


















« Prev Next »

The molecular evolutionary rate measures the frequency with which DNA or protein sequence mutations are fixed (i.e., shared by most individuals) in a population. On the other hand, the mutation rate refers to the amount of change in a DNA or protein sequence for a given unit of time. These two intrinsically related processes have been key to our basic understanding of molecular evolution. For example, the molecular clock theory (Zuckerkandl & Pauling 1962) predicts an increase in fixed amino acid mutations of protein sequences as a function of time. The molecular evolutionary rate is then expected to be constant, assuming that mutation rates are the same across the evolutionary lineages of a given protein (Figure 1). The neutral theory of molecular evolution (Kimura 1983) explains this pattern by proposing that most mutations do not have an effect on the fitness of an organism. Under this model, fixed DNA nucleotide mutations that do not result in changes to protein sequences are referred to as neutral or synonymous substitutions. Also, beneficial mutations are expected to be rare and deleterious ones quickly removed by natural selection. Although the molecular clock and neutral theories are foundational to modern molecular biology, they do not always explain why evolutionary rates vary among genes, proteins, and species. As a result, biologists have begun to consider how organismal-level traits such as life history could influence variation in molecular evolution across the tree of life. This is a central pursuit in evolutionary biology as it is relevant to key concepts such as speciation and the ability of an organism to adapt.
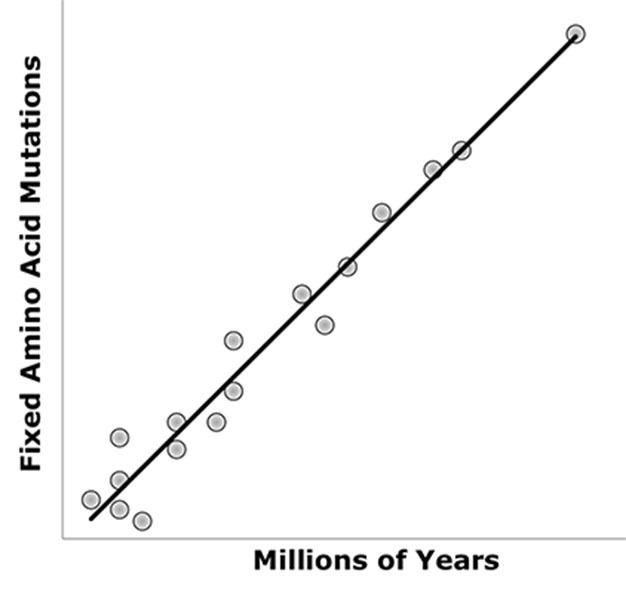
Figure 1: Under a molecular clock, fixed amino acid mutations of a protein sequence (vertical axis) increase as a function of time (hotizonal axis).
Thus, the molecular evolutionary rate is expected to be constant (bold line). Each dot represents divergent evolutionary lineages of a given protein. Proteins in divergent evolutionary lineages that are not plotted on the line represent deviations from theory. Those to the left of the line are expected to have accelerated rates of evolution, while those to the right are expected to have decreased rates.
© 2013 Nature Education Modified from concepts in Graur & Li 2000. All rights reserved. 
Molecular Evolution and Life history
What determines variation in the mutation rate and ultimately the molecular evolutionary rate? The answer to this question is debatable for a number of reasons. Evidence at the cell and molecular levels clearly supports the finding that mutations often derive from DNA replication errors or mutagens in the environment. Organismal-level traits and population genetic processes, in turn, could influence the frequency of DNA replication errors and mutagen levels. Mutations are transmitted from generation to generation in the germ cell line of sexually reproducing organisms. Because DNA in germ cells replicates during meiosis (gamete differentiation), we expect short-lived species to have higher mutation rates. For example, mice reproduce frequently and have short generation times. Their germ cell lines undergo more rounds of meiosis, thereby increasing the chances of DNA replication errors. Compared to long-lived species, mice also are expected to have larger populations with more individuals that are available to reproduce — that is, a larger effective population size. Consequently, synonymous (i.e., neutral) mutations are more likely to occur, leading to higher molecular evolutionary rates. Furthermore, the larger effective population size promotes selection against non-synonymous mutations that reduce fitness, thereby removing them from the population. Clearly, the nucleotide chains that comprise the nuclear and organellar genomes of eukaryotic cells do not evolve independently of organismal-level traits such as life history.
Broadly speaking, life history refers to traits that function to regulate the life cycles of species (Roff 2002) (Figure 2). Examples of such traits include the timing of reproductive effort or its magnitude (fecundity). When is the best time to reproduce for an organism in a given habitat? How often does reproduction take place? These life-history traits are interrelated, making their potential effects on molecular evolutionary rates difficult to discern. However, biologists have proposed two models that establish a clear link between life history and molecular evolution: the generation time and metabolic rate hypotheses. The generation time hypothesis, illustrated in the mouse example, is related to the metabolic rate hypothesis because short-lived species with smaller body size usually have higher metabolic rates. Specifically, the metabolic rate hypothesis predicts that species with high metabolic rates have increased mutation rates due to mutagenic elements resulting from mitochondrial respiration (Galtier et al. 2009). Sometimes free oxygen radicals generated during respiration escape the mitochondrial electron transport chain that produces energy for the cell. These molecules cause damage to nearby mitochondrial DNA by oxidizing nucleotides.
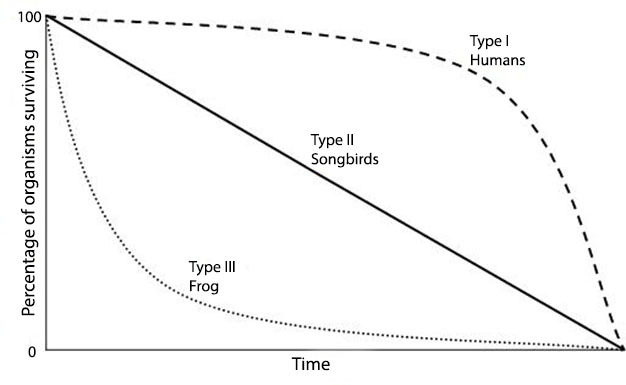
Figure 2: Organisms employ reproductive strategies that are adapted to their habitats.
In this comparison, humans have a Type I survivorship curve in which a high percentage of offspring survive to adulthood and generation time is long. Frogs feature an opposite pattern (Type III) in which high numbers of offspring are produced but few survive. The small percentage that survives reaches adulthood quickly to produce high offspring numbers. Birds display an intermediate strategy (Type II).
Photo courtesy of Ray Husthwaite.
To determine whether life-history traits such as generation time or metabolic rate affect molecular evolutionary rates, it is necessary to make comparisons across multiple evolutionary lineages. Molecular evolutionary rates and life-history differences among taxonomic groups are contrasted after accounting for shared common ancestry. The rate of molecular evolution is measured by using computer algorithms that estimate the number of substitutions (fixed mutations) per unit time in DNA sequences. This process is aided by using adequately dated fossils and well-supported hypotheses on the evolutionary relationship of species. We will briefly explore recent case studies that have adopted such approaches. Some of the most striking examples that illuminate the relationship between life history and molecular evolutionary rates are from plant and animal genomes.
Life History and Molecular Evolution in Plants
Differences in life-history strategies within taxonomic groups of flowering plants appear to influence molecular evolutionary rates (Figure 3). A comprehensive phylogenetic assessment of angiosperms found that rates of DNA sequence (chloroplast + mitochondrial + nuclear) evolution in long-lived (perennial) species are low compared to those of short-lived (annual) species (Smith & Donoghue 2008). This finding was consistent with the generation time hypothesis, and the results were subsequently validated by genome-wide assessments on model plant species. However, correlations were stronger in nuclear genomes as these featured higher molecular evolutionary rates than those of chloroplasts (Yue et al. 2010). Even when taking into account that some annual plant species are self-breeding, generation time still emerges as the strongest life-history correlate of molecular evolutionary rates (Muller & Albach 2010). Overall, the generation time hypothesis is the best-supported life-history model that explains molecular evolutionary rate variation in plants. There is little evidence to support the metabolic rate hypothesis in plants, although mitochondrial evolutionary rates are generally lower compared to those of animals.

Figure 3: Difference in the molecular evolutionary rate of annual (herbs) vs. perennial (trees) plants.
© 2013 Nature Education (tree) Courtesy of Cordero. (flower) Photo via Wikimedia Commons. All rights reserved.
Life History and Molecular Evolution in Animals
Among mammals, mitochondrial evolutionary rates range from one substitution per 1-2 million years to one substitution per more than 100 million years — a difference of 2 orders of magnitude (Nabholz et al. 2008A). Following the assumption that animal body size is correlated with metabolism, theoretical models strongly support the idea that molecular evolutionary rate variation is influenced by basal metabolic rate (Gillooly et al. 2005). In practice, support for this hypothesis is limited because few studies have addressed mutations in the germ cell line (Galtier et al. 2009). Germ cell line mutation rate was assessed in mutant strains of the nematode Caenorhabditis elegans with deficiencies in the mitochondrial electron transport chain. However, this experimental approach did not provide evidence for an increase in mitochondrial DNA mutation due to metabolic oxidative stress (Joyner-Matos et al. 2011).
The relationship between life history and molecular evolutionary rates is more apparent in animal nuclear genomes. While a study on the effect of 14 life-history traits on molecular evolutionary rates of mitochondria in mammals did not provide strong support for the generation time hypothesis (Nabholz et al. 2008B), life history appears to influence molecular evolution in the nuclear DNA of mammals (Bazin et al. 2006). Evidence suggests that their nuclear genomes evolve according to expectations from neutral theory. For example, the nuclear DNA of humans has a lower molecular evolutionary rate compared to primates that have shorter generation times. Specifically, in mammals, neutral evolutionary rates depend on generation time, while non-synonymous rates depend on population size (Nikolaev et al. 2007). Similar patterns have been demonstrated in invertebrate animals with the exception that generation time was correlated with non-synonymous rates (Thomas et al. 2010). These studies further support the generation time hypothesis, but we must keep in mind that it is not mutually exclusive from population size effects.
Does life history drive molecular evolutionary rates?
We have discussed evidence to support the idea that molecular evolutionary rates are driven by life history. By comparing differences among a wide variety of organisms, biologists can test the prediction that DNA nucleotide sequences do indeed evolve according to a rate that, at least partially, depends on organism-level traits. Generation time and metabolism, each to some degree or in combination, affect the mutation rates of some organisms and, thus, their molecular evolutionary rates. Even so, some relationships among generation time, metabolism, and molecular evolution depend on whether the organism is a plant or an animal and the location of the genome within the cell (i.e., nuclear vs. organellar). Also, differences in neutral vs. non-synonymous rates, when averaged together across long DNA sequences, could further complicate interpretations.
Our understanding of variation in molecular evolutionary rates is likely to improve when more knowledge from molecular and organismal biology is made available. It is important to point out that the drivers of variation in molecular evolutionary rate even among genomic regions of a single species are not entirely understood. Work focusing on hypotheses concerned with DNA repair mechanisms, environmental effects, gene duplication, hypermutable nuclear DNA regions, population genetics, and sex-specific mutation bias may provide additional insights as to why rates of molecular evolution vary within and among species. An appreciation of these processes is necessary to clarify the origins of biological diversity and other evolutionary phenomena.References and Recommended Reading
Bazin, E. et al. Population size does not influence mitochondrial genetic diversity in animals. Science 312, 570-572 (2006).
Galtier, N. et al. Mitochondrial whims: Metabolic rate, longevity and the rate of molecular evolution. Biology Letters 5, 413-416 (2009).
Gillooly, J. F. et al. The rate of DNA evolution: effects of body size and temperature on the molecular clock. Proceedings of the National Academy of Sciences (USA) 102, 140-145 (2005).
Graur, D., & Li, W.-H. Fundamentals of Molecular Evolution. Sunderland, MA: Sinauer Associates, 2000.
Joyner-Matos, J. et al. No evidence of elevated germline mutation accumulation under oxidative stress in Caenorhabditis elegans. Genetics 189, 1439-1447 (2011).
Kimura, M. The Neutral Theory of Molecular Evolution. Cambridge, UK: Cambridge University Press, 1983.
Muller, K., & Albach, D.C. Evolutionary rates in Veronica L. (Plantaginaceae): Disentangling the influence of life history and breeding system. Journal of Molecular Evolution 70, 40-56 (2010).
Nabholz, B. et al. Strong variations of mitochondrial mutation rate across mammals-the longevity hypothesis. Molecular Biology and Evolution 25, 120-130 (2008A).
Nabholz, B. et al. Determination of mitochondrial genetic diversity in mammals. Genetics 178, 351-361 (2008B).
Nikolaev, S.I. et al. Life-history traits drive the evolutionary rates of mammalian coding and noncoding genomic elements. Proceedings of the National Academy of Sciences (USA) 104, 20443-20448 (2007).
Roff, D. Life History Evolution. Sunderland, MA: Sinauer Associates, 2002.
Smith, S. A., & Donoghue, M. J. Rates of molecular evolution are linked to life history in flowering plants. Science 322, 86-89 (2008).
Thomas, J. A. et al. A generation time effect on the rate of molecular evolution in invertebrates. Molecular Biology and Evolution 27, 1173-1180 (2010).
Yue, J.-X. et al. Genome-wide investigation reveals high evolutionary rates in annual model plants. BMC Plant Biology 10, 242 (2010).
Zuckerkandl, E. & Pauling, L. B. "Molecular disease, evolution, and genetic heterogeneity," in Horizons in Biochemistry, eds. M. Kasha & B. Pullman.(New York: Academic Press, 1962)189-225.









