
Abstract
Lowland residents adapt to the reduced oxygen availability at high altitude through a process known as acclimatisation, but the molecular changes underpinning these functional alterations are not well understood. Using an integrated biochemical/whole-body physiology approach we here show that plasma biomarkers of NO production (nitrite, nitrate) and activity (cGMP) are elevated on acclimatisation to high altitude while S-nitrosothiols are initially consumed, suggesting multiple nitrogen oxides contribute to improve hypoxia tolerance by enhancing NO availability. Unexpectedly, oxygen cost of exercise and mechanical efficiency remain unchanged with ascent while microvascular blood flow correlates inversely with nitrite. Our results suggest that NO is an integral part of the human physiological response to hypoxia. These findings may be of relevance not only to healthy subjects exposed to high altitude but also to patients in whom oxygen availability is limited through disease affecting the heart, lung or vasculature and to the field of developmental biology.
Similar content being viewed by others

Introduction
Barometric pressure falls with increasing altitude whilst the percentage of oxygen within the atmosphere remains constant. The consequent reduction in inspired oxygen impairs physical performance and may even threaten survival.1 Graded and continued altitude exposure permits acclimatisation, a progressive tolerance to hypoxia traditionally ascribed to cardiopulmonary and haematological responses that support global convective oxygen delivery.2 However, such responses do not provide a full explanation of the acclimatisation process given that exercise tolerance remains limited even though arterial oxygen content returns to sea-level values.3,4 Furthermore, genetic selection pressure amongst high-altitude residents of the Tibetan Plateau (in situ for >20,000 years) favours lower rather than higher levels of oxygen-carrying haemoglobin.5,6 Finally, the marked inter-individual differences which exist in the ability to adapt to hypoxia are not explained by differences in arterial oxygen content.3 Changes in tissue oxygen handling, mediated in part through changes in the microcirculation and in mitochondrial oxygen use, are increasingly considered more important to the development of hypoxia tolerance than is the augmentation of its global convective delivery.7,8,9 If acclimatisation is in part dependent upon these mechanisms, how might this occur?
Nitric oxide (NO) is a ubiquitous signalling molecule produced through the metabolism of L-arginine by nitric oxide synthases (NOS). Its formation is crucial for the control of blood pressure, blood flow and other vital bodily functions.10 Amongst these, it is an important antioxidant,11 and a regulator of intermediary metabolism12 and cellular energy production by mitochondria.13 NO availability may be reduced at altitude since i) its enzymatic production depends on the availability of oxygen and ii) exposure to hypoxia results in a paradoxical increase in the production of reactive oxygen species (in particular when combined with physical activity)14 leading to NO inactivation. This is consistent with the notion that hypoxia leads to an increase in the expression of different NOS isoforms via upregulation of HIF-1α, a master transcriptional regulator of oxygen homeostasis. However, the situation is complex inasmuch as HIF-1α stability is controlled not only by the availability of oxygen but also that of NO.15 Moreover, recent studies suggest an alternative pathway of NO generation16 that involves sequential reduction of nitrate (NO3−) to nitrite (NO2−) and further to NO and which is inhibited by oxygen.17 Perhaps for these reasons, Tibetan highlanders exhibit elevated levels of circulating NO products (including plasma nitrate and nitrite) when compared to lowlanders. The combination of high NO production and low haemoglobin concentration in this population is associated with increased forearm blood flow.18 Intriguingly, lowlanders adapting to high altitude exhibit reduced blood flow in the microcirculation (arterioles, capillaries and venules less than 100 µm in diameter).19 Whether or not this phenomenon is modulated by endogenous NO or its metabolites is unknown. Recent reports of reduced oxygen cost of exercise and improved mitochondrial efficiency following dietary nitrate supplementation at sea-level20,21 support the notion that high nitrite/nitrate levels in Tibetans might be associated with beneficial metabolic adaptation to hypoxia. Thus, enhanced NO production may not be unique to highlanders but part of the integral human physiological response to hypoxia. We hypothesized that NO availability would be increased in lowland residents acclimatizing to altitude and that this would be associated with (i) reduced oxygen cost and improved efficiency of oxygen utilization during exercise and (ii) increased microcirculatory blood flow.9
Results
Basic physiological changes on ascent to Everest Base Camp
Incremental exposure of healthy lowlanders to profound hypoxia at high altitude was accompanied by a progressive increase in respiratory rate (particularly upon exercise), heart rate and blood pressure (Fig. 1a), the expected cardiopulmonary adjustments to the unavoidable reduction in oxygen availability. Unchanged plasma osmolality (Fig. 1b) indicates that individuals remained well hydrated. Furthermore, unchanged plasma protein and creatinine concentrations provide no evidence of protein catabolism or renal dysfunction. Differences in circulating biomarker levels, therefore, reflect true concentration changes rather than body fluid shifts. Average body weight changes remained below 5% at all altitudes. Unchanged plasma levels of the (pro)inflammatory cytokines IL-6 and TNF-α document the absence of acute inflammation (Fig. 1b).

Ascent Profile and Altitude-Related Effects on the Cardio-Pulmonary System as well as the Delivery and Consumption of Oxygen During the 2007 Caudwell Xtreme Everest Expedition.
(a) Arrows in top chart indicate location and timing of exercise testing and blood sampling. Breathing frequency and heart rate before and after a standardized 2-min step exercise protocol and resting blood pressure were recorded daily for all members of the Core Team (see Methods for details). (b) Changes in haematocrit, plasma osmolality and protein concentration were quantified together with circulating levels of interleukin-6 (IL-6) tumor necrosis factor alpha (TNF-α), creatinine and erythropoietin (EPO). (c) Variables affecting oxygen delivery included partial pressure of oxygen (black bars), arterial oxygen saturation at rest (dark grey bars), haemoglobin concentration (light grey bars) and arterial oxygen content (open bars). (d) Effects of hypoxia on exercise performance, expressed as rates of oxygen consumption at the lactate threshold (LaT) and at maximal work rate (VO2max). Results are means ± SEM; n = 24.
Oxygen saturation of arterial haemoglobin decreased with ascent due to the reduction in inspired oxygen partial pressure (Fig. 1c). Consequently, haemoglobin concentration increased under the influence of elevated erythropoietin such that oxygen content was maintained at sea-level values at all altitudes. Despite this, oxygen consumption at the lactate threshold and at VO2peak decreased with ascent, with peak oxygen consumption falling by 35% at 5,300m (Fig. 1d).
Ascent to high altitude is accompanied by increased oxidative stress
Altitude acclimatisation was accompanied by elevation in the levels of three independent indices of oxidative stress: increased concentrations of (i) 8-iso-prostaglandin F2α (F2-isoprostane, a lipid oxidation product and potential disease mediator frequently used in translational studies) and (ii) 4-hydroxy-2-nonenal (HNE, a lipid peroxidation product known to form stable adducts with cysteine and histidine residues in proteins and to play a role in cell signalling) and (iii) a decrease in the ratio of reduced/oxidized glutathione, the most abundant low-molecular weight antioxidant (Fig. 2a). While the biological sources of the lipid oxidation products and the main site(s) of glutathione oxidation remain unclear and changes in these plasma indices peak at different altitudes (presumably due to variations in chemical reactivity and pathways of formation), a consistent elevation of all three biomarkers compared to sea-level values indicates sustained oxidative stress. In the absence of a compensatory increase in NO production, such changes would be expected to result in enhanced inactivation of NO by superoxide22 and perhaps other reactive oxygen species, causing reduced NO availability and consequently impaired local NO-cGMP signalling.

Ascent to Everest Base Camp is Associated with Increased Oxidative Stress as well as Enhanced Formation and Availability of Nitric Oxide.
(a) Effects of hypoxia on oxidative stress biomarkers are expressed as differences in plasma concentration of 8-iso-prostaglandin-F2α (8-iso-PGF; F2-isoprostanes), 4-hydroxy-2-nonenal (HNE) and the ratio of reduced to oxidized glutathione (GSH/GSSG) at the indicated altitudes from normoxic concentrations at sea level (75 m, London, UK); (b) Circulating concentrations of the nitric oxide related metabolites, nitrite (NO2−) and nitrate (NO3−) as well as the second messenger cyclic guanosine 3’, 5’-monophosphate (cGMP) are elevated in hypoxia while plasma S-nitrosothiols (RSNOs) experienced a dramatic drop compared to sea level conditions on arrival at Kathmandu (1,300 m) before progressively increasing. In contrast, pro atrial natriuretic peptide (proANP), brain natriuretic peptide (BNP) and C-type natriuretic peptide (CNP) variations are minimal. Plasma concentrations at each altitude correspond to time of exercise testing; means ± SEM (n = 24). (c) Correlations between NO metabolites and variables of oxygen delivery and consumption at sea level (all variables log-transformed).
Production and availability of nitric oxide are enhanced during ascent to high altitude
NO formation and availability were assessed by quantification of the oxidative decomposition products, nitrite and nitrate, the second messenger cyclic 3’,5’-guanosine monophosphate (cGMP) and protein nitrosation products in plasma. Plasma nitrate and nitrite levels were increased at all altitudes above 1,300m, indicative of elevated NO production, with maximal levels at 3,500m, following the greatest rate of altitude gain (Fig. 2b). Elevation of cGMP, in the context of minimal fluctuations in plasma natriuretic peptide (ANP, BNP, CNP) levels (an alternative pathway through which cGMP could be enhanced), indicates increased NO availability. Cyclic GMP was elevated at all altitudes above sea level, also peaking at 3,500m. In contrast, protein nitrosation products, approximately half of which corresponded to S-nitrosothiols (RSNOs), dropped by >50% upon arrival at 1,300m and then gradually rose to reach sea-level values one week after arrival at 5,300m. Surprisingly, cGMP levels did not correlate with NO metabolite concentrations, suggesting that multiple pathways modulated NO availability. For example, elevated cGMP at 1,300m, in the face of negligible changes in nitrite and nitrate and markedly reduced plasma nitrosothiols, suggests utilization of S-nitrosoalbumin as a source of NO during (passive) exposure to hypobaric hypoxia during the flight from London to Kathmandu. Moreover, higher nitrite and nitrate concentrations were associated with greater resting oxygen consumption at sea level. However, only nitrate was inversely related to haemoglobin concentration and blood oxygen content while the opposite was true for nitrosothiols (Fig. 2c), suggesting a complex interplay of bioactive NO metabolites affecting oxygen delivery and consumption. Several of these associations dynamically changed during ascent to high altitude. For example, while the correlation between cGMP and resting oxygen saturation gradually increases in slope the positive association of nitrite with oxygen consumption inverts on ascent (Fig. 3a). The latter suggests enhanced extraction of nitrite from the circulation as hypoxia increases.

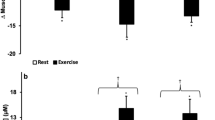
Altitude Dependent Associations between Plasma NO Metabolites, VO 2 peak and Net Efficiency, Relationship between Plasma Nitrite and Microvascular Blood Flow and Sub-group Analyses.
(a) Directional change in associations between peak oxygen consumption (VO2peak) and plasma nitrite at different altitudes (log-transformed values) along with an association matrix of altitude-dependent changes in NO metabolites with net efficiency at 40W. (b) Correlations between microvascular blood flow in small and medium-sized vessels and plasma nitrite as well as cyclic guanosine 3’, 5’-monophosphate (cGMP; insets) concentrations. (c) Subgroup analyses for nitrate, cGMP and erythropoietin (EPO) levels in lab staff (n = 10) and climbers (n = 14); means ± SEM.
Changes in oxygen cost of exercise and microvascular blood flow during ascent
Unexpectedly, the oxygen cost of steady-state exercise (defined as the increase in oxygen consumption per unit increase in work rate in ml O2/min/watt20) was unchanged at altitude. Furthermore, mechanical efficiency expressed as delta efficiency (sea-level: 27.3% (3.0); 5,300m 26.8% (3.2), mean (sd)) and net efficiency (sea-level: 22.3% (1.8); 5,300m 23.3% (3.1)) remained unchanged. Again, complex altitude-dependent patterns of associations were observed between NO metabolites and net efficiency during ascent (Fig. 3a).
As previously reported,19 microvascular blood flow in small (<25 µm diameter) and medium-sized (26–50 µm) vessels is reduced with ascent. Unexpectedly, blood flow in the microcirculation was consistently and inversely correlated with nitrite levels at 3,500m and 5,300m but positively correlated with cGMP at 5,300m (Fig. 3b), consistent with a role for cGMP in blood flow regulation.
Phenotypical differences are apparent already at moderate altitudes
Intriguingly, subgroup analyses comparing the fourteen members of the climbing team (all of whom had significant previous extreme-altitude experience and later ascended above 5,300m9) with the ten base-camp laboratory staff (with less previous altitude exposure and who remained at 5,300m for the duration of the expedition9) revealed higher erythropoietin, nitrate and cGMP levels in the climbers (Fig. 3c). Although not statistically significant, these differences are indicative of higher NOS activity in climbers, apparent already at moderate altitude (3,500m) and may represent evidence of self-selection for higher NO production or epigenetic imprinting from previous altitude exposure.
Discussion
Using an integrated multi-biomarker/whole-body physiology approach in humans, we here demonstrate that NO formation increases in lowlanders ascending to high altitude, in agreement with a role in adaptation to hypoxia. Our findings are consistent with earlier observations of elevated NO metabolite concentrations in the blood of Tibetan highlanders18 and show that enhanced NO production is not unique to this particular population (perhaps as a result of evolutionary selection pressure over millennia) but an integral physiological response to hypoxic stress in humans. In spite of a parallel increase in oxidative stress with hypoxia, in well acclimatised lowlanders not only the production but also the availability of NO is enhanced at all altitudes. Furthermore, we show that enhanced NO levels are associated with changes in microcirculatory blood flow which may affect local tissue oxygen delivery. Contrary to our expectations, we found no evidence for a change in the oxygen cost of exercise. Unchanged oxygen cost with physiological elevation of nitrite in lowlanders at altitude contrasts with reduced oxygen cost of work shown in studies of dietary nitrate supplementation at sea level20,21 and in Tibetans at altitude.23 However, nitrite levels in Tibetan highlanders exceed those observed in our subjects by an order of magnitude,18 raising the possibility that improved mechanical efficiency may have evolved as an adaptation to residence in a hypoxic environment in this population.
In many cases, the biochemical changes in response to high altitude exposure are so striking that almost everything is statistically significant when compared to baseline. However, uncovering the biological relevance of those alterations remains a challenge. Relatively small (and statistically insignificant) changes in the steady-state concentration of biomarkers and signalling molecules may be associated with biologically significant changes in physiology, in particular when regulatory circuits are operating at their limits. In other cases, statistically significant changes from baseline may simply indicate that the system is operating at a different setpoint (which may still be well within the regulatory range). Given the significance of NO in the regulation of so many vital bodily functions, including vascular tone and mitochondrial activity, an enhanced NO availability would seem to be important for sustained local NO signalling under conditions of globally elevated reactive oxygen species production, as we here document to occur on sojourn to high altitude. This seems to be accomplished by a combination of factors, including elevated de novo synthesis by NOS (resulting in higher levels of circulating nitrite and nitrate), release of NO by redox-activation of plasmatic storage forms (as evidenced by the initial drop in S-nitroso species) and reduction of nitrite (indicated by gradual directional changes in association between plasma nitrite concentrations and some of the physiological parameters on exposure to increasing levels of hypoxia). Reduction of nitrite (and possibly also nitrate)16 may also account for our observation that peak nitrite and nitrate levels were not sustained on prolonged stay at high altitude; in addition, it may have limited the magnitude of the increases measured. That elevated NO production indeed translates into enhanced NO bioavailability (in spite of concomitantly elevated reactive oxygen species production) is suggested by the increases in circulating cGMP concentrations we detected at high altitude; in the absence of major changes in natriuretic peptide levels, which could also have contributed to those changes via stimulation of particulate guanylyl cyclase and no reason to suspect an inhibition of phosphodiesterase activity24, this signature is consistent with an enhanced NO activity. Although elevations in circulating cGMP levels appear to be rather robust this does not necessarily translate into increases in blood flow, a prerequisite for enhanced convective delivery of oxygen through the blood. This conclusion is supported by the contrasting associations of microvascular blood flow with circulating nitrite and cGMP levels. Pharmacological doses of nitrite can lower blood pressure,25,26 but whether or not this extends all the way down to physiological levels is currently unknown. Our observation that physiological nitrite concentrations, at sea level and across altitudes, are negatively correlated with blood flow in the microcirculation was unexpected but is consistent with (i) its lack of direct vasodilator activity at low-to-moderate concentrations27 and (ii) the increase in blood pressure at low nitrite doses observed in animals28 and humans.29 Thus, while the positive correlation of microvascular blood flow with cGMP at 5,300m suggests that NO formation (from NOS or nitrite reduction) is enhanced, this appears to be insufficient to normalize reduced microcirculatory blood velocity. An alternative or additional explanation, given that all metabolites were measured in venous plasma, is that nitrite present in arterial blood was metabolized to NO in upstream capillary beds and that the cGMP and blood flow changes are reflections of that process. Paradoxically, if diffusion limitation of oxygen is a critical factor with respect to performance, slower blood flow at high altitude19 may be a beneficial adaptation allowing increased time for oxygen unloading in tissues. Alterations in microcirculatory function have been associated with morbidity and mortality in human sepsis.30 It would seem to be important, therefore, to clarify whether these alterations are indeed unfavourable perturbations of normal physiology or perhaps an intentional feature in tissue hypoxemia and whether or not therapeutic enhancement of microvascular blood flow under these conditions has merit.
Although our results suggest that several different pathways and sources of NO cooperate to enhance NO availability in response to hypoxia, the precise mechanisms that confer enhanced hypoxia tolerance remain unclear. Our study is limited inasmuch as it does not provide information about accompanying changes in NO metabolite levels in vascular tissue, skeletal muscle, or red blood cells, compartments also expected to contribute to the physiological adaptations observed. Moreover, it remains unclear at this stage whether NO itself or a change in NO-related post-translational modification (such as S-nitrosation) of proteins or transcription factors, or all of the above, contribute(s) to beneficial adaptation to hypoxia. Of note, S-nitrosothiols transduce the ventilatory response to hypoxia31 and HIF-1α itself is a target for S-nitrosation.32 Thus, nitrosothiols may not only act as sources of NO (as indicated by the initial drop in S-nitrosoalbumin in the present study) but also as signal transducers,33 and changes in S-nitrosation status may affect many other downstream processes. Although the main objective of our study was to assess the temporal and quantitative changes of the NO pathway in adaptation to hypoxia during ascent to high altitude several intriguing associations between biochemical parameters and physiological variables (e.g., NO production with oxygen consumption at rest, S-nitrosothiols with blood oxygen content and the inverse relationship between nitrate and haemoglobin) at sea-level are worthy of further investigation. Clearly, additional mechanistic investigations are warranted to identify the sources of NO and reactive oxygen species production (i.e. whether they originate in blood cells, vascular tissue and skeletal muscle or are derived from other organs), the sequence of signalling pathways involved and their relationship with tissue phenotypic changes (e.g. microcirculatory blood flow). Nevertheless, our results suggest that the observed changes in microvascular function are likely to involve cGMP-independent effects13 mediated by longer-lived NO metabolites such as nitrosothiols, nitrite and nitrate. Considering the rich chemical biology of NO in relation to its reaction with oxygen and reactive oxygen species34 it is likely that in addition to HIF-1α several other redox-sensitive signalling nodes are involved in the acclimatisation process.
Our findings may be of relevance not only to healthy subjects exposed to hypobaric hypoxia, but also to patients in whom oxygen delivery is limited through disease affecting the heart, lung or vasculature,8 and to the field of developmental biology.35 Cellular hypoxia with consecutive organ dysfunction/damage is a near universal problem in critical illness. Patient management decisions with large resource implications are often based on very limited ‘hard data’ and few biomarkers other than indices of inflammation or acute tissue damage are available to guide this process. Cellular hypoxia may result from anemia, from pulmonary, cardiac micro-and macrovascular pathology, or from abnormal oxygen handling. Whilst tolerance to hypoxia has traditionally been considered to depend on increased convective delivery, efforts to maximize oxygen supply or utilization either provide no benefit or are harmful.36,37 Mechanisms underpinning inter-individual differences in the ability to cope with hypoxia in human (patho)physiology, which ultimately translate into differences in outcome, are largely unexplored - in part because the complex (often multi-system) nature of critical illness represents a major obstacle to mechanistic investigation. The search for prognostic markers in critical illness is particularly challenging due to the multitude of derangements in signalling/metabolic pathways that characterize conditions involving multiple organ failure. Moreover, many of the classical markers are affected by the often aggressive treatment and invasive procedures applied, posing a challenge to untangling cause/effect relationships. An alternative approach might begin with the detailed study of healthy individuals exposed to profound and prolonged hypoxia, where functional heterogeneity can be linked to biological diversity by use of an integrated biochemical/whole-body physiology assessment and translation through application of derived data to the patient population. Thus, systematic identification and study of the molecular signatures of pathways involved in hypoxic adaptation in healthy individuals may hold the key to understanding adaptation to reduced oxygen availability in critically ill patients and help identify novel therapeutic targets. In this regard, our data suggest that individuals unable to mount an adequate intrinsic response to hypoxia may benefit from direct (nitrite, nitrate) or indirect (e.g., small-molecule modulators of NOS expression/activity) manipulation of NO metabolism to activate the physiological program that confers tolerance to hypoxia. The results of the current study also make us wonder whether the findings of greater mortality in septic shock patients treated with a NOS inhibitor a decade ago38 might now perhaps be interpreted as proof-of-concept that survival in stress/hypoxia depends on a sufficient production of NO. Finally, our study raises the intriguing possibility that maximizing arterial oxygen levels (PaO2) in patients who have been exposed to sustained hypoxia may be harmful, if cellular energetics and organ function have become reliant on reductive rather than oxidative pathways of NO generation. This may be particularly pertinent to the neonatal intensive care setting where, after months of embryonic development under conditions of physiological hypoxia, our youngest patients are likely to suffer disproportionately from additional oxidative damage due to a low-capacity anti-oxidative defence system.
Methods
This study was approved by the University College London Research Ethics Committee, in accordance with the Declaration of Helsinki. Verbal and written informed consent was obtained from all subjects.
Expedition setting and ascent profiles
Twenty four healthy subjects (18 male; mean age 35.2 years; age range 19–59 years)who were investigators on the Caudwell Xtreme Everest 2007 research expedition to Mount Everest participated in this study. The design and conduct of this study is described in detail elsewhere.9 All subjects were free of cardiovascular or respiratory disease and took no medication. Subjects were sea level natives who had been resident at sea level for a minimum of 3 months and had not participated in any form of hypoxic training prior to departure for the expedition. The subjects underwent baseline testing in London (altitude 75 m; barometric pressure 754mmHg, 100kPa) before travelling by plane to Kathmandu, Nepal (1,300 m; 650mmHg, 86.7kPa; PiO2 650mmHg, 86.7kPa). From there they flew to Lukla (2,800 m, expedition day 1) and then trekked to Everest Base Camp (EBC; 5,300 m, 404mmHg, 53.8KPa; PiO2 74.7 mmHg, 9.9 kPa), arriving on expedition day 13. The ascent profile is detailed in Fig. 1a. Testing was repeated during the ascent in field laboratories in Kathmandu (1,300 m; day −3 to 0), Namche (3,500 m; day 4 to 6), Pheriche (4,250 m; day 9 to 10) and at EBC (5,300 m; day 15 to 17), with ambient temperatures well controlled during testing. All subjects followed an identical ascent profile during the ascent to EBC and, to standardize hypoxic exposure, were prevented from excursions of more than 300 m from the group altitude at any time. The subjects were divided into two subgroups, Group 1 (laboratory staff) who subsequently remained at EBC for the duration of the expedition (n = 10) and Group 2 (climbers; n = 14) who subsequently ascended above 5,300 m, to a maximum of 8,848 m on Mount Everest. Criteria for participation in the climbing team were previous extreme altitude experience (event-free ascents over 6,500 m), general mountaineering experience (assessed by the expedition leader and climbing leader) and a demonstrated ability to conduct medical research at high altitude. An additional criterion for summit climbers was a previous successful event-free ascent over 8,000 m.
Basic physiological measurements and peripheral oxygen saturation
All subjects completed a physiological and symptom scoring diary for each day of the expedition. The basic physiological profile included resting heart rate, blood pressure (BP), respiratory rate and peripheral arterial oxygen saturation (SpO2), repeated (with the exception of BP) after a standardized 2-min step exercise challenge (CXE step test: 20 cm step, firm base, alternate feet sequentially, 1 step up or down s−1 by electronic metronome). SpO2 was measured using a pulse oximeter (Onyx 9500, Nonin, USA). This device has been previously validated at moderate altitude (maximum of 2,250m) and in acute hypoxia to an SpO2 of approximately 70%; all measurements were taken from the right index finger. The value displayed in the last 10 s of a continuous one minute measurement period at rest was recorded. Data collection for the diary study was completed shortly after waking each morning, before the ingestion of any food or caffeine containing beverages. All subjects rested in a sitting position for a minimum of 5 min prior to recording resting measurements and subjects were blinded to their own measurements to prevent voluntary manipulation of respiratory patterns.
Cardiopulmonary exercise testing and oxygen consumption measurements
Incremental exercise testing to exhaustion was performed to identify the lactate threshold (anaerobic threshold or gas exchange threshold39,40) and peak oxygen consumption (VO2peak). Subjects fasted for 3 hrs prior to the exercise test and refrained from caffeine on the day of testing. The subjects performed an incremental ramp test to the limit of tolerance using an electromagnetically braked cycle ergometer (Lode Corival, Lode, Groningen, The Netherlands) and a breath-by-breath cardiopulmonary exercise testing system (CPET) (Metamax 3b, Cortex, Leipzig, Germany). A ramp slope of 20–35 W/minute was chosen depending on the sex, age and physical fitness of the subjects in order to obtain a test duration of approximately 10–15 min.41 This was kept constant at all altitudes. Resting measurements were recorded for 3 min, followed by 3 min of unloaded cycling followed by initiation of the incremental ramp.42 The limit of tolerance (maximum) was defined as the point at which the subject could not maintain a pedalling cadence of 60 rpm despite encouragement.
A full calibration of the breath-by-breath system was performed before each test using precision gas mixtures (BOC, Guildford UK) (21% oxygen, 0.03% carbon dioxide and 15% oxygen/5% carbon dioxide). The validity of the CPET device was confirmed with a metabolic calibrator (Vacumed, CA) prior to testing in London and in the laboratory in Nepal.43 The accuracy and reliability of the Metamax 3B gas analyzers and flowmeter were established in a hypobaric chamber (RAF Hendon, UK) prior to departure and were demonstrated to correspond to the specifications defined in the American Thoracic Society guidelines.44 The ergometer was calibrated with a torque-based mechanical ergometer calibrator (Vacumed, CA) prior to testing in London and in the laboratory at EBC. Accuracy and reliability of the ergometer were also established in a hypobaric chamber prior to departure. The gas exchange at the lactate threshold was identified firstly using the V-slope method and confirmed using the ventilatory equivalents method. The methods previously described by Whipp et al. were used to identify these points.39 Peak oxygen consumption (VO2peak) was calculated as the average oxygen consumption rate for the individual breaths taken in the final 20 s of the exercise test.
To determine the oxygen cost of moderate intensity exercise and mechanical efficiency, subjects performed three 10 minute constant work rate tests (CWR) below the lactate threshold. The mean oxygen consumption was calculated from the last 5 minutes of each work rate after steady-state had been established. Mechanical efficiency was expressed as delta efficiency (the ratio of the change in energy expenditure to the change in external work) and net efficiency (the ratio of total oxygen consumption less resting oxygen consumption to external work).
Microcirculation assessment
The sublingual microcirculation was visualized at rest by means of sidestream dark-field imaging using the Microscan SDF camera (Microvision Medical, Amsterdam, The Netherlands).19 All images were obtained by a single investigator experienced in this technique. Subjects were required to rest for 10 minutes with a closed mouth prior to the investigation. Microvascular blood flow was assessed using the microcirculatory flow index. Two experienced investigators independently assessed each film to generate an MFI score according to published guidelines.45
Blood sampling and determination of haemoglobin concentration
All blood samples were taken prior to exercise testing and drawn from the antecubital vein into EDTA containers (BD Vacutainer). Haemoglobin (Hb) concentrations in blood were determined immediately after sampling using a handheld photometric device (Hemocue® Whole Blood Haemoglobin System, HemoCue AB, Angelhoim, Sweden), which has been shown to measure venous Hb concentration as accurately as an automated spectrophotometer. Measurement ‘cuvettes’ for the Hemocue were stored according to manufacturer’s recommendations within a thermostatically controlled storage unit for the duration of the expedition.
Calculation of arterial oxygen content
Arterial oxygen content (CaO2) was calculated as the product of Hb concentration, SpO2 and Hufner’s constant [CaO2 = (SaO2 × Hb × 1.39)]. This formula calculates the proportion of oxygen bound to Hb within red blood cells but does not account for dissolved oxygen in plasma. As the latter normally corresponds to <2% of total oxygen carriage in the blood it was considered a reasonable estimation of arterial oxygen content under the study conditions.
Plasma biomarker analyses
Venesection was performed for the measurement of circulating biomarkers. Plasma was separated from blood cells by centrifugation of whole blood at 800xg for 15 min and immediately frozen in 1 ml aliquots in liquid nitrogen. Samples were stored for the duration of the expedition including their transport back to Kathmandu at liquid nitrogen temperatures. They were then transported back to the UK on dry ice and kept at −40°C in a commercial cryostorage facility until analysis.
Oxidative stress markers were analyzed in duplicate directly from EDTA plasma. Isoprostanes (8-iso-prostaglandin F2α) and 4-hydroxy-2-nonenal (HNE) were quantified using direct competitive enzyme immunoassay (Assay Designs, Ann Arbor, MI) and ELISA (OxiSelect HNE-HIS Adduct ELISA kit, Cell Biolabs, San Diego, CA), respectively. Reduced and oxidized glutathione were determined spectrofluorimetrically (DetectX, Arbor Assays, Ann Arbor, MI). Concentrations of cyclic GMP and natriuretic peptides were determined by commercial immunoassay kits (R&D Systems, Abingdon, UK; Biomedica, Vienna, Austria).
NO metabolite concentrations were quantified immediately after thawing of frozen plasma aliquots in the presence of an excess of N-ethylmaleimide (NEM, in PBS; 10 mM final concentration). For the analysis of circulating S-nitrosothiol concentrations aliquots of NEM-treated EDTA plasma were directly injected into a triiodide-containing reaction chamber and the NO produced from the reduction of protein nitroso species was quantified by gas phase chemiluminescence (CLD 77sp, EcoMedics), as described.46 The concentration of S-nitrosothiols in these samples was estimated from the difference in NO signal before and after sample pre-treatment with mercuric chloride. For nitrite/nitrate analysis, NEM-treated samples were deproteinized with ice-cold methanol (1∶1 v/v), cleared by centrifugation and subjected to analysis by high pressure liquid chromatography using a dedicated nitrite/nitrate analyzer (ENO20, Eicom). Sample processing was performed in a staggered fashion to ensure reproducible processing times. All determinations were carried out in duplicate, with repeated daily calibrations and reported values are corrected for background contaminant levels of nitrite/nitrate.
Erythropoietin was quantified by ELISA (Bender MedSystems, Vienna, Austria) and interleukin-6 and TNF-α using XMAP technology (BioPlex 200, Luminex/BioRad). Plasma creatinine and protein concentrations were determined by the Jaffe and Bradford reactions, respectively and plasma osmolality using a micro-osmometer (Model 3320, Advanced Instruments).
References
Penaloza, D. & Arias-Stella, J. The heart and pulmonary circulation at high altitudes: healthy highlanders and chronic mountain sickness. Circulation. 115, 1132–1146 (2007).
West, J. B., Schoene, R. B. & Milledge, J. S. High Altitude Medicine and Physiology (Hodder Arnold, London, 2007).
Calbet, J. A. et al. Why is VO2 max after altitude acclimatization still reduced despite normalization of arterial O2 content? Am J Physiol Regul Integr Comp Physiol. 284, R304–R316 (2003).
Grocott, M. P. et al. Arterial blood gases and oxygen content in climbers on Mount Everest. N Engl J Med. 360, 140–149 (2009).
Beall, C. M. et al. Natural selection on EPAS1 (HIF2alpha) associated with low hemoglobin concentration in Tibetan highlanders. Proc Natl Acad Sci USA. 107, 11459–11464 (2010).
Yi, X. et al. Sequencing of 50 human exomes reveals adaptation to high altitude. Science. 329, 75–78 (2010).
Aragones, J. et al. Deficiency or inhibition of oxygen sensor Phd1 induces hypoxia tolerance by reprogramming basal metabolism. Nat Genet. 40, 170–180 (2008).
Grocott, M., Montgomery, H. & Vercueil, A. High-altitude physiology and pathophysiology: implications and relevance for intensive care medicine. Crit Care. 11, 203 (2007).
Levett, D. Z. et al. Design and conduct of Caudwell Xtreme Everest: an observational cohort study of variation in human adaptation to progressive environmental hypoxia. BMC Med Res Methodol. 10, 98 (2010).
Moncada, S., Palmer, R. M. & Higgs, E. A. Nitric oxide: physiology, pathophysiology and pharmacology. Pharmacol Rev. 43, 109–142 (1991).
Wink, D. A. et al. Mechanisms of the antioxidant effects of nitric oxide. Antioxid Redox Signal. 3, 203–213 (2001).
Jobgen, W. S., Fried, S. K., Fu, W. J., Meininger, C. J. & Wu, G. Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates. J Nutr Biochem. 17, 571–588 (2006).
Taylor, C. T. & Moncada, S. Nitric oxide, cytochrome c oxidase and the cellular response to hypoxia. Arterioscler Thromb Vasc Biol. 30, 643–647 (2010).
Dosek, A., Ohno, H., Acs, Z., Taylor, A. W. & Radak, Z. High altitude and oxidative stress. Respir Physiol Neurobiol. 158, 128–131 (2007).
Brüne, B. & Zhou, J. Hypoxia-inducible factor-1alpha under the control of nitric oxide. Methods Enzymol. 435, 463–478 (2007).
Lundberg, J. O., Weitzberg, E. & Gladwin, M. T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov. 7, 156–167 (2008).
Feelisch, M. et al. Tissue processing of nitrite in hypoxia: an intricate interplay of nitric oxide-generating and -scavenging systems. J Biol Chem. 283, 33927–33934 (2008).
Erzurum, S. C. et al. Higher blood flow and circulating NO products offset high-altitude hypoxia among Tibetans. Proc Natl Acad Sci USA. 104, 17593–17598 (2007).
Martin, D. S. et al. Changes in sublingual microcirculatory flow index and vessel density on ascent to altitude. Exp Physiol. 95, 880–891 (2010).
Bailey, S. J. et al. Dietary nitrate supplementation reduces the O2 cost of low-intensity exercise and enhances tolerance to high-intensity exercise in humans. J Appl Physiol. 107, 1144–1155 (2009).
Larsen, F. J. et al. Dietary inorganic nitrate improves mitochondrial efficiency in humans. Cell Metab. 13, 149–159 (2011).
Moncada, S., Palmer, R. M. & Gryglewski, R. J. Mechanism of action of some inhibitors of endothelium-derived relaxing factor. Proc Natl Acad Sci USA. 83, 9164–9168 (1986).
Marconi, C. Economy of locomotion in high-altitude Tibetan migrants exposed to normoxia. J Physiol. 569, 667–675 (2005).
Spoto, G., Di Giulio, C., Contento, A. & Di Stilio, M. Hypoxic and hyperoxic effect on blood phosphodiesterase activity in young and old rats. Life Sci. 63, PL349–PL353 (1998).
Butler, A. R. & Feelisch, M. Therapeutic uses of inorganic nitrite and nitrate: From the past to the future. Circulation. 117, 2151–2159 (2008).
Cosby, K. et al. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med. 9, 1498–1505 (2003).
Lauer, T. et al. Plasma nitrite rather than nitrate reflects regional endothelial nitric oxide synthase activity but lacks intrinsic vasodilator action. Proc Natl Acad Sci USA. 98, 12814–12819 (2001).
Bryan, N. S. et al. Nitrite is a signaling molecule and regulator of gene expression in mammalian tissues. Nat Chem Biol. 1, 290–297 (2005).
Reichert, E. T. & Mitchell, S. W. On the physiological action of potassium nitrite. Am J Med Sci. 156, 158–180 (1880).
Trzeciak, S. et al. Microcirculatory Alterations in Resuscitation and Shock Investigators. Early microcirculatory perfusion derangements in patients with severe sepsis and septic shock: relationship to hemodynamics, oxygen transport and survival. Ann Emerg Med. 49, 88–98 (2007).
Lipton, A. J. et al. S-nitrosothiols signal the ventilatory response to hypoxia. Nature. 413, 171–174 (2001).
Sumbayev, V. V., Budde, A., Sumbayev, V. V., Budde, A., Zhou, J. & Brüne, B. HIF-1 alpha protein as a target for S-nitrosation. FEBS Lett. 535, 106–112 (2003).
Lima, B., Forrester, M. T., Hess, D. T. & Stamler, J. S. S-nitrosylation in cardiovascular signaling. Circ Res. 106, 633–646 (2010).
Thomas, D. D. et al. The chemical biology of nitric oxide: implications in cellular signaling. Free Radic Biol Med. 45, 18–31 (2008).
Martin, D. S., Khosravi, M., Grocott, M. P. & Mythen, M. G. Concepts in hypoxia reborn. Crit Care. 14, 315 (2010).
Hayes, M. A. et al. Elevation of systemic oxygen delivery in the treatment of critically ill patients. N Engl J Med. 330, 1717–1722 (1994).
Gattinoni, L. et al. A trial of goal-oriented hemodynamic therapy in critically ill patients. SvO2 Collaborative Group. N Engl J Med. 333, 1025–1032 (1995).
Lopez, A. et al. Multiple-center, randomized, placebo-controlled, double-blind study of the nitric oxide synthase inhibitor 546C88: effect on survival in patients with septic shock. Crit Care Med. 32, 21–30 (2004).
Whipp, B. J., Ward, S. A. & Wasserman, K. Respiratory markers of the anaerobic threshold. Adv Cardiol. 35, 47–64 (1986).
Beaver, W. L., Wasserman, K. & Whipp, B. J. A new method for detecting anaerobic threshold by gas exchange. J Appl Physiol. 60, 2020–2027 (1986).
Hansen, J. E., Sue, D. Y. & Wasserman, K. Predicted values for clinical exercise testing. Am Rev Respir Dis. 129, S49–S55 (1984).
Whipp, B. J., Davis, J. A., Torres, F. & Wasserman, K. A test to determine parameters of aerobic function during exercise. J Appl Physiol. 50, 217–221 (1981).
Huszczuk, A., Whipp, B. J. & Wasserman, K. A respiratory gas exchange simulator for routine calibration in metabolic studies. Eur Respir J. 3, 465–468 (1990).
ATS/ACCP Statement on cardiopulmonary exercise testing.. Am J Resp Crit Care Med. 167, 211–277 (2003).
De Backer, D. et al. How to evaluate the microcirculation: report of a round table conference. Crit Care. 11, R101 (2007).
Rassaf, T., Bryan, N. S., Kelm, M. & Feelisch, M. Concomitant presence of N-nitroso and S-nitroso proteins in human plasma. Free Radic Biol Med. 33, 1590–1596 (2002).
Acknowledgements
Caudwell Xtreme Everest (CXE) is a research project coordinated by the Centre for Altitude, Space and Extreme Environment Medicine, University College London, UK. The aim of CXE is to conduct research into hypoxia and human performance at high altitude in order to improve understanding of hypoxia in critical illness. The CXE Research Groups members all contributed to the collection of the data described in this study. Membership, roles and responsibilities of the CXE Research Group can be found at http://www.caudwell-xtreme-everest.co.uk/team. This research was funded from a variety of sources, most of which are not public: Mr John Caudwell, BOC Medical (now part of Linde Gas therapeutics), Eli Lilly Critical Care, The London Clinic, Smiths Medical, Deltex Medical and The Rolex Foundation (unrestricted grants). Peer-reviewed research grants were awarded by the Association of Anaesthetists of Great Britain and Ireland (AAGBI), the UK Intensive Care Foundation and the Sir Halley Stewart Trust. The CXE volunteers who trekked to Everest base camp also kindly donated to support the research. Some of this work was undertaken at University College Hospitals - University College London Comprehensive Biomedical Research Centre which received a portion of funding from the UK Department of Health Research Biomedical Research Centres funding scheme. Part of the work carried out at the University of Warwick was supported by funds from the Medical Research Council (Strategic Appointment Scheme, to MF). None of the funding bodies or the institutions the authors are affiliated with had any role in the study design; collection, analysis and interpretation of data; manuscript preparation, or decision to publish.
Author information
Authors and Affiliations
Consortia
Contributions
MPG, DZL, DSM, HEM, KM, MGM and MF: designed research; DZL, BOF, HLR, DSM, KM, CAL, MGM: performed research, DZL, BOF, HLR, DSM, CI, KM, BJW, CAL, MPG and MF: analyzed data; and MF, DL and MPG wrote the paper. All authors read and approved the final version of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Levett, D., Fernandez, B., Riley, H. et al. The role of nitrogen oxides in human adaptation to hypoxia. Sci Rep 1, 109 (2011). https://doi.org/10.1038/srep00109
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00109
This article is cited by
-
HighAltitudeOmicsDB, an integrated resource for high-altitude associated genes and proteins, networks and semantic-similarities
Scientific Reports (2023)
-
Toxicological and histopathological alterations in the heart of young and adult albino rats exposed to mosquito coil smoke
Environmental Science and Pollution Research (2023)
-
A quantitative proteomic approach revealed variations of salivary proteome during high altitude de-acclimatization
Journal of Proteins and Proteomics (2023)
-
Sympathetic neurovascular transduction following acute hypoxia
Clinical Autonomic Research (2021)
-
Biofluids in hypobaric hypoxia: best possible use, investigative strategies and putative markers
Journal of Proteins and Proteomics (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
