
Abstract
Aspergillus species are emerging causative agents of non-dermatophyte mold onychomycosis. In this study, 48 Aspergillus isolates were obtained from patients with onychomycosis in Mashhad, Iran, during 2015–2018. The aim is to identify the Aspergillus isolates to the species level by using partial calmodulin and beta-tubulin gene sequencing and MALDI-TOF MS, and to evaluate their in vitro susceptibility to ten antifungal drugs: terbinafine, itraconazole, voriconazole, posaconazole, ravuconazole, isavuconazole, caspofungin, micafungin, anidulafungin and amphotericin B according to CLSI M38-A3. Our results indicate that A.flavus (n = 38, 79%) is the most common Aspergillus species causing onychomycosis in Mashhad, Iran. Other detected species were A. terreus (n = 3), A. tubingensis (n = 2), A. niger (n = 1), A. welwitschiae (n = 1), A. minisclerotigenes (n = 1), A. citrinoterreus (n = 1) and A. ochraceus (n = 1). Aspergillus flavus, A. terreus and A. niger isolates were correctly identified at the species level by MALDI-TOF MS, while all cryptic species were misidentified. In conclusion, A. flavus is the predominant Aspergillus species causing onychomycosis due to Aspergillus spp. in Mashhad, Iran. MALDI-TOF MS holds promise as a fast and accurate identification tool, particularly for common Aspergillus species. It is important that the current database of reference spectra, representing different Aspergillus species is expanded to increase the precision of the species-level identification. Terbinafine, posaconazole and echinocandins were in vitro most active against the studies Aspergillus isolates and terbinafine could be the first choice for treatment of onychomycosis due to Aspergillus.
Similar content being viewed by others


Introduction
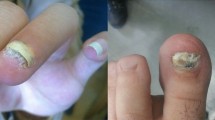
Onychomycosis is a common fungal infection of the nail and is mainly caused by dermatophytes. However, the incidence of onychomycosis due to yeasts and non-dermatophyte molds (NDMs) is increasing1,2. Aspergillus species have been reported as causal agents of non-dermatophyte mold onychomycosis (NDMO)2,3. Bongomin et al. reviewed 42 epidemiological studies globally and found that more than 50% (23/42) of the studies reported Aspergillus species as infectious agents of onychomycosis, accounting for 50–100% of the NDMs4.
Aspergillus flavus is most frequently isolated as causal agent of NMDO in Iran1,2,5,6,7. Due to the non-specific clinical presentation of onychomycosis caused by Aspergillus, clinical diagnosis always requires confirmation in a (mycological) laboratory by using direct microscopy, culturing and/or molecular identification4. Most of the published studies on NDMO provided diagnosis only based on morphological identification of the isolates, and few reports included data on antifungal activity. In fact, distinguishing closely related Aspergillus species based on morphology is difficult or even impossible, whereas a correct identification to species level is critical for optimal antifungal treatment because Aspergillus species can have variable drug susceptibility8.
Partial calmodulin (CaM) and beta-tubulin (BenA) gene sequences are currently used for species identification within a given Aspergillus species complex9. Recent studies have demonstrated that matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) could be an useful alternative for sequence-based identification of Aspergilli10,11, particularly for phenotypically similar or indistinguishable Aspergillus species11,12.
In this study, partial CaM and BenA gene sequences in combination with MALDI-TOF MS analysis are used to identify 48 Aspergillus isolates obtained from onychomycosis patients in two hospitals of Mashhad, Iran. Furthermore, antifungal susceptibility testing on those isolates to 10 antifungal drugs including terbinafine is performed using CLSI M38-A3.
Materials and methods
Isolation
A total 48 clinical Aspergillus isolates were collected from patients diagnosed with onychomycosis during the period from 2015 to 2018 at Emam Reza and Ghaem hospitals in Mashad, Iran. Diagnosis of Aspergillus onychomycosis was made based on dystrophic nail appearance and mycological criteria following literature4: (1) positive direct microscopy with hyphae presented in nail specimen; (2) positive culture of NDM; (3) repeated culturing of the NDM with absence of dermatophytes and yeasts at different time and samples; (4) molecular identification of the Aspergillus isolates.
All specimens were collected using a scalpel and direct microscopic examination was performed using a 20% KOH solution. Sabouraud Glucose Agar (SGA) (Difco, Detroit, MI, USA) containing chloramphenicol (50 mg/L) and cycloheximide (400 mg/L) was used for the isolation of dermatophytes and SGA with chloramphenicol (50 mg/L) for molds. All isolation plates were incubated at 27 °C for up to 4 weeks.
Sequenced-based and MALDI-TOF MS identification
The isolates were identified by CaM and BenA gene sequencing9,13. The obtained sequences were compared to the NCBI nucleotide database (BLAST; http://blast.ncbi.nlm.nih.gov/Blast.cgi) and the internal sequence database of the Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands, containing verified CaM and BenA gene sequences of all accepted Aspergillus species14.
MALDI-TOF MS was performed by the formic acid extraction method according to the manufacturer’s instruction (AUTOF MS1000, Autobio, China) and a previous publication with minor modification15. All used chemical reagents were of LC–MS grade. Briefly, each Aspergillus isolate was cultured on SGA for 3–5 days at 35 °C. After growth, an appropriate amount of sample was collected in a 1.5 mL centrifuge tube containing 1.0 mL 75% ethanol (Sigma-Aldrich, St.Louis, MO, USA). After mixing, the sample was centrifuged at 15,000 × g for 5 min and the supernatant discarded. After drying of the residue at 37 °C, 40 µL of lysis solution 1 (containing formic acid) was added and air-dried at room temperature. Subsequently, 1µL was transferred on a target plate, dried naturally in a bio-safety cabinet, and afterwards 1µL of matrix solution was added and dried again. Eventually, for each isolate a mass spectrum was generated and integrated to give a sum spectrum using AUTOF MS1000 (Autobio, China) with in house database.
Species identification by CaM and BenA gene sequencing and MALDI-TOF MS of 48 Aspergillus isolates are shown in Table 1. GenBank accession numbers for BenA and CaM gene sequences generated in this study are listed in Suppl. Table 1.
Antifungal susceptibility testing and statistical analysis
All isolates were tested according to Clinical and Laboratory Standards Institute (CLSI) M38-A3 document16. The ten antifungal agents included in this study are itraconazole, voriconazole, posaconazole, ravuconazole, isavuconazole, caspofungin, anidulafungin, amphotericin B (Sigma, Poole, United Kingdom), terbinafine (Aladdin, California, United States) and micafungin (Toronto-Research-Chemicals, Toronto, Ontario, Canada). The antifungal agents were tested at concentrations ranging from 0.008–4 mg/L for echinocandins (micafungin, anidulafungin and caspofungin) and 0.031–16 mg/L for the other compounds. Briefly, all Aspergillus isolates were grown on Potato Dextrose Agar (PDA) at 35 °C for 4–5 days to induce sporulation. Conidia were harvested using sterile saline with grinding, and the final inoculum concentration of the suspension was adjusted to 0.4–5 × 104 colony-forming units (CFU) per mL in RPMI 1640 buffered with morpholinepropanesulfonic acid. The plates for echinocandins were incubated at 35 °C for 24 h, and the other plates were incubated at 35 °C for 48 h. Both minimum inhibitory concentrations (MIC) and minimum effective concentrations (MEC) were determined microscopically (Olympus, Japan) at × 40 magnification. Aspergillus fumigatus ATCCMYA-3627 and Candida parapsilosis ATCC-22019 were used as quality control. MIC/MEC ranges, Geometric mean MIC/MEC, Modal MIC/MEC were calculated for all isolates.
All methods were performed in accordance with the relevant guidelines and regulations as references were given.
Ethics approval
Ethical approval to conduct the study was obtained from the Ethics Committee of Mashhad University of Medical Sciences (IR.MUMS.MEDICAL.REC.1397.660) and all patients involved understood and agreed to the use of these clinical specimens in the present study.
Results
Identification
All 48 isolates were identified based on morphology in combination with BLAST analysis of the generated BenA and CaM sequences. Gene sequencing revealed presence of eight species (Table 1) belonging to four Aspergillus sections, and included four non-cryptic species (A. flavus, A. niger, A. terreus and A. ochraceus) and four cryptic species (A. minisclerotigenes, A. citrinoterreus, A. tubingensis and A. welwitschiae). Thirty-nine (81%) isolates belonged to section Flavi (A. flavus, n = 38; A.minisclerotigenes, n = 1), four (8%) to section Terrei (A. terreus, n = 3; A. citrinoterreus, n = 1), four (8%) to section Nigri (A. tubingensis, n = 2; A. niger, n = 1; A. welwitschiae, n = 1) and one isolate was identified as A. ochraceus (section Circumdati).
An 89.5% concordance between MALDI-TOF MS and molecular identification was found MALDI-TOF MS allowed the identification of the four common non-cryptic species, but failed to correctly identify the four cryptic species.
Antifungal susceptibility
The ranges MIC/MEC, Geometric mean MIC/MEC, Modal MIC/MEC, distribution of MICs/MECs of ten antifungal agents against 48 Aspergillus isolates are presented in Table 2.
In general, among 48 Aspergillus isolates tested in this study, the lowest Modal MIC/MEC (< 0.008 mg/L) were those of anidulafungin and micafungin, followed by terbinafine and posaconazole (0.031 mg/L). The other four azole compounds tested were active in vitro, with MICs of ≤ 1 mg/L. Posaconazole and caspofungin revealed variable MIC/MEC values with MIC ranges from 0.031 to 0.5 mg/L and MEC 0.063–0.25 mg/L, respectively.
The 38 A. flavus isolates showed relatively high MIC value to amphotericin B (range: 1–4 mg/L, Modal MIC: 2 mg/L, n = 24), but lower MICs than the epidemiological cutoff value (ECV) (1 mg/L) for all azoles tested. When testing the susceptibility to terbinafine, all A. flavus isolates revealed low MIC with Modal MIC 0.031 mg/L (n = 35). A similar MIC value is observed for the A. minisclerotigenes isolate.
For section Terrei, three A. terreus isolates had a similar antifungal susceptibility profile as A. flavus, except terbinafine, which has higher MICs (Modal MIC 0.25 mg/L, n = 3). In contrast to A. terreus, the isolate of the cryptic species A. citrinoterreus had a MIC of 4 mg/L to amphotericin B.
All section Nigri isolates revealed lower MIC and Modal MIC ≤ 1 mg/L to the five azoles tested. The majority isolates of section Nigri have an MIC value of 0.031 mg/L to terbinafine except one A. tubingensis isolate with an MIC value of 0.5 mg/L.
Discussion
This study presented, for the first time, an overview of the occurrence of Aspergillus species causing onychomycosis in Mashhad, Iran, including a molecular characterization of the isolates and in vitro susceptibility to 10 antifungal agents. In total 48 Aspergillus isolates were obtained from nail in patients with onychomycosis in two medical centers of Mashhad, Iran.
Our study highlights the epidemiological features that A. flavus is the predominant species causing NDMO due to Aspergillus. in Mashhad, Iran. This finding is similar to those reported from other places in Iran1,6,7,17,18 and differs from Sri Lanka19, Cameroon20, India21,22, Turkey23 where A. niger is the most frequently species, and from Italy24 and UK25 where A. fumigatus and A. terreus are the most common agents, respectively. Data from Iran suggest that A. flavus is most common agent involved in NDMO5,6,7. Besides onychomycosis, A. flavus is also the leading cause of chronic fungal rhinosinusitis in Iran26. This could be due to the arid climate in Iran that favors the growth of thermo-tolerant fungi like A. flavus. Aspergillus tubingensis and A. welwitschiae (former name A. awamorii) in section Nigri have previously been reported as emerging causal agents of onychomycoses2,27. Both species have been reported as the causal agents of otomycoses as well28,29,30. In our study, four cryptic species were identified based on the analysis of BenA and CaM gene sequences: A. minisclerotigenes in section Flavi, A. citrinoterreus in section Terrei and A. tubingensis and A. welwitschiae in section Nigri. To our knowledge, A. minisclerotigenes, A. citrinoterreus and A. ochraceus are for the first time identified as causative agents of NDM onychomycosis. Aspergillus minisclerotigenesis is closely related to A. flavus31. Dehghan et al.32 reported a human infection caused by A. minisclerotigenes from Iran in 2014 and Esfahani et al.33 reported the first case of fungal keratitis due to this species in Iran in 2019, suggesting that the occurrence of A. minisclerotigenes might have been underreported due to identification based on morphology. The recently described A. citrinoterreus within section Terrei34 has been reported as causative agent involved inhuman invasive aspergillosis35,36,37. Aspergillus ochraceus belongs to Aspergillus section Circumdati. This species has rarely been related to human infection38,39. So far, only two cases reported A. ochraceus as a pathogenone case was an invasive pulmonary aspergillosis from Poland38, another case was allergic bronchopulmonary aspergillosis39.
In our study, 38 isolates were molecularly identified as A. flavus or A. oryzae. These two species are genetically very similar and indistinguishable by BenA and CaM gene sequencing40. Identification of these species is based on the origin and/or toxin production potential: A. oryzae is used in (food) fermentations and biotechnology and does not produce aflatoxins, while A. flavus is not domesticated and can produce aflatoxins, though non-toxigenic strains also occur41.Of note, all 38 A. flavus isolates were corrected identification by MALDI-TOF MS analysis based on the AUTOF MS 1000 (Autobio, China) in-house database. Similarly, De Carolis et al.40 reported that MALDI-TOF MS easily differentiated A. flavus and A.oryzae on the species level. In contrast to some investigators42,43, who reported that only 18 of 200 isolates were confirmed as A. flavus using MALDI-TOF MS based on the Bruker score database among a set of 200 clinical and environmental A. flavus isolates identified by sequencing. Masih et al. 43 also showed that A. oryzae can easily be identified as A. flavus. Those discrepancy could be due to differences between MALDI-TOF MS machines, the species coverage in reference databases, interpretive cutoffs and methodology applied for sample preparation11.
In our study, A. minisclerotigenes was misidentified as A. flavus by MALDI-TOF MS. Similarly, A. tubingensis and A. welwitschiae were misidentified as A. niger and A. citrinoterreus was misidentified as A. terreus. These are due to the inadequate number of reference spectra in the used in-house database (Suppl. Table 2). Therefore, databases should be expanded with well-validated spectra of cryptic species in order to get an accurate identification of all (clinical relevant) Aspergillus species. Our study confirms the discriminatory power of MALDI-TOF MS for common clinical Aspergillus species40.
The antifungal susceptibility data generated in this study indicate that terbinafine has excellent in vitro activity against the eight species tested. Terbinafine appears more active against isolates of section Flavi (GM 0.032 mg/L and section Nigri (GM 0.077 mg/L) than to section Terrei isolates (GM 0.25 mg/L), and has superior activity against A. flavus compared to the azoles and amphotericin B. Similar results are obtained in previous studies44,45. Thus, terbinafine could be placed as an alternative drug for treatment of onychomycosis caused by Aspergillus species with the confirmation of clinical efficacy of terbinafine in the management of onychomycosis due to Aspergillus spp.4.
Based on the proposed epidemiologic cut-off values (ECV) of A. flavus (posaconazole 0.5 mg/L; itraconazole 1 mg/L; voriconazole 1 mg/L; isavuconazole1 mg/L and amphotericin B 4 mg/L)46, all azoles tested in our study exhibited good activity to 38 A. flavus isolates and this is in agreement with previous reports18,19. Posaconazole had the lowest GM value (0.097 mg/L), followed by voriconazole and isavuconazole (both 0.274 mg/L), itraconazole and ravuconazole (0.311 and 0.394 mg/L, respectively) in an increasing order. Although no antifungal susceptibility ECV are available for A. flavus to ravuconazole, in our study, all A. flavus isolates had MIC values less than 1 mg/L (MIC range 0.25–0.5 mg/L), indicating the potency of this antifungal against A. flavus, which is in agreement with those reported by Pfaller et al.47. Amphotericin B showed relatively high MICs (GM 2.151 mg/L; MIC range 1–4 mg/L) against all A. flavus isolates, including a MIC of 4 mg/L for nine (23.6%) isolates of A. flavus, which is similar to previous reports from the United States, Europe48,49, and the Middle East50,51. The A. minisclerotigenes isolate had similar MIC values as A. flavus.
Among the section Nigri isolates, the azoles tested were active against all isolates, although an A. tubingensis isolate presented a slightly higher MIC value for azoles, which is similar to results reported previously52.
The three A. terreus and one A. citrinoterreus isolate displayed low MICs for the tested azoles. Posaconazole was the most effective azole against A. terreus (modal MIC 0.063 mg/L, n = 2) which is similar to previous studies53,54.
Although A terreus is intrinsically resistant to amphotericin B, 12–13% of the isolates have low amphotericin B MICs55,56. Our results also show that the three A. terreus isolates exhibited MICs < 2 mg/L, which is below the proposed ECV (MIC 4 mg/L). However, the A. citrinoterreus isolate had a MIC of 4 mg/L for amphotericin B, which is in agreement with those reported by Imbert et al.35.
Echinocandins showed good activity against the most Aspergillus isolates in this collection, while anidulafungin and micafungin showed a lowest Modal MEC of 0.008 mg/L, followed by caspofungin with Modal MEC of 0.125–0.5 mg/L. Anidulafungin and micafungin appeared more potent than caspofungin and this is consistent with several previous studies44,57. Of note, the four section Terrei isolate have higher MEC values (0.5 mg/L), above ECV (0.125 mg/L). The similar results were reported by Lass-Flörl et al. With a set of 48 clinical and 31 environmental A. terreus isolates, they showed that caspofungin has higher MECs (MEC90 2 mg/L) than anidulafungin (MEC90 0.03 mg/L) and micafungin (MEC90 0.02 mg/L)58.
Conclusion
Based on molecular and MALDI-TOF MS identification, A. flavus (79%) is the most common Aspergillus species in NDM onychomycosis due to Aspergillus in Mashhad, Iran. The other isolates showed a wider species diversity. We emphasize the importance of using molecular methods to accurately identify Aspergillus at the species level because different species may vary in terms of susceptibility to antifungal agents. However, our results are limited by the relatively low number of clinical Aspergillus isolates obtained in Mashhad, Iran. Terbinafine, posaconazole, and echinocandins are shown in vitro to be the most potent antifungal agents against Aspergillus spp. Terbinafine could be first line drug for treatment of onychomycosis due to Aspergillus, the in vivo efficacy remains to be determined.
Data availability
The samples utilized in our study were obtained from preexisting samples isolated from patients who routinely referred to the hospital for direct examination and culture in Clinical Lab. And we used the culture for this research, and no additional samples were taken from the patient.
References
Motamedi, M. et al. Growing incidence of non-dermatophyte onychomycosis in Tehran, Iran. Jundishapur J. Microbiol. 9, e40543. https://doi.org/10.5812/jjm.40543 (2016).
Nouripour-Sisakht, S. et al. Aspergillus species as emerging causative agents of onychomycosis. J. mycol. Med. 25, 101–107. https://doi.org/10.1016/j.mycmed.2014.12.001 (2015).
Zotti, M., Agnoletti, A. F., Vizzini, A., Cozzani, E. & Parodi, A. Onychomycosis from Aspergillus melleus, a novel pathogen for humans. Fungal identification and in vitro drug susceptibility. Exp. Dermatol. 24, 966–968. https://doi.org/10.1111/exd.12807 (2015).
Bongomin, F., Batac, C. R., Richardson, M. D. & Denning, D. W. A Review of onychomycosis due to aspergillus species. Mycopathologia 183, 485–493. https://doi.org/10.1007/s11046-017-0222-9 (2018).
Chadeganipour, M., Nilipour, S. & Ahmadi, G. Study of onychomycosis in Isfahan, Iran. Mycoses 53, 153–157. https://doi.org/10.1111/j.1439-0507.2008.01679.x (2010).
Hashemi, S. J. et al. Onychomycosis in Tehran: mycological study of 504 patients. Mycoses 53, 251–255. https://doi.org/10.1111/j.1439-0507.2009.01703.x (2010).
Mikaeili, A. & Karimi, I. The incidence of onychomycosis infection among patients referred to hospitals in Kermanshah province, Western Iran. Iran. J. Public Health 42, 320–325 (2013).
Ostrosky-Zeichner, L. Invasive mycoses: diagnostic challenges. Am. J. Med. 125, S14-24. https://doi.org/10.1016/j.amjmed.2011.10.008 (2012).
Samson, R. A. et al. Phylogeny, identification and nomenclature of the genus of Aspergillus. Stud. Mycol. 78, 141–173. https://doi.org/10.1016/j.simyco.2014.07.004 (2014).
Sanguinetti, M. & Posteraro, B. MALDI-TOF mass spectrometry: any use for Aspergilli?. Mycopathologia 178, 417–426. https://doi.org/10.1007/s11046-014-9757-1 (2014).
Sanguinetti, M. & Posteraro, B. Identification of Molds by Matrix-Assisted Laser Desorption Ionization-Time ofipaon, M., Torres-Sanchez, M. J. & Aznar, J. Identification of clinical isolates of Aspergillus, including cryptic species, by matrix assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS). Med. Mycol. 56, 838–846, https://doi.org/10.1093/mmy/myx115 (2018).
Siqueira, J. P. Z. et al. Species of asp flight mass spectrometry. J. Clin. Microbiol. 55, 369–379. https://doi.org/10.1128/jcm.01640-16 (2017).
Vidal-Acuna, M. R. Ruiz-Perez de Pergillus section Aspergillus from clinical samples in the United States. Med. Mycol. 56, 541–550. https://doi.org/10.1093/mmy/myx085 (2018).
J.Houbraken. et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): an overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 95, 5–169, https://doi.org/10.1016/j.simyco.2020.05.002 (2020).
Cassagne, C. et al. Mould routine identification in the clinical laboratory by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. PLoS ONE 6, e28425. https://doi.org/10.1371/journal.pone.0028425 (2011).
Wayne, P. Reference method for broth dilution antifungal susceptibility testing of filamentous fungi, 3rd ed. Approved standard. CLSI document M38-A3. Clinical and Laboratory Standards Institute (2017).
Sigurgeirsson, B. & Baran, R. The prevalence of onychomycosis in the global population: a literature study. JEADV 28, 1480–1491. https://doi.org/10.1111/jdv.12323 (2014).
Afshar, P., Khodavaisy, S., Kalhori, S., Ghasemi, M. & Razavyoon, T. Onychomycosis in north-East of iran. Iran. J. Microbiol. 6, 98–103 (2014).
Ranawaka, R. R., de Silva, N. & Ragunathan, R. W. Non-dermatophyte mold onychomycosis in Sri Lanka. Dermatol. Online J. 18, 7 (2012).
Nkondjo Minkoumou, S., Fabrizi, V. & Papini, M. Onychomycosis in Cameroon: a clinical and epidemiological study among dermatological patients. Int. J. Dermatol. 51, 1474–1477. https://doi.org/10.1111/j.1365-4632.2012.05509.x (2012).
Adhikari, L., Das Gupta, A., Pal, R. & Singh, T. S. Clinico-etiologic correlates of onychomycosis in Sikkim. Indian J. Pathol. Microbiol. 52, 194–197. https://doi.org/10.4103/0377-4929.48915 (2009).
Das, N. K. et al. A study on the etiological agent and clinico-mycological correlation of fingernail onychomycosis in eastern India. Indian J. Dermatol. 53, 75–79. https://doi.org/10.4103/0019-5154.41651 (2008).
Hilmioglu-Polat, S. et al. Non-dermatophytic molds as agents of onychomycosis in Izmir, Turkey: a prospective study. Mycopathologia 160, 125–128. https://doi.org/10.1007/s11046-005-6872-z (2005).
Gianni, C. & Romano, C. Clinical and histological aspects of toenail onychomycosis caused by Aspergillus spp.: 34 cases treated with weekly intermittent terbinafine. Dermatology 209, 104–110. https://doi.org/10.1159/000079593 (2004).
English, M. P. & Atkinson, R. Onychomycosis in elderly chiropody patients. Br. J. Dermatol. 91, 67–72. https://doi.org/10.1111/j.1365-2133.1974.tb06718.x (1974).
Romano, C., Papini, M., Ghilardi, A. & Gianni, C. Onychomycosis in children: a survey of 46 cases. Mycoses 48, 430–437. https://doi.org/10.1111/j.1439-0507.2005.01161.x (2005).
Tsang, C. C. et al. Genetic diversity of Aspergillus species isolated from onychomycosis and Aspergillus hongkongensis sp nov, with implications to antifungal susceptibility testing. Diagn. Microbiol. Inf. Dis. 84, 125–134. https://doi.org/10.1016/j.diagmicrobio.2015.10.027 (2016).
Szigeti, G. et al. Species assignment and antifungal susceptibilities of black aspergilli recovered from otomycosis cases in Iran. Mycoses 55, 333–338. https://doi.org/10.1111/j.1439-0507.2011.02103.x (2012).
Szigeti, G. et al. Molecular identification and antifungal susceptibilities of black Aspergillus isolates from otomycosis cases in Hungary. Mycopathologia 174, 143–147. https://doi.org/10.1007/s11046-012-9529-8 (2012).
Hagiwara, S. et al. The Molecular Identification and Antifungal Susceptibilities of Aspergillus Species Causing Otomycosis in Tochigi, Japan. Mycopathologia 184, 13–21. https://doi.org/10.1007/s11046-018-0299-9 (2019).
Pildain, M. B. et al. Two novel aflatoxin-producing Aspergillus species from Argentinean peanuts. Int. J. Syst. Evol. Microbiol. 58, 725–735. https://doi.org/10.1099/ijs.0.65123-0 (2008).
Dehghan, P. et al. Multilocus variable-number tandem-repeat analysis of clinical isolates of Aspergillus flavus from Iran reveals the first cases of Aspergillus minisclerotigenes associated with human infection. BMC Infect. Dis. 14, 358. https://doi.org/10.1186/1471-2334-14-358 (2014).
Karimizadeh Esfahani, M., Eslampoor, A., Dolatabadi, S., Najafzadeh, M. J. & Houbraken, J. First case of fungal keratitis due to Aspergillus minisclerotigenes in Iran. Current Med. Mycol. 5, 45–48,https://doi.org/10.18502/cmm.5.2.1162 (2019).
Guinea, J. et al. Aspergillus citrinoterreus, a new species of section Terrei isolated from samples of patients with nonhematological predisposing conditions. J. Clin. Microbiol. 53, 611–617. https://doi.org/10.1128/JCM.03088-14 (2015).
Imbert, S. et al. Species identification and in vitro antifungal susceptibility of Aspergillus terreus species complex clinical isolates from a french multicenter study. Antimicrobial Agents Chemother. 62, doi:https://doi.org/10.1128/AAC.02315-17 (2018).
Risslegger, B. et al. A prospective international Aspergillus terreus survey: an EFISG, ISHAM and ECMM joint study. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases 23, 776 e771–776 e775, https://doi.org/10.1016/j.cmi.2017.04.012 (2017).
Zoran, T. et al. Azole-resistance in Aspergillus terreus and related species: An emerging problem or a rare phenomenon?. Front. Microbiol. 9, 516. https://doi.org/10.3389/fmicb.2018.00516 (2018).
Wierzbicka, M., Podsiadlo, B. & Janczarski, M. Invasive pulmonary aspergillosis caused by Aspergillus ochraceus. Pneumonol. Alergol. Pol. 65, 254–260 (1997).
Novey, H. S. & Wells, I. D. Allergic bronchopulmonary aspergillosis caused by Aspergillus ochraceus. Am. J. Clin. Pathol. 70, 840–843. https://doi.org/10.1093/ajcp/70.5.840 (1978).
De Carolis, E. et al. Species identification of Aspergillus, Fusarium and Mucorales with direct surface analysis by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Microbiol. Inf. 18, 475–484. https://doi.org/10.1111/j.1469-0691.2011.03599.x (2012).
Frisvad, J. C. et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 93, 1–63. https://doi.org/10.1016/j.simyco.2018.06.001 (2019).
Hedayati, M. T. et al. Discrimination of Aspergillus flavus from Aspergillus oryzae by matrix-assisted laser desorption/ionisation time-of-flight (MALDI-TOF) mass spectrometry. Mycoses 62, 1182–1188. https://doi.org/10.1111/myc.13010 (2019).
Masih, A. et al. Identification by molecular methods and matrix-assisted laser desorption ionization-time of flight mass spectrometry and antifungal susceptibility profiles of clinically significant rare Aspergillus species in a referral chest Hospital in Delhi, India. J. Clin. Microbiol. 54, 2354–2364. https://doi.org/10.1128/jcm.00962-16 (2016).
Schmitt, H. J. et al. MIC and fungicidal activity of terbinafine against clinical isolates of Aspergillus spp. Antimicrob. Agents Chemother. 32, 780–781. https://doi.org/10.1128/aac.32.5.780 (1988).
Moore, C. B., Walls, C. M. & Denning, D. W. In vitro activities of terbinafine against Aspergillus species in comparison with those of itraconazole and amphotericin B. Antimicrob. Agents Chemother. 45, 1882–1885. https://doi.org/10.1128/AAC.45.6.1882-1885.2001 (2001).
Wayne, P. Reformance standards for antifungal susceptibility testing of filamentous fungi, 1 st ed. CLSI document M61. Clinical and Laboratory Standards Institute (2017).
Pfaller, M. A., Messer, S. A., Hollis, R. J. & Jones, R. N. Antifungal activities of posaconazole, ravuconazole, and voriconazole compared to those of itraconazole and amphotericin B against 239 clinical isolates of Aspergillus spp. and other filamentous fungi: report from SENTRY Antimicrobial Surveillance Program, 2000. Antimicrobial Agents Chemother. 46, 1032–1037, https://doi.org/10.1128/aac.46.4.1032-1037.2002 (2002).
Colozza, C. et al. In vitro activities of amphotericin B and Am Bisome against Aspergillus isolates recovered from Italian patients treated for haematological malignancies. Int. J. Antimicrob. Agents 39, 440–443. https://doi.org/10.1016/j.ijantimicag.2012.01.013 (2012).
Goncalves, S. S., Stchigel, A. M., Cano, J., Guarro, J. & Colombo, A. L. In vitro antifungal susceptibility of clinically relevant species belonging to Aspergillus section Flavi. Antimicrob. Agents Chemother. 57, 1944–1947. https://doi.org/10.1128/AAC.01902-12 (2013).
Taghizadeh-Armaki, M. et al. Genetic Diversity and In Vitro Antifungal Susceptibility of 200 Clinical and Environmental Aspergillus flavus Isolates. Antimicrobial Agents Chemother. 61, https://doi.org/10.1128/AAC.00004-17 (2017).
Al-Wathiqi, F., Ahmad, S. & Khan, Z. Molecular identification and antifungal susceptibility profile of Aspergillus flavus isolates recovered from clinical specimens in Kuwait. BMC Infect. Dis. 13, 126. https://doi.org/10.1186/1471-2334-13-126 (2013).
Li, Y., Wan, Z., Liu, W. & Li, R. Identification and susceptibility of Aspergillus section nigri in china: prevalence of species and paradoxical growth in response to echinocandins. J. Clin. Microbiol. 53, 702–705. https://doi.org/10.1128/JCM.03233-14 (2015).
Lass-Florl, C. et al. Epidemiology and outcome of infections due to Aspergillus terreus: 10-year single centre experience. Br. J. Haematol. 131, 201–207. https://doi.org/10.1111/j.1365-2141.2005.05763.x (2005).
Espinel-Ingroff, A. et al. Wild-type MIC distributions and epidemiological cutoff values for amphotericin B and Aspergillus spp. for the CLSI broth microdilution method (M38-A2 document). Antimicrobial Agents Chemother. 55, 5150–5154, https://doi.org/10.1128/AAC.00686-11 (2011).
Lass-Flörl, C. et al. Epidemiology and outcome of infections due to Aspergillus terreus: 10-year single centre experience. Br. J. Haematol. 131, 201–207. https://doi.org/10.1111/j.1365-2141.2005.05763.x (2005).
Baddley, J. W., Pappas, P. G., Smith, A. C. & Moser, S. A. Epidemiology of Aspergillus terreus at a university hospital. J. Clin. Microbiol. 41, 5525–5529. https://doi.org/10.1128/jcm.41.12.5525-5529.2003 (2003).
Pfaller, M. A. et al. In vitro susceptibility of clinical isolates of Aspergillus spp. to anidulafungin, caspofungin, and micafungin: a head-to-head comparison using the CLSI M38-A2 broth microdilution method. J Clin. Microbiol. 47, 3323–3325, https://doi.org/10.1128/JCM.01155-09 (2009).
Lass-Florl, C., Alastruey-Izquierdo, A., Cuenca-Estrella, M., Perkhofer, S. & Rodriguez-Tudela, J. L. In vitro activities of various antifungal drugs against Aspergillus terreus: Global assessment using the methodology of the European committee on antimicrobial susceptibility testing. Antimicrob. Agents Chemother. 53, 794–795. https://doi.org/10.1128/AAC.00335-08 (2009).
Acknowledgements
We gratefully acknowledge funding from Suzhou Health and Family Planning Commission (LCZX201728), Suzhou New District (2017Z008) to Shuwen Deng and (2019F002) to Xue Xu, partly by an international joint project in National Natural Science Foundation of China (81720108026). We thank Danyang Hu for excellent assistance on molecular identification of the isolates.
Author information
Authors and Affiliations
Contributions
X.X. and A.N. performed experiments, analyzed data and prepared figures and the manuscript. J.H. and F.A. performed experiments and analyzed data. X.-D.W., R.-F.Z. and H.Z. analyzed data and revised the manuscript. M.J.N and S.W.D. designed experiments, supervised the experiments and finalized the manuscript. All the authors have read and approved the paper. XX. and A.N. are contributed equally to this work.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xu, X., Naseri, A., Houbraken, J. et al. Identification and in vitro antifungal susceptibility of causative agents of onychomycosis due to Aspergillus species in Mashhad, Iran. Sci Rep 11, 6808 (2021). https://doi.org/10.1038/s41598-021-86038-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-86038-z
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
