
Abstract
Objective:
A sexual dimorphism exists in body fat distribution; females deposit relatively more fat in subcutaneous/inguinal depots whereas males deposit more fat in the intra-abdominal/gonadal depot. Our objective was to systematically document depot- and sex-related differences in the accumulation of adipose tissue and gene expression, comparing differentially expressed genes in diet-induced obese mice with mice maintained on a chow diet.
Research Design and Methods:
We used a microarray approach to determine whether there are sexual dimorphisms in gene expression in age-matched male, female or ovariectomized female (OVX) C57/BL6 mice maintained on a high-fat (HF) diet. We then compared expression of validated genes between the sexes on a chow diet.
Results:
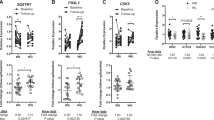
After exposure to a high fat diet for 12 weeks, females gained less weight than males. The microarray analyses indicate in intra-abdominal/gonadal adipose tissue in females 1642 genes differ by at least twofold between the depots, whereas 706 genes differ in subcutaneous/inguinal adipose tissue when compared with males. Only 138 genes are commonly regulated in both sexes and adipose tissue depots. Inflammatory genes (cytokine–cytokine receptor interactions and acute-phase protein synthesis) are upregulated in males when compared with females, and there is a partial reversal after OVX, where OVX adipose tissue gene expression is more ‘male-like’. This pattern is not observed in mice maintained on chow. Histology of male gonadal white adipose tissue (GWAT) shows more crown-like structures than females, indicative of inflammation and adipose tissue remodeling. In addition, genes related to insulin signaling and lipid synthesis are higher in females than males, regardless of dietary exposure.
Conclusions:
These data suggest that male and female adipose tissue differ between the sexes regardless of diet. Moreover, HF diet exposure elicits a much greater inflammatory response in males when compared with females. This data set underscores the importance of analyzing depot-, sex- and steroid-dependent regulation of adipose tissue distribution and function.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
Dua A, Hennes MI, Hoffman RG, Maas DL, Krakower GR, Sonnenberg GE et al. Leptin: a significant indicator of total body fat but not of visceral fat and insulin insensitivity in African-American women. Diabetes 1996; 45: 1635–1637.
Havel PJ, Kasim-Karakas S, Dubuc GR, Mueller W, Phinney SD . Gender differences in plasma leptin concentrations. Nat Med 1996; 2: 949–950.
Kotani K, Tokunaga K, Fujioka S, Kobatake T, Keno Y, Yoshida S et al. Sexual dimorphism of age-related changes in whole-body fat distribution in the obese. Int J Obes Relat Metab Disord 1994; 18: 207–202.
Legato MJ . Gender-specific physiology: how real is it? How important is it? Int J Fertil Womens Med 1997; 42: 19–29.
Wajchenberg BL . Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev 2000; 21: 697–738.
Clegg DJ, Brown LM, Woods SC, Benoit SC . Gonadal hormones determine sensitivity to central leptin and insulin. Diabetes 2006; 55: 978–987.
Gambacciani M, Ciaponi M, Cappagli B, Piaggesi L, De Simone L, Orlandi R et al. Body weight, body fat distribution, and hormonal replacement therapy in early postmenopausal women. J Clin Endocrinol Metab 1997; 82: 414–417.
Haarbo J, Hansen BF, Christiansen C . Hormone replacement therapy prevents coronary artery disease in ovariectomized cholesterol-fed rabbits. Apmis 1991; 99: 721–727.
Haarbo J, Marslew U, Gotfredsen A, Christiansen C . Postmenopausal hormone replacement therapy prevents central distribution of body fat after menopause. Metabolism 1991; 40: 1323–1326.
Lovejoy JC, Sainsbury A . Sex differences in obesity and the regulation of energy homeostasis. Obes Rev 2009; 10: 154–167.
Despres JP . The insulin-resistance-dyslipidemic syndrome of visceral obesity: effect on patients’ risk. Obes Res 1998; 6 (Suppl 1): 8S–17S.
Lamarche B . Abdominal obesity and its metabolic complications: implications for the risk of ischaemic heart disease. Coron Artery Dis 1998; 9: 473–481.
Van Pelt RE, Jankowski CM, Gozansky WS, Schwartz RS, Kohrt WM . Lower-body adiposity and metabolic protection in postmenopausal women. J Clin Endocrinol Metab 2005; 90: 4573–4578.
Edens NK, Fried SK, Kral JG, Hirsch J, Leibel RL . In vitro lipid synthesis in human adipose tissue from three abdominal sites. Am J Physiol 1993; 265: E374–E379.
Fried SK, Leibel RL, Edens NK, Kral JG . Lipolysis in intraabdominal adipose tissues of obese women and men. Obes Res 1993; 1: 443–448.
Jensen MD . Health consequences of fat distribution. Horm Res 1997; 48 (Suppl 5): 88–92.
Shadid S, Koutsari C, Jensen MD . Direct free fatty acid uptake into human adipocytes in vivo: relation to body fat distribution. Diabetes 2007; 56: 1369–1375.
Tchernof A, Belanger C, Morisset AS, Richard C, Mailloux J, Laberge P et al. Regional differences in adipose tissue metabolism in women: minor effect of obesity and body fat distribution. Diabetes 2006; 55: 1353–1360.
D’Eon TM, Souza SC, Aronovitz M, Obin MS, Fried SK, Greenberg AS . Estrogen regulation of adiposity and fuel partitioning. Evidence of genomic and non-genomic regulation of lipogenic and oxidative pathways. J Biol Chem 2005; 280: 35983–35991.
Friedman JM, Halaas JL . Leptin and the regulation of body weight in mammals. Nature 1998; 395: 763–770.
Guerre-Millo M . Adiponectin: an update. Diabetes Metab 2008; 34: 12–18.
Arner P . Differences in lipolysis between human subcutaneous and omental adipose tissues. Ann Med 1995; 27: 435–438.
Bolinder J, Engfeldt P, Ostman J, Arner P . Site differences in insulin receptor binding and insulin action in subcutaneous fat of obese females. J Clin Endocrinol Metab 1983; 57: 455–461.
Hellmer J, Marcus C, Sonnenfeld T, Arner P . Mechanisms for differences in lipolysis between human subcutaneous and omental fat cells. J Clin Endocrinol Metab 1992; 75: 15–20.
Macotela Y, Boucher J, Tran TT, Kahn CR . Sex and depot differences in adipocyte insulin sensitivity and glucose metabolism. Diabetes 2009; 58: 803–812.
Ostman J, Arner P, Engfeldt P, Kager L . Regional differences in the control of lipolysis in human adipose tissue. Metabolism 1979; 28: 1198–1205.
Tordjman J, Guerre-Millo M, Clement K . Adipose tissue inflammation and liver pathology in human obesity. Diabetes Metab 2008; 34: 658–663.
Dicker A, Ryden M, Naslund E, Muehlen IE, Wiren M, Lafontan M et al. Effect of testosterone on lipolysis in human pre-adipocytes from different fat depots. Diabetologia 2004; 47: 420–428.
Heldring N, Pike A, Andersson S, Matthews J, Cheng G, Hartman J et al. Estrogen receptors: how do they signal and what are their targets. Physiol Rev 2007; 87: 905–931.
Hoffstedt J, Arner P, Hellers G, Lonnqvist F . Variation in adrenergic regulation of lipolysis between omental and subcutaneous adipocytes from obese and non-obese men. J Lipid Res 1997; 38: 795–804.
Nilsson M, Dahlman I, Ryden M, Nordstrom EA, Gustafsson JA, Arner P et al. Oestrogen receptor alpha gene expression levels are reduced in obese compared to normal weight females. Int J Obes (Lond) 2007; 31: 900–907.
Boivin A, Brochu G, Marceau S, Marceau P, Hould FS, Tchernof A . Regional differences in adipose tissue metabolism in obese men. Metabolism 2007; 56: 533–540.
Boyns DR, Crossley JN, Abrams ME, Jarrett RJ, Keen H . Oral glucose tolerance and related factors in a normal population sample. I. Blood sugar, plasma insulin, glyceride, and cholesterol measurements and the effects of age and sex. Br Med J 1969; 1: 595–598.
Li AC, Brown KK, Silvestre MJ, Willson TM, Palinski W, Glass CK . Peroxisome proliferator-activated receptor gamma ligands inhibit development of atherosclerosis in LDL receptor-deficient mice. J Clin Invest 2000; 106: 523–531.
Strissel KJ, Stancheva Z, Miyoshi H, Perfield 2nd JW, DeFuria J, Jick Z et al. Adipocyte death, adipose tissue remodeling, and obesity complications. Diabetes 2007; 56: 2910–2918.
Xiao XQ, Grove KL, Smith MS . Metabolic adaptations in skeletal muscle during lactation: complementary deoxyribonucleic acid microarray and real-time polymerase chain reaction analysis of gene expression. Endocrinology 2004; 145: 5344–5354.
Cinti S, Mitchell G, Barbatelli G, Murano I, Ceresi E, Faloia E et al. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J Lipid Res 2005; 46: 2347–2355.
Doniger S, Hofmann T, Yeh J . Predicting CNS permeability of drug molecules: comparison of neural network and support vector machine algorithms. J Comput Biol 2002; 9: 849–864.
Guerre-Millo M, Leturque A, Girard J, Lavau M . Increased insulin sensitivity and responsiveness of glucose metabolism in adipocytes from female versus male rats. J Clin Invest 1985; 76: 109–116.
Yki-Jarvinen H . Sex and insulin sensitivity. Metabolism 1984; 33: 1011–1015.
Frias JP, Macaraeg GB, Ofrecio J, Yu JG, Olefsky JM, Kruszynska YT . Decreased susceptibility to fatty acid-induced peripheral tissue insulin resistance in women. Diabetes 2001; 50: 1344–1350.
Hevener A, Rei D, Janez A, Olefsky J . Female rats do not exhibit free fatty acid-induced insulin resistance. Diabetes 2002; 51: 1907–1912.
Soeters MR, Sauerwein HP, Groener JE, Aerts JM, Ackermans MT, Glatz JF et al. Gender-related differences in the metabolic response to fasting. J Clin Endocrinol Metab 2007; 92: 3646–3652.
Corsetti JP, Sparks JD, Peterson RG, Smith RL, Sparks CE . Effect of dietary fat on the development of non-insulin dependent diabetes mellitus in obese Zucker diabetic fatty male and female rats. Atherosclerosis 2000; 148: 231–241.
Trevaskis JL, Meyer EA, Galgani JE, Butler AA . Counterintuitive effects of double-heterozygous null melanocortin-4 receptor and leptin genes on diet-induced obesity and insulin resistance in C57BL/6J mice. Endocrinology 2008; 149: 174–184.
Zierath JR, Houseknecht KL, Gnudi L, Kahn BB . High-fat feeding impairs insulin-stimulated GLUT4 recruitment via an early insulin-signaling defect. Diabetes 1997; 46: 215–223.
Geisler RW, Hansen RJ . Effects of insulin on the adaptation of glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase in rat adipose tissue. Biochim Biophys Acta 1972; 279: 139–145.
Koutsari C, Snozek CL, Jensen MD . Plasma NEFA storage in adipose tissue in the postprandial state: sex-related and regional differences. Diabetologia 2008; 51: 2041–2048.
Kjeldsen L, Cowland JB, Borregaard N . Human neutrophil gelatinase-associated lipocalin and homologous proteins in rat and mouse. Biochim Biophys Acta 2000; 1482: 272–283.
Yan QW, Yang Q, Mody N, Graham TE, Hsu CH, Xu Z et al. The adipokine lipocalin 2 is regulated by obesity and promotes insulin resistance. Diabetes 2007; 56: 2533–2540.
Kehrer JP . Lipocalin-2: pro- or anti-apoptotic? Cell Biol Toxicol 2009.
van Nas A, Guhathakurta D, Wang SS, Yehya N, Horvath S, Zhang B et al. Elucidating the role of gonadal hormones in sexually dimorphic gene coexpression networks. Endocrinology 2009; 150: 1235–1249.
Acknowledgements
We thank the ISIS network for their funding and support of the research. We also thank the members of the ISIS group: Jeffrey Chang, Jennifer Lovejoy, Nori Geary, Joel Elmquist, Philipp Scherer, Randy Seeley, Richard Simerly and Steve Smith. For technical assistance, we thank Sarah Williams, salary support RR00163 (KLG), as well as Jody Caldwell and Kathi Smith.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on International Journal of Obesity website
Supplementary information
Rights and permissions
About this article
Cite this article
Grove, K., Fried, S., Greenberg, A. et al. A microarray analysis of sexual dimorphism of adipose tissues in high-fat-diet-induced obese mice. Int J Obes 34, 989–1000 (2010). https://doi.org/10.1038/ijo.2010.12
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ijo.2010.12
Keywords
This article is cited by
-
Interaction between gut microbiota and sex hormones and their relation to sexual dimorphism in metabolic diseases
Biology of Sex Differences (2023)
-
Maternal obesity and programming of metabolic syndrome in the offspring: searching for mechanisms in the adipocyte progenitor pool
BMC Medicine (2023)
-
Sex differences in hearing impairment due to diet-induced obesity in CBA/Ca mice
Biology of Sex Differences (2023)
-
Branched-chain amino acid catabolism in muscle affects systemic BCAA levels but not insulin resistance
Nature Metabolism (2023)
-
Obesogenic and diabetic effects of CD44 in mice are sexually dimorphic and dependent on genetic background
Biology of Sex Differences (2022)
