


















« Prev Next »
The cell cycle allows faithful inheritance of the genetic material from mother to daughter cells. Cell cycle events are controlled tightly by several kinase families including CDKs, Plks, and Aurora kinases. What is interesting about these kinases is that they don't always act alone. They sometimes collaborate with each other, making the regulatory network much more complex but also more accurate.
Phosphorylation Modification: A Universal Way to Turn on/off Signaling Pathways
Why is protein phosphorylation important? Phosphorylation is a universal protein modification in eukaryotic cells, and participates in every aspect of cellular life. The human genome encodes over 500 protein kinases that interact with diverse substrates including other kinases, transcription factors, and regulatory proteins. The cell cycle is no exception, and is tightly regulated by protein phosphorylation. Substrates are phosphorylated in a spatiotemporally specific manner by protein kinases on serine, threonine, or tyrosine residues that can subsequently be recognized by other proteins that contain a "phosphopeptide binding domain." In this way, signaling cascades initiated by protein phosphorylation trigger a series of changes in protein-protein interactions. The assembly or disassembly of protein complexes takes place at a specific time and location within the cell, leading to turning a signaling pathway "on" or "off" (Figure 1).
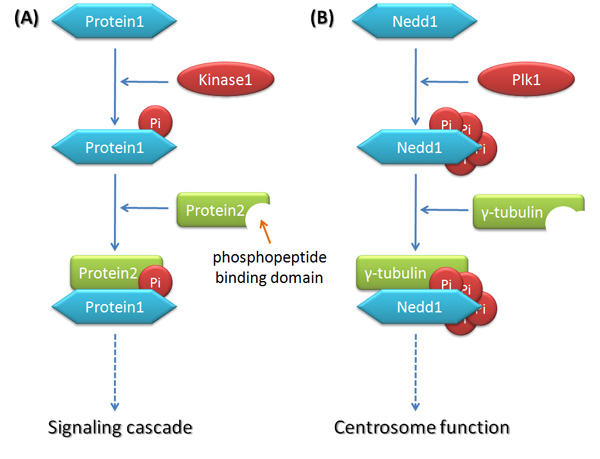
Figure 1: Phosphorylation modification in signaling pathway
(A) Illustrates the general model. Protein1 is phosphorylated by kinase1, and is subsequently recognized by protein 2 which contains the phosphopeptide binding domain. The assembly or disassembly of protein complexes takes place in spatiotemporal manner within the cell, leading to switching the signaling pathways on or off. (B) Illustrates a specific example. During mitosis, Plk1 phosphorylates Nedd1 at several sites, which is further recognized by γ-tubulin. The complex of Nedd1 and γ-tubulin together localizes to the centrosome and enhances the microtubule nucleation around the centrosome.
© 2010 Nature Education All rights reserved. 
Decades ago, scientists discovered cyclin-dependent kinases (CDKs), proteins that act as master regulators of the cell cycle. As their name indicates, the activity of CDKs is controlled by cyclins, other proteins whose expression level oscillates throughout the cell cycle. Different CDKs operate during different phases of the cell cycle and provide the driving force for cell cycle progression. Besides CDKs, scientists have found several protein kinases that also control the cell cycle, primarily Plks (Polo-Like kinases) and Aurora kinases. These kinases control protein phosphorylation in a spatiotemporal manner — meaning they control where and when phosphorylation happens. In fact, these kinases are major regulators of centrosome function, spindle assembly, chromosome segregation, and cytokinesis. Unlike CDKs, which promote cell cycle progression, these kinases are responsible for avoiding errors during cell cycle progression (Figure 2; Archambault & Glover 2009, Hochegger et al. 2008, Fu et al. 2007, Barr et al. 2004, Carmena et al. 2003).
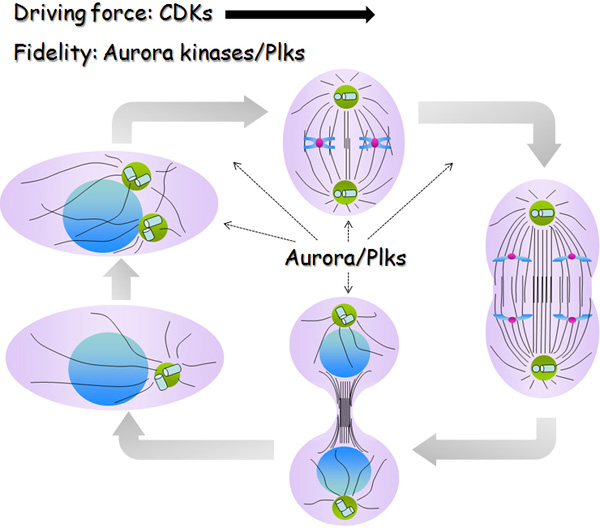
Figure 2: CDKs, Plks and Aurora kinases are important kinase families in cell cycle regulation.
Different CDKs operate at different stages of the cell cycle and together make the driving force to direct the progression. Plks and Aurora kinases exert a fine spatiotemporal control of the cell cycle events in different ways, ranging from centrosome function, spindle assembly, and chromosome segregation to cytokinesis. Their mission is to maintain the fidelity of cell division.
© 2010 Nature Education All rights reserved.
Phosphorylation Pattern: A Kinase versus a Substrate
How do scientists study protein phosphorylation? They use powerful tools developed by structural biologists to understand the mechanics of protein phosphorylation. The interaction between kinase and substrate is quite dynamic, and therefore difficult to follow. But under proper conditions, protein kinases form complexes with peptides derived from substrates. In order to maintain a stable kinase-substrate complex for analysis, scientists use ATPγS (a non-hydrolyzable substrate for ATPases) in a cellular system. This blocks the phosphoryl transfer and the phosphorylation process. With this blockage, the resulting crystal structures of the kinase together with its substrate in active conformations are quite informative. Their structure and binding shapes can be then analyzed by biologists.
Also, we now know that the active conformations of many protein kinases are similar, and so we can classify groups of kinases based on their similar structure. For example, CDK2 and Aurora kinase possess a two-lobe architecture in their kinase domain: the N-terminal lobe and the C-terminal lobe. The active site of these kinases is buried at the interface of these two lobes, which forms a kind of groove, and also includes the ATP binding site and the kinase activation segment (Figure 3; Jeffrey et al. 1995; Fu et al. 2007).
How does the kinase-substrate interaction facilitate phosphorylation? First, the kinase has to properly position the catalytic site and ATP, and then correctly orient the substrate binding site. The kinase itself needs to be phosphorylated at a threonine residue within the activation segment. The docking portion of the substrate then fits into the groove on the surface. Then the kinase transfers a phosphate group from the ATP onto the Ser/Thr/Tyr of the substrate. Protein kinases usually get activated by binding to other entities, called activators. For example, in the absence of the activator TPX2, the activation segment of Aurora A overlaps with the substrate binding site, so that substrate binding is not optimal. With the binding of TPX2, the two lobes of Aurora A kinase domain switch to a conformation that is suitable for substrate binding and phosphoryl transfer. (Fu et al. 2007). Similarly, CDK2 also requires an interaction with Cyclin A to be fully activated (Jeffrey et al. 1995).
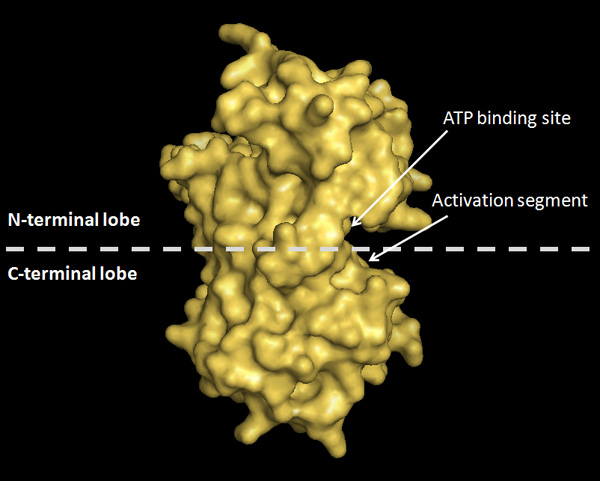
Figure 3: Crystal structure of Aurora A kinase domain to illustrate the common characteristics of protein kinases
The kinase domain of Aurora A is comprised of a two-lobe architecture: N-terminal lobe and C-terminal lobe. The active sites are buried at the interface between two lobes. The arrows indicate the ATP binding site and the kinase activation segment.
© 2010 Nature Education All rights reserved.
Collaboration between Kinases: Sequential Phosphorylation of a Single Substrate
Although scientists have known for some time that CDKs, Plks, and Aurora kinases play central roles in cell cycle regulation, they only recently began to consider possible crosstalk among their three signaling pathways.
CDK1 as a Priming Kinase to Plk1
How was the collaboration among different mitotic kinases discovered? In 2003, scientists devised a proteomic screen that used phosphopeptides derived from substrates of CDKs as bait to identify proteins that were phosphorylated by CDKs. Based on the presumption that protein kinases and phosphopeptide binding domains tend to recognize overlapping sequence motifs, the screen would identify proteins/domains that would preferentially bind to the phosphopeptides that have already been primed — phosphorylated by CDKs (Elia et al. 2003). This approach led to the identification of Plk1 as a phosphopeptide binding domain, which means Plk1 coordinates with CDKs to act on the same substrates. So far, the accumulated data supports the theory that most substrates of Plk1 in metaphase are regulated by the following mechanism. The priming phosphorylation on a substrate by CDKs subsequently promotes the interaction between the substrate and Plk1. Plk1 consists of an N-terminal kinase domain and a C-terminal phosphopeptide binding domain named PBD (Polo Box Domain). In the absence of a bound substrate, the PBD forms an intramolecular interaction with the kinase domain, and inhibits the kinase activity. When a substrate is primed by another kinase like one of the CDKs, it binds to the PBD and releases the kinase domain for Plk1 function (Figure 4; Zhang et al. 2009).
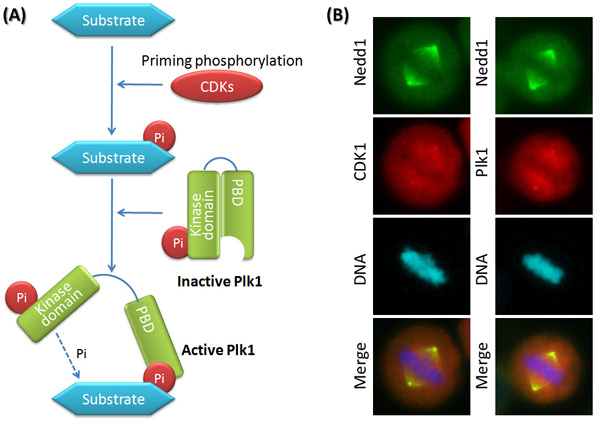
Figure 4: CDKs are the priming kinases for Plk1.
(A) shows a general model. In the absence of a bound substrate, Plk1 is inactive due to the intramolecular interactions. To bind Plk1, the substrate must be primed by another kinase like CDKs. Once phosphorylated, the substrate binds to Plk1 and releases the self-inhibited Plk1. Active Plk1 subsequently phosphorylates the substrate at more sites. (B) gives a specific example. During mitosis, CDK1, Plk1 and Nedd1 co-localize at the centrosome where CDK1 and Plk1 sequentially phosphorylate Nedd1.
© 2010 Nature Education All rights reserved.
Plk1 as a Priming Kinase to Aurora B
Scientists later discovered
a similar interaction between
Plk1 and Aurora B. While examining Xenopus egg extracts, they observed that substrates of Aurora B alone could inhibit the kinase activity. This observation raised the question of whether there were any cofactors facilitating the activity of Aurora B. They
discovered that the cofactors Plk1 and Haspin exert priming phosphorylation on
the substrates, and reverse the inhibitory effect on Aurora B. (Macurek et al.
2008). Could these findings suggest a common
mechanism of crosstalk among the three major mitotic kinases? Specifically,
does CDK1 prime substrates by phosphorylation, which then in turn activates
Plk1, which then further activates Aurora B by priming its substrates?
Collaboration between Kinases: One Kinase Phosphorylates Another
Aurora A Together with Bora Activates Plk1
Is there another pattern of collaboration among mitotic kinases? Indeed, scientists have exposed yet another point of interaction. Besides sequential phosphorylation on single substrate, protein kinases also phosphorylate and activate each other. During the cell cycle, the activation of Plk1 relies on the phosphorylation of a threonine residue within the activation segment. Scientists recently discovered that Aurora A is responsible for activating Plk1 at the G2/M transition by phosphorylating Plk1 at the same threonine residue. Plk activation by Aurora A requires another cofactor Bora, which dramatically enhances the ability of Aurora A to phosphorylate Plk1. Interestingly, this effect of Bora is specific to Plk1, rather than all Aurora A substrates, which hints at the possibility that Bora might be working directly on Plk1. Most likely, Bora induces the conformational change of Plk1 and makes it more accessible for Aurora A, but researchers have not yet proved this. Consistent with the model above, Bora needs to be primed by phosphorylation by CDK1 before it can bind to Plk1 (Figure 5; Seki et al. 2008, Taylor et al. 2008).
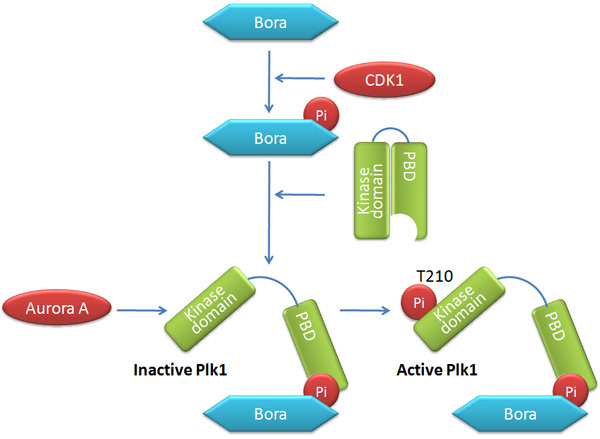
Figure 5: Aurora A together with Bora activates Plk1
During the cell cycle, the activation of Plk1 relies on the phosphorylation at T210 within the activation segment, which is performed by another kinase Aurora A. This process requires a cofactor Bora, which may induce Plk1 into a conformation that is easier for Aurora A to approach. The way Bora binds Plk1 fits the classical model as Bora needs to be primed itself by CDK1 phosphorylation.
© 2010 Nature Education All rights reserved.
Summary
Protein phosphorylation is a common way to regulate signaling pathways in the cell cycle. Kinases catalyze phosphoryl transfer from ATP to substrates and change downstream protein-protein interaction in such way that a signaling pathway is either switched on or shut off. Scientists have established the central roles of CDKs, Plks, and Aurora kinases in cell cycle regulation. Whereas CDKs provide the driving force for cell cycle progression, Plks and Aurora kinases function in a spatiotemporal manner to ensure correct cell division. These major mitotic kinases have attracted great attention in the cell cycle field, and their signaling pathways are each active subjects of research. Importantly, the emerging evidence of collaboration among these proteins uncovers a novel view of their complex regulation network both in time and space, and advances our understanding of cell cycle control.
References and Recommended Reading
Archambault, V. & Glover, D. M. Nature Reviews Molecular Cell Biology 10, 265–275 (2009) doi:10.1038/nrm2653.
Barr, F. A., Silljé, H. H. W. et al. Nature Reviews Molecular Cell Biology 5, 429–441 (2004) doi:10.1038/nrm1401.
Carmena, M. & Earnshaw, W. C. The cellular geography of Aurora. Nature Reviews Molecular Cell Biology 4, 842–854 (2003) doi:10.1038/nrm1245.
Fu, J., Bian, M., et al. Roles of Aurora kinases in mitosis and tumorigenesis. Molecular Cancer Research 5, 1–10 (2007) doi:10.1158/1541-7786.MCR-06-0208.
Hochegger, H., Takeda, S., et al. Cyclin-dependent kinases and cell-cycle transitions: does one fit all? Nature Reviews Molecular Cell Biology 9, 910–916 (2008) doi:10.1038/nrm2510.
Jeffrey, P. D., Russo, A. A., et al. Mechanism of CDK activation revealed by the structure of a
cyclinA-CDK2 complex. Nature 376, 313–320 (1995) doi:10.1038/376313a0.
Nigg, E. A. Mitotic kinases as regulators of cell division and its checkpoints. Nature Reviews Molecular Cell Biology 2, 21–32 (2001) doi:10.1038/35048096.
Ubersax, F. A & Ferrell Jr., J. E. Mechanisms of specificity in protein phosphorylation Nature Reviews Molecular Cell Biology 8, 530–541 (2007) doi:10.1038/nrm2203.










