


















« Prev Next »

Animals have to endure many stressors in their natural environments. For example, they experience food shortages, dwell in areas where predator or parasite densities are high, engage in conflicts with neighbors or group members, and face fluctuations in food and water availability and temperature. Although individuals can often predict when stressors will occur (e.g., more stressors generally in winter than summer), predictions about the intensity or duration of individual stressors are generally not possible (e.g., a failed predation attempt). Consequently, most species have evolved both constitutive and inducible mechanisms for coping with stressors (McEwen & Wingfield 2003).
Glucocorticoids: Vertebrate Stress Hormones
In vertebrates, an important mechanism for coping with stressors begins with adrenally-derived glucocorticoid hormones: corticosterone in amphibians, reptiles, and birds, or cortisol in fish and many mammals. These molecules drive gluconeogenesis, suppress reproductive processes, alter movement and feeding rates, impact immune functions, and generally help an individual enter a “state of emergency” when an environmental stressor induces their release (Wingfield et al. 1998). As an example, consider a wild, high-latitude bird getting ready to breed. For that individual, several factors may incite the release of corticosterone: an unexpected snowstorm, a conflict with a neighbor, a chase by a predator, or even an infection by a parasite. Growing evidence indicates that glucocorticoids are released in response to infection partly to support the use and mobilization of immune defenses.Although it has been known for some time that there are causal connections between stressors, glucocorticoids, and immune function, there is a dearth of data and theories addressing why they are connected in the first place (Martin 2009). Two non-exclusive hypotheses have been proposed. One proposes that stress hormones tend to suppress immune function, so that an individual’s resources are preferentially allocated to processes more crucial for surviving the stressor. The other proposes that attenuation of immune activities in response to stressors keeps the body from harming itself. During stress responses, many host-derived, novel molecules (e.g., antigens) are revealed to the immune system, and stress hormones prevent immune cells from mounting misdirected attacks (Råberg et al. 1998). Without stress hormones, the body would over-respond to benign substances and hence cause more damage than protection. To date, neither of these hypotheses has been tested directly.
The Importance of Understanding Stress-Immune Interactions
The study of interactions between stress hormones and immune functions is still a young field, yet research is critical for enhancing our knowledge of disease-coping mechanisms in wild animals, especially in light of escalating anthropogenic changes to the biosphere. Indeed, climate change, non-native species introductions, pollutant exposure, and habitat alterations — especially urbanization — all have the potential to alter stress responses and cause disease (Martin et al. 2010). Pollutants especially are implicated; their effects might be indirect via changes in host behavior, or direct via negative impacts on glucocorticoid regulation and subsequently immune cells and tissues. For example, exposure to atrazine, the world’s most common herbicide, at doses below standards set by the United States Environmental Protection Agency, can negatively impact immune functions (Rohr et al. 2008). Similar effects have recently been observed for chlorothalonil too, the most common fungicide used in the US (McMahon et al. in review).Anthropogenic stressors can impact vertebrates in less direct ways too though. Many urban bird populations regulate stress hormones differently than do their rural or suburban cousins (Bonier et al. 2007, Partecke et al. 2007, Fockidis et al. 2008), and the same urban populations can also experience greater parasite prevalence than rural and suburban populations (Bradley & Altizer 2007). Whether variation in stress hormones, immune functions, vector or parasite densities drive the above patterns is not clear, but mounting evidence suggests that combinations of factors are most likely responsible (Bradley & Altizer 2007, Natoli et al. 2005).
As with urbanization, few data exist on the impacts of introduced species on stress hormone-associated disease in native species. Only one study has reported that the presence of an introduced species impacted glucocorticoid regulation in a native species (Berger et al. 2007). In this study, neither immunological nor disease consequences of stress hormone variation was considered, but the possibility exists that introduced species may have negative impacts on stress hormones in native species, indirectly facilitating parasite spillover or even spillback (Kelly et al. 2009). Likewise, for climate change there are no reliable examples that indicate that climate change compromises immune functions via glucocorticoid-mediated pathways.
Stress Hormones Regulate Immune Functions in Natural Populations
To date, most of what we know about stress-immune interactions research has come from domesticated species. However, since domesticated species have been either actively or passively selected for docility, amenability, and human handling, their immune and stress hormone response systems may be unrepresentative of their free-living counterparts (Trut et al. 2009). Two ways in which wild animal stress-immune interactions will likely differ from domesticated animals, but be illuminating for how we understand stress-immune interactions generally, include balancing of parasite resistance versus tolerance and immune redistribution.
Balancing parasite resistance and tolerance
Physicians have long known of the immunosuppressive effects of glucocorticoids, and have used synthetic glucocorticoids to suppress immune functions and assuage the symptoms of, or complications from, inflammatory disorders (e.g., asthma, arthritis, etc). Whereas all immune functions are eventually suppressed by glucocorticoids, adaptive immune defenses are most sensitive. After only a few days of glucocorticoid elevation, T and B cells can be induced to self-destruct (i.e., apoptose) (Sternberg 2006). Innate immune cells, and especially macrophages, can be quite resistant to glucocorticoids however, and in some cases, their resistance can increase in response to chronic glucocortoid or stressor exposure (Bailey et al. 2004).
By and large, most research on glucocorticoid effects on immune system function has focused on suppressive effects, not surprisingly given many of the above effects. However, glucocorticoids have complex effects on immune functions, an observation consistent with an emerging theme in ecology that hosts can either resist or tolerate parasites (Råberg et al. 2009). Resistance involves mechanisms that limit parasite burden, whereas tolerance involves mechanisms that limit the impacts of a given burden on reproductive fitness or performance. It is important to distinguish between these mechanisms because they impact both the persistence and emergence of disease as well as the evolution of virulence differently (Restif & Koella 2004). For these reasons, it will be important to determine whether and how stress hormones impact tolerance. Several indirect lines of evidence indicate that glucocorticoids mediate tolerance (e.g., release of the cytokine, interleukin-10) (Sternberg 2006). More importantly, once resistance barriers are breached, tolerance mechanisms must be activated. As glucocorticoids can reduce resistance (i.e., immunosuppression), they may also foster tolerance (Figure 1). To test this hypothesis, one could assess whether for a given parasite burden individuals with high glucocorticoids also perform better than individuals with low glucocorticoids. A more direct option would be to implant or block glucocorticoids, expecting that glucocorticoids would benefit performance at a given parasite burden if glucocorticoids promote tolerance. Although some of these data are likely already available, poor study design (i.e., lack of consistency in parasite exposure and lack of variation in parasite burden) make the observations difficult to interpret.
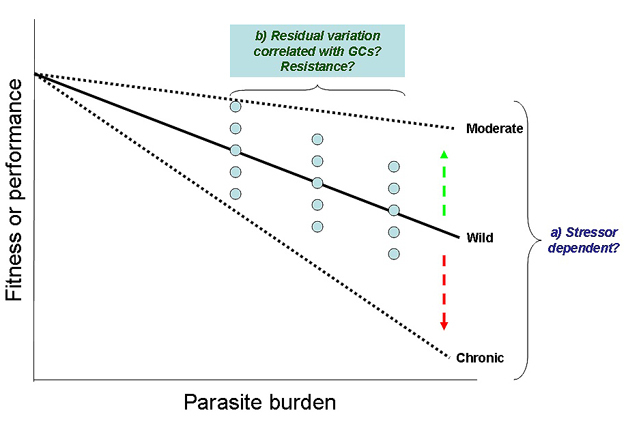
By and large, most research on glucocorticoid effects on immune system function has focused on suppressive effects, not surprisingly given many of the above effects. However, glucocorticoids have complex effects on immune functions, an observation consistent with an emerging theme in ecology that hosts can either resist or tolerate parasites (Råberg et al. 2009). Resistance involves mechanisms that limit parasite burden, whereas tolerance involves mechanisms that limit the impacts of a given burden on reproductive fitness or performance. It is important to distinguish between these mechanisms because they impact both the persistence and emergence of disease as well as the evolution of virulence differently (Restif & Koella 2004). For these reasons, it will be important to determine whether and how stress hormones impact tolerance. Several indirect lines of evidence indicate that glucocorticoids mediate tolerance (e.g., release of the cytokine, interleukin-10) (Sternberg 2006). More importantly, once resistance barriers are breached, tolerance mechanisms must be activated. As glucocorticoids can reduce resistance (i.e., immunosuppression), they may also foster tolerance (Figure 1). To test this hypothesis, one could assess whether for a given parasite burden individuals with high glucocorticoids also perform better than individuals with low glucocorticoids. A more direct option would be to implant or block glucocorticoids, expecting that glucocorticoids would benefit performance at a given parasite burden if glucocorticoids promote tolerance. Although some of these data are likely already available, poor study design (i.e., lack of consistency in parasite exposure and lack of variation in parasite burden) make the observations difficult to interpret.
Figure 1: Predicted effects of glucocorticoids on tolerance.
Black text indicates duration, arrows depict directional change in tolerance, and blue circles indicate individual (residual) variation for tolerance.
© 2012 Nature Education All rights reserved. 
Immune redistribution
A dominant focus in stress-coping research has been the effects of acute versus chronic stressors on performance and fitness, including immune functions. As above, chronic stressors tend to suppress immune functions, although immune cells and tissues vary in their sensitivity to stressors and stress hormones. Interestingly though, acute stressors often enhance immune functions as a result of glucocorticoid-immune interactions. These effects include increased memory of antigens (among other effects), but the best-studied effects are enhancements of skin inflammatory responses and immune cell influx to wounds. These latter effects are especially relevant for wild animals, because wounds are a common consequence of failed predation attempts and territorial conflicts, two of the most common stressors wild animals experience. These observations led to the hypothesis that acute stressor induced immune enhancement might include immune redistribution (Dhabhar & McEwen 1997). In support of this hypothesis, brief restraint of domesticated rodents prior to the induction of a skin inflammatory response enhanced both the physical size of the immune response and the number of immune cells entering the skin (Dhabhar 2009).
Analogous experiments have not been successfully performed in wild animals, partly because it is difficult to conduct such studies on animals that perceive handling by humans, and housing in captivity, as predation events. For example, while attempting to determine whether acute stressors enhanced immune function in the skin of wild house sparrows (Kuhlman & Martin 2007), captivity itself was found to impact immune responses. After a couple of days in captivity, the type and abundance of immune cells infiltrating the skin began to change; after four weeks in captivity, the makeup of the immune cells entering the skin was altogether different than that of a wild-caught bird. Subsequent studies have indicated that glucocorticoid regulation breaks down over captivity in house sparrows: baseline levels remain very high, release in response to stressors no longer occurs, and hormone receptor expression in the brain changes after a month in what are perceived to be benign, controlled lab conditions (Martin et al. in review). Thus, to determine whether immune redistribution is important in wild animals, future experiments will require creative study designs to overcome the confounding effects of captivity.
Prospects for Future Studies
Glucocorticoid and other stress hormones regulate immune defenses and hence impact how hosts control parasites and disease. We still know too little, however, about when and how these processes occur in wild animals, and what consequences their dysregulation has for the health of natural communities. Furthermore, there are still no studies that definitively link a stressor, stress hormones, and disease in free-living species. In other words, a stressor alters stress hormones, stress hormone alterations impact immune functions, and the altered immune functions change the risk of mortality or morbidity to infections. In an era when so much effort (and money) tends to favor technical (i.e., molecular and genomic) research approaches, we feel that study of whole organisms in their natural contexts and with evolutionary principles in mind will be insightful and beneficial (Calisi & Bentley 2009).References and Recommended Reading
Bailey, M. T. et al. Physical defeat reduces the sensitivity of murine splenocytes to the suppressive effects of corticosterone. Brain Behavior and Immunity 18, 416-424 (2004). doi:10.1016/j.bbi.2003.09.012
Berger, S. et al. Behavioral and physiological adjustments to new predators in an endemic island species, the Galapagos marine iguana. Hormones and Behavior 52, 653-663 (2007). doi:10.1016/j.yhbeh.2007.08.004
Bonier, F. et al. Sex-specific consequences of life in the city. Behavioral Ecology 18, 121-129 (2007). doi:10.1093/beheco/ar1050
Bradley, C. A., & Altizer, S. Urbanization and the ecology of wildlife diseases. Trends in Ecology & Evolution 22, 95-102 (2007). doi:10.1016/j.tree.2006.11.001
Calisi, R. M., & Bentley, G. E. Lab and field experiments: Are they the same animal? Hormones and Behavior 56, 1-10 (2009). doi:10.1016/j.yhbeh.2009.02.010
Dhabhar, F. S., & McEwen, B. S. Acute stress enhances while chronic stress suppresses cell-mediated immunity in vivo: A potential role for leukocyte trafficking. Brain Behavior and Immunity 11, 286-306 (1997).
Dhabhar, F. S. Enhancing versus Suppressive Effects of Stress on Immune Function: Implications for Immunoprotection and Immunopathology. Neuroimmunomodulation 16, 300-317 (2009). doi:10.1159/000216188
Fokidis, H. B., Greiner, E. C., & Deviche, P. Interspecific variation in avian blood parasites and haematology associated with urbanization in a desert habitat. Journal of Avian Biology 39, 300-310 (2008). doi:10.1111/j.2008.0908-8857.04248.x
Kelly, D. W. et al. Parasite spillback: A neglected concept in invasion ecology? Ecology 90, 2047-2056 (2009). doi:10.1890/08-1085.1
Kuhlman, J. R., & Martin, L. B. Captivity affects immune redistribution to skin in a wild bird. Functional Ecology 24, 830-837 (2010). doi:10.1111/j.1365-2435.2010.01710.x
Martin, L. B. Stress and immunity in wild vertebrates: Timing is everything. General and Comparative Endocrinology 163, 70-76 (2009). doi:10.1016/j.ygcen.2009.03.008
Martin, L. B. et al. The effects of anthropogenic global changes on immune functions and disease resistance. Annals of the New York Academy of Sciences (2010).
Martin, L. B. et al. Does immune suppression occur to free resources for other physiological processes? General and Comparative Endocrinology (in review).
McEwen, B. S. & Wingfield, J. C. The concept of allostasis in biology and biomedicine. Hormones and Behavior 43, 2-15 (2003).
McMahon, T. et al. Chlotothalonil: An immunomodeulatory and deadly fungicide to amphibians. (in review).
Natoli, E. et al. Bold attitude makes male urban feral domestic cats more vulnerable to Feline Immunodeficiency Virus. Neuroscience and Biobehavioral Reviews 29, 151-157 (2005). doi:10.1016/j.neubiorev.2004.06.011
Partecke, J., Schwabl, I., & Gwinner, E. Stress and the city: Urbanization and its effects on the stress physiology in European Blackbirds. Ecology 87, 1945-1952 (2006).
Råberg, L., Graham, A. L., & Read, A. F. Decomposing health: Tolerance and resistance to parasites in animals. Philosophical Transactions of the Royal Society B: Biological Sciences 364, 37-49 (2009). doi:10.1098/rstb.2008.0184
Råberg, L. et al. On the adaptive significance of stress-induced immunosuppression. Proceedings of the Royal Society B: Biological Sciences 265, 1637-1641 (1998).
Restif, O., & Koella, J. C. Concurrent evolution of resistance and tolerance to pathogens. The American Naturalist 164, E90-102 (2004).
Rohr, J. R. et al. Evaluating the links between climate, disease spread, and amphibian declines. Proceedings of the National Academy of Sciences of the United States of America 105, 17436-17441 (2008). doi:10.1073/pnas.0806368105
Sternberg, E. M. Neural regulation of innate immunity: A coordinated nonspecific host response to pathogens. Nature Reviews Immunology 6, 318-328 (2006), doi:10.1038/nri1810
Trut, L., Oskina, I., & Kharlamova, A. Animal evolution during domestication: The domesticated fox as a model. Bioessays 31, 349-360 (2009). doi:10.1002/bies.200800070
Wingfield, J. C. et al. Ecological bases of hormone-behavior interactions: The "emergency life history stage". American Zoologist 38, 191-206 (1998).










