



















« Prev Next »
Introduction
The upper altitudinal limit of trees is a global phenomenon. This limit, often called the treeline, bounds the conditions that allow trees to grow, survive, and reproduce. Beyond it, the maintenance of viable populations and expansion of tree distribution are constrained. Despite widespread recognition of the existence of treelines and scientific study since the early 1900s (e.g., Brockmann-Jerosch 1919, Däniker 1923) and the likely influence of temperature, there is no overarching explanation for what causes treelines around the world. This stems mainly from:
- the many environmental conditions that change when moving from low-elevation tree-covered areas to high-elevation treeless ecosystems, and
- the multiple biological mechanisms that could explain the boundary based on these different environmental conditions.
Trees, Treelines, and Global Patterns
Before one can explain the biological mechanisms behind treeline limits, it is valuable to consider what is a "tree" and to think about whether a line is an accurate concept for the limit of tree existence. Trees are upright, woody plants with a single, dominant above-ground stem. Many scientists restrict this definition to plants that are somewhere between 2 and 5 m in height (Holtmeier 2009). Trees by this definition also have a crown that is closely coupled to the surrounding atmosphere (Körner 1998), as opposed to the shrubs and herbaceous vegetation that exist beyond the treeline.
The next problem is identifying where the treeline occurs, since the natural boundary is rarely a defined line and instead forms a fuzzy transitional zone (Figure 1). Geographic analysis has advanced scientists' ability to detect treeline position (Allen & Walsh 1996) and analyze the factors that influence it in a spatial context (Malanson et al. 2007), but multiple definitions are still used depending on the focus of each study. The boundary can be defined as the end of closed forest (the timberline), the upper limit of all tree existence (the tree form limit, Smith et al. 2003) or an altitude between these two points where tree density decreases beyond a certain threshold (Körner & Paulsen 2004; Figure 2). Consequently, there is no single "treeline" (Malanson et al. 2007) and it is important for researchers to state their assumptions clearly.

Figure 1: Satellite view of a treeline in Colorado, USA, demonstrating the complexity of the treeline boundary
© 2010 Nature Education All rights reserved. 
Using a rough definition of the treeline as "the elevation where tree density declines or drops to zero," it is possible to identify variation in treeline position around the globe. A strong pattern exists between treeline altitude (in meters above sea level) and latitude globally (Figure 3). In a similar analysis with another set of data, Körner (1998) found a nearly identical pattern and identified three important trends:
- There is no uniform relationship between treeline position and latitude. Treelines decline steeply between temperate and boreal latitudes but are relatively constant between ~32°N and 20°S, and the pattern is not symmetrical around the equator.
- Variation within latitudinal bands suggest that other climatic factors not related to latitude (like microclimate variation) influence the altitude of treelines.
- Treeline positions follow a similar trend to the permanent snow line (not shown), suggesting a common physical driver (e.g., temperature).
An additional important trend is the observation that treeline on oceanic islands is substantially lower than the continental treeline (Figure 3).
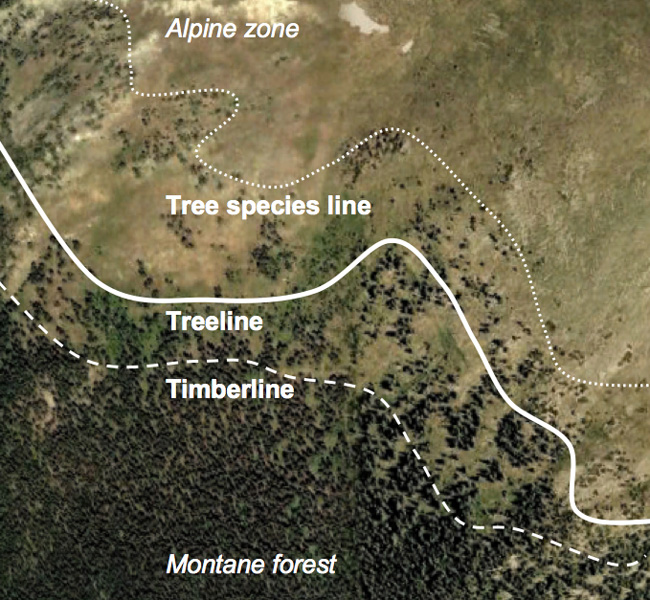
Figure 2: Conceptual representation of the timberline (the limit of closed tall forest), the treeline (the limit of groups of trees over 3 m high), and the tree species line (the limit of all individual trees)
© 2010 Nature Education All rights reserved.
Explaining Treeline Patterns
Problems with Multiple Conditions
To explain where treelines occur on a global scale, it is first valuable to identify those environmental variables that are most similar across treeline locations. These variables could influence the biological mechanisms behind treeline formation and maintenance. The main problem with identifying climatic factors that influence treelines is that numerous variables co-vary with both latitude and altitude.
Latitudinally, equatorial zones have higher mean annual temperatures, lower seasonal variation in temperature, longer growth periods, larger diurnal variation in temperature, and higher incident solar radiation (Körner 1998). Subtropical zones also have relatively little cloudiness. All these variables could theoretically be used to explain latitudinal variation in treeline position (Figure 3). Additionally, across the treeline boundary from low to high elevation in temperate latitudes, temperature and growing season length often decrease while precipitation, cloudiness, and wind speeds tend to increase.
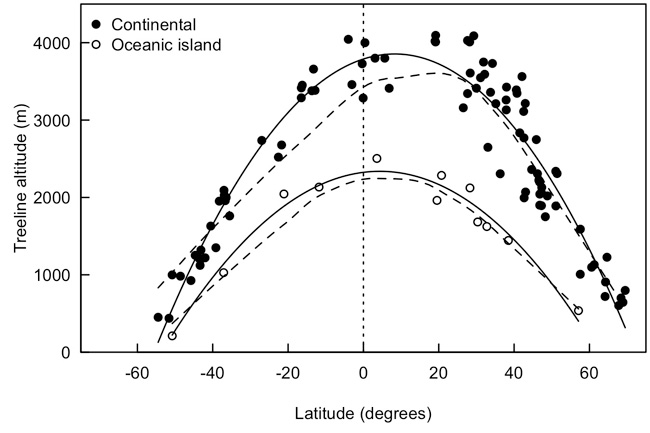
Figure 3: Treeline position around the globe by latitude for continental (filled points) and oceanic island (open points) sites (N = 105)
Treeline was identified with satellite imagery as a point where tree density decreased sharply. Data points were fit with quadratic (solid lines) and loess (dashed lines) curves. Data were collected by Aaron Berdanier, Anna Ellis, Katherine Geist, Melissa Kattke, Casey Perkins, Alyssa Schultz, and Jared Wynne.
© 2010 Nature Education All rights reserved.
Many attempts have been made to define global and regional climate indices for treeline boundaries, such as mean or summed air/soil temperatures. For example, across sites around the globe, mean air temperature during the growing season at the treeline appears to vary roughly between 5.5 and 7.5°C (Körner 1998, Körner & Paulsen 2004). Although multiple outliers make it an inconclusive pattern (Holtmeier 2009), this relatively tight range is surprising given large differences in growing season length and seasonal and diurnal temperature fluctuations among treelines globally. Consistency in growing season temperature at the treeline supports the possibility of a thermal boundary preventing the survival, growth, and/or reproduction of trees at high altitudes. Globally, this could explain lowered treeline position at high latitudes as well as the tropical plateau (Figure 3), given reduced solar radiation and less radiative warming that is associated with tropical cloudiness (Körner 1998). However, while mean temperatures are an indicator of the thermal conditions at the treeline, they cannot solely explain why a boundary exists.
Possible Explanations
While treelines are global in nature and can be compared to climate, the mechanisms behind their existence act at smaller scales (Holtmeier & Broll 2005). Individuals respond to their local environment and form populations that are used to define the treeline boundary (Malanson et al. 2007). Scientists have identified a handful of possible mechanisms that could explain large scale patterns in treeline position, given global variation in treeline altitude and changes in important climatic variables. Körner (1998) and Smith et al. (2003) summarize these explanations with five main hypotheses. These hypotheses are distinct from one another but are not necessarily mutually exclusive. Additionally, while each factor may determine the high-elevation limit of tree existence for individual populations, some may be more important than others for explaining the general, global pattern of treeline altitudes (Körner 1998).
- Stress — physiological tissue damage due to low temperature or dessication. During the coldest part of the year, frost can damage cells by freezing or photoinhibition. This is pronounced for trees due to their height, which results in a tighter coupling to the atmosphere than other short, alpine plants. Additionally, frozen soil can limit water availability, causing dessication of plant tissue, contributing to reduced tree growth at high elevations. However, this factor does not appear to be widespread at all sites, especially for established trees (Körner 1998), although it could play a role in limiting growth in young trees (Smith et al. 2003).
- Disturbance — mechanical damage due to wind abrasion, herbivory, snow loading, or fungi infection. In high-elevation ecosystems, snow and ice loading can break or remove biomass on trees. Additional damage to needles occurs with high wind that often exists at and above the treeline. Wind damage may be more pronounced for trees than for the short, alpine vegetation that often exists far beyond the treeline. Additionally, when snow covers much of the surrounding alpine vegetation, the vegetative growth and meristems of developing trees are particularly susceptible to herbivory. Snow mold and other fungal pathogens can also damage tissue. As with physical stress, disturbance is likely important at a local/regional level but is not widespread globally, as many tropical treelines form without the influence of snow and the presence of herbivores/pathogens is inconsistent.
- Reproduction — reduced seedling and sapling establishment due to decreased pollination, seed development, seed dispersal, germination, or seedling establishment. While the treeline is a clear boundary for the existence of trees, small shrubs and other low-lying forbs and grasses often exist above the treeline (in the "alpine" vegetation zone). The upper edge of the treeline boundary could be created by an inability of trees to produce seeds, disperse, colonize, and/or survive at high-elevations. While there is no evidence that trees at high elevations are widely limited by production and dispersal of seeds (Körner 1998), low average temperatures can limit seed maturation, and climatic limitation of germination and seedling establishment could diminish the ability of trees to expand their range to higher altitudes (see below).
- Carbon balance — photosynthetic carbon gain minus respiratory demands is not enough to maintain minimum growth. The primary component of growth is net carbon gain, through photosynthesis minus respiration. High-elevation plants appear to be acclimated to cold temperatures such that photosynthesis per leaf area is relatively insensitive to low temperature over the average range of growing season values (Körner 1998). The main restriction of carbon gain in these ecosystems is from the length of the growing period. If temperatures are too low and the growth period is too short for photosynthesis in the growing season to exceed the annual respiratory demand, a tree will not be able to survive.
- Growth limitation — reduced development of new plant tissues due to low temperatures. The second component of growth is the use of photosynthetic sugars and amino acids to produce new cells and grow new plant tissues. Even if carbon supply from photosynthesis is sufficient, low temperature could limit growth and development. This limitation may be more pronounced for trees than for other plant forms because of differences in the vegetative microclimate (Körner 1998). For example, both above-ground and below-ground tree plant tissues may experience colder conditions than low-lying alpine vegetation (see below).
The depressed treeline position on oceanic islands (open points, Figure 3) provides a unique opportunity to consider why these sites are different from continental sites. Four hypotheses have been proposed to explain this pattern:
- the immaturity of volcanic soils found on most of the islands,
- the absence of well-adapted high-altitude tree species that are found on continents,
- aridity and drought above the treeline on trade wind exposed islands, and
- altered vertical temperature gradients on mountains of different sizes (called Massenerhebungseffekt, or the mountain mass effect, Leuschner 1996).
These explanations emphasize the importance of site-specific variables for determining treeline position, particularly site history and local microclimate (Malanson et al. 2007, Holtmeier 2009).
Current Main Theories for Treeline Position
The main explanations for global treeline position focus on a distinction between establishment (Smith et al. 2003) and growth (Shi et al. 2008) limitations, depending on where one defines the treeline. Other explanations (e.g., disturbance, drought, and site history) are important in many places throughout the world but are less consistent globally. Both establishment and growth hypotheses are based on a strong relationship between vegetation and microclimate, emphasizing a connection to temperature, while one focuses on range expansion at the upper tree limit and the other on survival.
Limited Seedling Establishment
Reduced seedling establishment is a critical limiting factor on the colonization of trees at high altitudes. The inability of trees to become established defines the upper boundary of tree existence (the tree form limit). Both tree germination capacity (Viereck 1979, Holtmeier 2009) and establishment (Germino et al. 2002) sharply decline at higher altitudes due to both large-scale climate and local microclimate variation. While seedlings within a patch of trees near the treeline are protected by their neighbors, at the tree form limit seedling establishment may depend on physical objects (e.g., rocks) and neighboring plants that alter microclimate and create conditions beneficial for plant growth (Germino et al. 2002, Smith et al. 2003). For example, in the winter, seedlings may be protected from ice crystal abrasion, dessication, and diurnal temperature fluctuations whereas in the summer seedlings benefit from decreased sky and wind exposure (increasing daytime temperature).
Growth Limitation
Once trees are established, survival depends on maintaining growth, which is limited by cold temperatures at high altitudes (Körner 1998). Trees at the treeline may experience colder temperatures than surrounding vegetation for two reasons:
- Shoot meristems in tall tree canopies do not experience warming from the canopy during the day or from the soil during the night. Instead, growth of these meristems is more tightly coupled to the surrounding atmosphere, and they experience convective cooling.
- Trees with large canopies prevent solar radiation from reaching and warming the soil around the base of the tree, reducing root zone temperatures and, potentially, root physiology.The growth limitation hypothesis, in contrast to the carbon-balance hypothesis, is supported by findings that treeline trees tend to have higher amounts of non-structural carbohydrates and lipids, suggesting that these organisms are limited by growth more than carbon assimilation (Shi et al. 2008, but see Hättenschwiler et al. 2002, who found an increase in treeline photosynthesis and growth with increased atmospheric CO2).
Resolving Hypotheses
While it is clear that very few variables are truly global in scope, and some variables are more important than others, an integrated theory for the global pattern of treelines is not yet resolved. Further work will require testing hypotheses at multiple sites to identify which mechanisms predominate. This could involve experimental, observational, and simulation modeling studies. For example, it is possible to experimentally
- alter carbon availability (through atmospheric CO2 enrichment), to explore the carbon balance hypothesis,
- cause physical damage, to explore disturbance hypotheses, and
- alter the microclimate, to explore establishment and growth hypotheses.
These experimental approaches have been used successfully in some locations and could be replicated at many other sites to increase knowledge. Where experimental manipulations are impractical, simulation modeling studies and observational analyses of microclimatic variables could greatly enhance our understanding of large-scale patterns. Regionally, comparisons of upper and lower tree limits could explore why some treeline ecotones are broader than others. Finally, modeling tree survival, growth, and reproduction, combined with observational data from numerous sites, could offer a way to explore the relative importance of underlying causes at a global scale.
Just as there are many "treelines," there are many factors that can influence where these treelines occur. Tree metabolism, growth, spread, and survival are all local phenomena. Predicting treeline position accurately requires consideration of all of the variables mentioned. However, global patterns exist (Figure 3) and are clearly constrained by cold temperatures (Holtmeier 2009). It is likely that both establishment and growth limitation could provide the best explanations for why trees decline sharply at high elevations. Further research will continue to provide insight into which mechanisms are most important in determining global treeline position.
References and Recommended Reading
Allen, T. R. & Walsh, S. J. Spatial and compositional pattern of alpine treeline, Glacier National Park, Montana. Photogrammetric Engineering & Remote Sensing 62, 1261–1268 (1996).
Brockmann-Jerosch, H. Baumgrenze und Klimacharakter in Pflanzengeographische Kommission der Schweiz. Naturforschenden Gesellschaft, Beiträge zur Geobotanischen Landesaufnahme, vol. 6. Rascher, Zürich, 1919.
Däniker, A. Biologische studien über baum- und waldgrenze, insbesondere über die klimatischen ursachen und deren zusammenhänge. Vierteljahresschrift Naturforschende Gesellschaft Zürich 68, 1–102 (1923).
Germino, M. J., Smith, W. K., & Resor, A. C. Conifer seedling distribution and survival in an alpine treeline ecotone. Plant Ecology 162, 157–168 (2002).
Hättenschwiler, S. et al. Atmospheric CO2 enrichment of alpine treeline conifers. New Phytologist 156, 363–375 (2002).
Holtmeier, F. Mountain Timberlines: Ecology, Patchiness, and Dynamics. New York, NY: Springer, 2009.
Holtmeier, F. & Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Global Ecology and Biogeography 14, 395–410 (2005).
Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 115, 445–459 (1998).
Körner, C. & Paulsen, J. A world-wide study of high altitude treeline temperatures. Journal of Biogeography 31, 713–732 (2004).
Leuschner, C. Timberline and alpine vegetation on the tropical and warm-temperate oceanic islands of the world: Elevation, structure and floristics. Vegetatio 123, 193–206 (1996).
Malanson, G. P. et al. Alpine treeline of Western North America: Linking organism-to-landscape dynamics. Physical Geography 28, 378–396 (2007).
Shi, P., Körner, C., & Hoch, G. A test of the growth-limitation theory for alpine tree line formation in evergreen and deciduous taxa of the eastern Himalayas. Functional Ecology 22, 213–220 (2008).
Smith, W. K. et al. Another perspective on altitudinal limits of alpine timberlines. Tree Physiology 23, 1101–1112 (2003).
Viereck, L. A. Characteristics of treeline plant communities in Alaska. Holarctic Ecology 2, 228–238 (1979).
Walter, H. Vegetation of the Earth in Relation to Climate and Eco-physiological Conditions. London, UK: English University Press, 1973.










