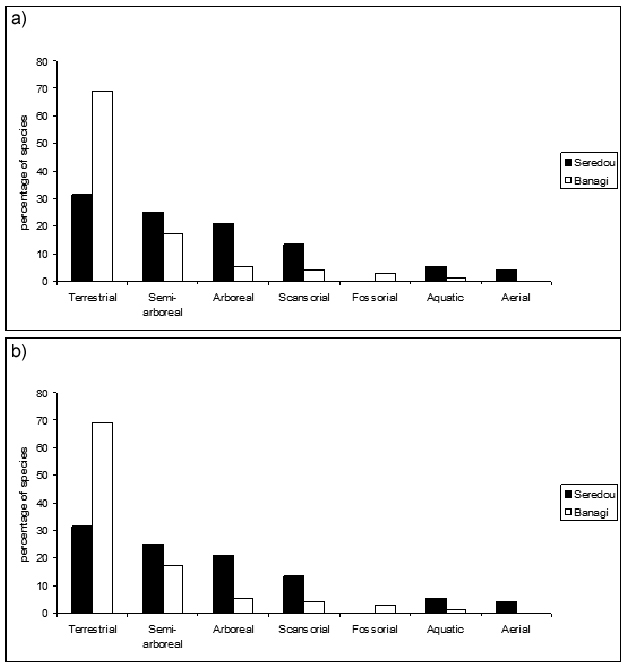
Figure 2
The percentage of a) locomotion and b) dietary adaptations of the mammals found in Seredou, Guinea (white bars) and Banagi, Tanzania (black bars). Which one is a tropical African forest and which one is an African woodland-bushland habitat?
© 2012 Nature Education All rights reserved.
Figure 2a details the percentage of locomotor adaptations (in addition to familiar categories such as terrestrial locomotion, specialist locomotion patterns such as scansoriality, a climbing adaption common in cats, and fossoriality, a digging and underground burrowing adaptation typical of some rodents, are considered) and Figure 2b displays the percentage of diets of the mammals present at these locales. Approximately 30% of the species in the Seredou forest are terrestrial, but if you combine locomotion categories pertaining to movement in the trees (arboreal, semi-arboreal and scansorial), there are twice as many of these species (59%) than terrestrial. Additionally, Seredou has a proportionally greater number of frugivores, who rely on fruiting trees. In contrast to Seredou, mammals resident in Banagi are predominantly terrestrial (70%). Banagi also has a greater number of grazing species and carnivores — presumably those that prey on the terrestrial grazing species which are so much more abundant there. Seredou, Guinea, is an African tropical forest, whereas Banagi, Tanzania is a mixed woodland-bushland.
To infer paleoecology in the fossil record we work backwards by evaluating the trophic and locomotor adaptations of the mammals at a given locality in order to reconstruct what ecological conditions supported a community in the past. Because bones and teeth, not ecologies, preserve as fossils, we rely on our understanding of how their overall shapes and other measureable characteristics (such as joint surfaces) relate to the exploitation of particular niches. Skeletal and dental morphologies that are shown to be adaptations to specific niches and habitat types are known as ecomorphologies. Dental ecomorphologies tell us about the diet that a species was adapted for and postcranial ecomorphologies (from the limbs in particular) tell us what mode of locomotion the species used. Ecomorphologies are assessed by taking direct measurements and statistically analyzing them to look for significant correlations with aspects of the organism's ecology such as eating particular diets or locomoting in specific spaces. Often paleoecologists focus on an evaluation of the ecomorphologies displayed by species of only one taxonomic group in an assemblage rather than a complete community diversity approach, like the one detailed above. By carefully selecting a family that is diverse , abundant and well-preserved at a fossil site, we avoid difficulties that may affect community-based analyses, in particular the taphonomic conditions that often delete certain types of species — and their ecological information — from the record. This commonly afflicts small-bodied taxa whose remains are not as robust as larger species and are therefore often destroyed naturally during the fossilization process. Certain carnivores are also capable of destroying a significant portion of the identifiable remains of their prey, so they too may be under-represented in some circumstances.
Laetoli, Tanzania: a Case Study
Community diversity and ecomorphological approaches are complementary techniques for reconstructing ecological conditions at one place in one point in time. Assemblages of fossil mammals from the same site can also be viewed temporally if the depositional contexts have preserved remains in discrete and datable strata, so that trends can be revealed. These techniques will be demonstrated using the site of Laetoli, Tanzania as a case study. This Plio-Pleistocene site is located south of Olduvai Gorge (Figure 3) and preserves the tracks and remains of a diverse faunal community that includes hominins such as Australopithecus afarensis and Paranthropus aethiopicus in the upper units of the Laetolil Beds (3.85–3.63 mya) and Ndolanya Beds (2.66 mya), respectively (references in Harrison, 2011 and Leakey & Harris 1987).
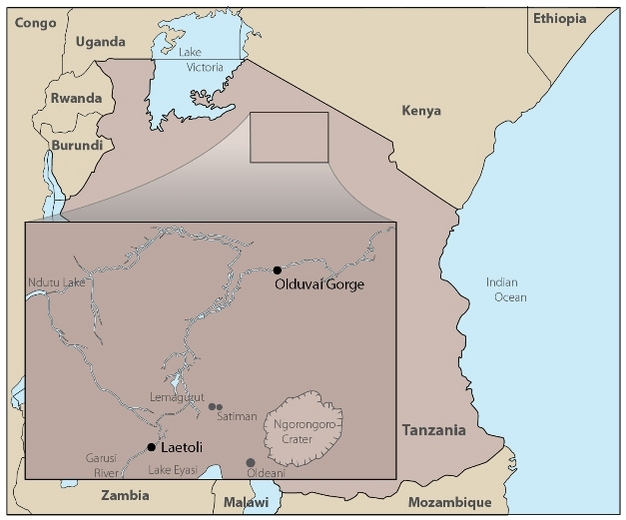
Figure 3: Location of Laetoli, Tanzania.
It is just south of Olduvai Gorge, where other hominin remains and artefacts are found.
© 2012 Nature Education All rights reserved.
Community Diversity
Previous work on the Upper Ndolanya Beds identified 44 mammalian species in the community (Ndessokia 1989). These species have been placed in a category that describes their body weight, locomotion pattern and diet. Figure 4 a-c illustrates the percentage of the Ndolanya species assigned to each category, showing us how the community fills the spatial and trophic niches; for instance, a high proportion of mammals exploit both the terrestrial (Fig. 4b) and grazing (Fig 4c) niches (Kovarovic et al 2002). In isolation this information can not tell us as much as it can when compared to other habitats. Figure 4d displays the proportion of mammalian locomotion adaptations found in many modern habitats around the world compared to the Upper Ndolanya Beds on the right-hand side of the graph. The Ndolanya Beds are unique because they have a much higher number of terrestrial species compared to the modern habitats, which can be explained by the taphonomic loss of some (probably small-bodied) species that move in other ways. However, when the overall pattern of the community structure is observed, it can be seen that the Ndolanya Beds are the most similar to African tropical arid bushland habitats (situated next to the Ndolanya Beds on the graph). These, too, have a high proportion of terrestrial species with few relying on arboreal, semi-arboreal and scansorial modes of movement. This is used to infer that the Ndolanya Beds represent an arid bushland habitat at 2.66 mya.
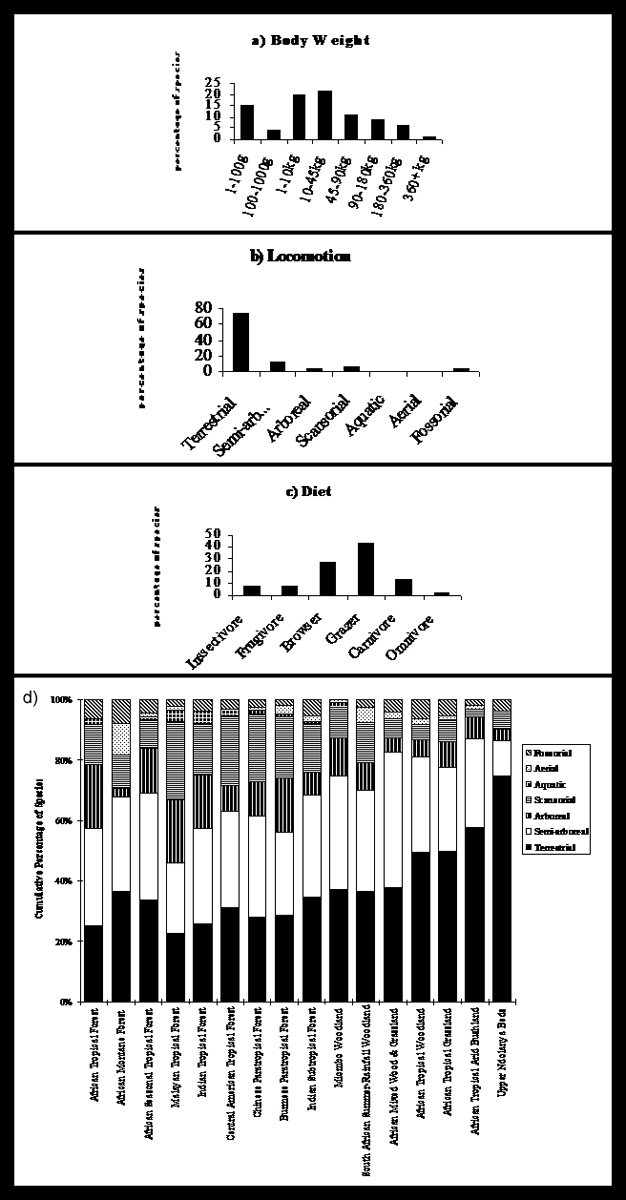
Figure 4
Classification of the a) body weight, b) locomotion and c) diet of the mammals from the Upper Ndolanya Beds at Laetoli, Tanzania. d) A comparison of the percentage of mammals adapted for different forms of locomotion in many modern habitats around the world. The Upper Ndolanya Beds are on the right-hand side of the chart (based on Kovarovic et al 2002).
© 2012 Nature Education All rights reserved.
The Upper Laetolil Beds, however, are approximately one million years older than the Ndolanya Beds, and a great deal can change over that length of time. Was Laetoli always like the arid bushland habitat observed at 2.66 mya? A community diversity analysis of these beds, focusing on mammals larger than 500 grams (which are more likely to survive in the fossil record), has noted that the Upper Laetolil Beds have a higher proportion of carnivores than the Upper Ndolanya Beds but that overall both beds are quite similar to modern Serengeti communities in the way the trophic and spatial niches are exploited (Su 2011). Earlier work, however, has shown that the Upper Laetolil fauna most closely resembles communities from wooded Serengeti habitats and not those from the open plains (Andrews 1989).
Ecomorphology
Investigations of the Laetoli bovid ecomorphologies lends weight to the conclusions drawn from ecological diversity studies. Without specific regard for the identity of the species present, bovid limb bone morphologies indicate that a greater percentage of Upper Laetolil Beds bovids possessed adaptations for wooded habitats than those from the Upper Ndolanya Beds (Bishop et al 2011; Kovarovic & Andrews 2007). It is interesting to note this shift in habitat types between the two beds, but the Upper Laetolil Beds themselves were deposited over a period of approximately 300,000 years. Are there ecological trends that can be reconstructed during this time?
Many paleontological sites have quite a finely reconstructed chronological sequence and fossils can be easily associated with datable units that sample short periods of time. Their sediments and fossils have often been quickly deposited through the action of water, such as the meandering of rivers or the sequential expansion and drying up of ancient lakes. However, Laetoli was not associated with a large permanent water source and fossils were deposited on the surface of the landscape and buried naturally. Chronological units at Laetoli are instead defined by dated volcanic tuffs and by lumping fossil finds together into stratigraphic units bounded by these tuffs, we can analyse the material as a progression. Although these units of time are fairly long compared to other geological sequences, it becomes clear that the ecology was not unchanging during the Upper Laetolil Beds. The percentage of ecomorphologies in four units, from the oldest to the youngest (left to right), are illustrated in Figure 5 (Kovarovic & Andrews 2011). Throughout the sequence at any given time heavy wood-bushland is the best represented group of ecomorphologies, whilst forest and grassland are not well-represented. The shift that occurs relates to the woodland habitats. The two youngest units have similar proportions of ecomorphologies indicating similar availability of habitat types, however things change halfway through the sequence after Tuff 5 when wooded-bushed grassland increases and overtakes the amount of light wood-bushland adaptations. It may seem like a subtle difference, but it means that grass cover was increasing at the expense of tree cover, thereby changing the availability of the trophic and spatial niches to which these bovids were best adapted. Of course, adaptations for an array of habitats are displayed by both Laetolil and Ndolanya communities, emphasizing that environments are not homogenous but generally characterized by a mosaic of different habitat types.
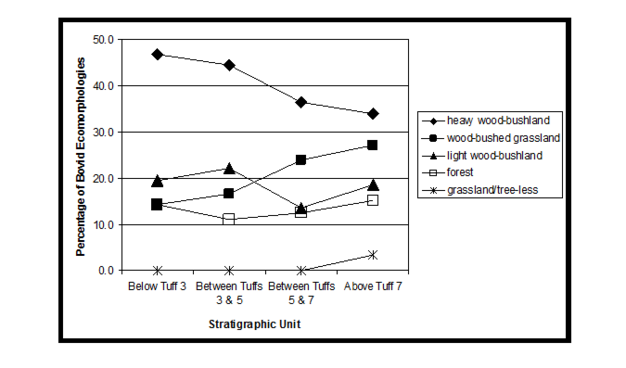
Figure 5
The percentage of bovid ecomorphologies in four units of the Upper Laetolil Beds, displayed from the oldest to the youngest (left to right) (based on Kovarovic & Andrews 2011). Over time, the proportion of wooded-bushed grassland adapted individuals increases.
© 2012 Nature Education All rights reserved.
Conclusion
The Laetoli region has always been characterized by habitat variability, but it had more woodland cover available to its inhabitants in the past than it does today (references in Harrison 2011). The transition to a more open mosaic of habitats began deep in the past and is observable in the fossil record both between the Upper Laetolil and Upper Ndolanya Beds, as well as within the Upper Laetolil Beds sequence.
Shifting habitat availability would have had implications for the relationships between the Laetoli mammals, including hominins, and their environment. Early in their history hominins were likely restricted to a vegetarian diet and comprised part of the general herbivore guild prior to their adoption of a diet based partially on animal resources when they came into competition with other carnivores. Additionally, a change in the amount of tree cover — which is used by many woodland adapted mammals not just to obtain dietary staples, but as a place to sleep and seek refuge — would have been a modification to the spatial niches available to our ancestors. In adapting to the selective pressures of a reduced woodland component in their environments, hominins evolved a greater reliance on terrestrial dietary resources and greater efficiency in terrestrial, upright locomotion.
Many other forms of data (such as fossil wood, pollen, isotopic signatures) can and have been used to approach palaeoecological questions, but evidence from the mammalian communities present at Laetoli has been particularly useful in corroborating other interpretations of hominin palaeoecology. Analyses of many African Plio-Pleistocene sites suggest that Australopithecus afarensis lived in woodland and bushland habitats much like those of the Upper Laetolil Beds and that Paranthropus species inhabited a similar mosaic on the more open end of this spectrum, like we see in the Upper Ndolanya Beds (Reed 1997). Palaeoclimatic data suggests that a large-scale trend towards greater habitat variability and aridity began approximately 2.8 mya (deMenocal 2004); the Ndolanya mammals may represent a community that had adapted to resultant changes in available ecological niches.References and Recommended Reading
Books:
Harrison, T., Ed. Paleontology and Geology of Laetoli: Human Evolution in Context. Volumes 1 & 2. Dordrecht: Springer, 2011.
Kingdon, J. The Kingdon Field Guide to African Mammals. London: Academic Press, 1997.
Leakey, M.D. & Harris, J.M, Eds. Laetoli: A Pliocene Site in Northern Tanzania. Oxford: Clarendon Press, 1987.
Ndessokia, P.N.S. The Mammalian Fauna and Archaeology of the Ndolanya and Olpiro Beds, Laetoli, Tanzania. PhD dissertation, University of California at Berkeley.
Journal Articles:
Andrews, P.J. Palaeoecology of Laetoli. Journal of Human Evolution 18, 173-181 (1989).
deMenocal, P.B. African climate change and faunal evolution during the Pliocene-Pleistocene. Earth and Planetary Science Letters 220, 3-24 (2004).
Kovarovic, K. & Andrews, P. Bovid postcranial ecomorphological survey of the Laetoli paleoenvironment. Journal of Human Evolution 52, 663-680 (2007).
Kovarovic, K., Andrews, P. & Aiello, L. The palaeoecology of the Upper Ndolanya Beds at Laetoli, Tanzania. Journal of Human Evolution 43, 395-418 (2002).
Reed, K.E. Early hominid evolution and ecological change through the African Plio-Pleistocene. Journal of Human Evolution 32, 289-322 (1997).
Selections in an Edited Volume:
Bishop, L.C., Plummer, T.W., Hertel, F. & Kovarovic, K. Paleoenvironments of Laetoli, Tanzania as determined by antelope habitat preferences. In Paleontology and Geology of Laetoli: Human Evolution in Context. Ed. Harrison, T. (Dordrecht: Springer, 2011). 355-366.
Su, D.F. Large mammal evidence for the paleoenvironment of the Upper Laetolil and Upper Ndolanya Beds of Laetoli, Tanzania. In Paleontology and Geology of Laetoli: Human Evolution in Context. Ed. Harrison, T. (Dordrecht: Springer, 2011). 381-392.









