
Abstract
Arthropods produce a great variety of natural compounds, many of which have unexplored biosynthesis. Among the armored harvestmen (Arachnida: Opiliones) of the suborder Laniatores, the defensive gland exudates contain vinyl ketones and other constituents of supposed polyketide origin. We have studied the biosynthesis of 1-hepten-3-one in the Neotropical harvestman Iporangaia pustulosa by feeding individuals with 13C-labeled precursors, demonstrating its mixed acetate/propionate origin. 13C NMR spectroscopy showed an unusual labeling pattern suggesting different propionate sources for starting and extender units. Our analysis also indicates the presence of methylmalonyl-CoA mutase, converting acetate into propionyl-CoA via succinyl-CoA, together with other C3 unit routes. This is the first biosynthetic study of alkyl vinyl ketones in arthropods. Our results shed light on the origin and diversification of chemical compounds in a major arthropod group.
Similar content being viewed by others
Introduction
Arthropods produce a great variety of natural compounds that are used in inter- and intra-species chemical communication. Although the identity of these info-chemicals have been intensively explored over the last 60 years1, their biosynthesis remains vastly unexplored when compared to plants and microorganisms2. Moreover, most biosynthetic studies with arthropods focus on insects and only a few on other major arthropod groups such as arachnids, whose species may produce venoms, repellent exudates and pheromones1. Arachnids of the order Opiliones, for instance, have a pair of exocrine glands that release a great variety of odorous compounds3 (Fig. 1) used as alarm pheromones4 and defense against predators5,6,7. However, their biosynthetic pathways have never been investigated.

Female of the harvestman Iporangaia pustulosa releasing two white droplets of scent gland exudate in the lateral margins of the carapace.
The photo was taken by Prof. Glauco Machado.
Among the armored harvestmen of the suborder Laniatores, the gland exudates contain mixtures of benzoquinones, alkylated phenols, or alkyl vinyl ketones8,9. Considering the phylogeny of the Neotropical family Gonyleptidae, in particular, the production of benzoquinones is the ancestral state, with at least four independent losses of these compounds10. The production of vinyl ketones, on the other hand, is derived and represents a synapomorphy of a major clade composed of five subfamilies and the production of alkyl-phenols is also derived and evolved at least five times independently10. The biosynthetic pathways of these gland exudate constituents in harvestmen may bring information on the putative origins and diversification of chemical compounds within this major arachnid group.
In a previous report, we detected 1-hepten-3-one (1) as the major component in the scent gland exudates of the gonyleptid harvestman Iporangaia pustulosa9. Similar vinyl ketones were also found in other gonyleptid harvestmen belonging to the clade known as K928,10. Here we describe the biosynthetic pathway of 1, revealing the condensation of Pr + Ac + Pr units to produce a polyketide chain. This pathway depends on a methylmalonyl-CoA mutase and other enzymes comprising an ensemble of parallel propionate/methylmalonate metabolic routes. Further, we revealed that there are different sources of C3 depending on the unit role of starter or extender. To our knowledge, this is the first biosynthetic study of an alkyl vinyl ketone in arthropods.
Results
Feeding and analysis
Four 13C-labeled precursors, denoted [13C3]propionate, [4-13C]methylmalonate, [1-13C]acetate and [1-13C]glucose, were added to the diet of Iporangaia individuals (males and females) and their incorporation into 1 was monitored by 13C NMR spectroscopy. The vinyl ketone 1 produced by Iporangaia individuals in each feeding experiment showed specific incorporation of the precursors.
[13C3]propionate incorporation
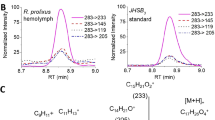
The enrichment by [13C3]propionate is given by the integration ratio of the satellite lines by the sum of all carbon signals11,12 ( Fig. 2 , Table 1 ). Positions C-1, C-2, C-5, C-6 and C-7 were labeled and at C-1 and C-2 the enrichment was weaker. All labeled positions showed increased satellite lines.

Results of the experiment in which Iporangaia pustulosa individuals were fed [13C3]propionate.
(a) 13C NMR spectrum (125 MHz, CDCl3) of the exudate containing 1 as major compound. (b) Extended signals of 13C enriched positions showing 13C-13C coupling patterns.
[4-13C]methylmalonate, [1-13C]acetate and [1-13C]glucose incorporation
For each carbon signal of 1 in the 13C NMR spectrum, the ratio of the signal height was calculated using one non-enriched position (C-4 for [1-13C]acetate and [4-13C]methylmalonate; C-5 for [1-13C]glucose), furnishing the R value. The r = Rlabeled group/Rcontrol group was calculated for all the positions of 1. The enrichment at each site was obtained by multiplying r by 1.1, which is the natural abundance of 13C ( Table 2 )11,12. Experiment with [4-13C]methylmalonate produced 1 enriched at C-1 (1.3%), C-6 (1.4%) and C-7 (1.5%) (Supplementary Data S3). Individuals fed on [1-13C]acetate produced 1 showing increased intensities at C-1 (2.5%), C-3 (3.0%), C-5 (1.6%) and C-6 (1.3%) (Supplementary Data S4). [1-13C]glucose incorporation yielded 1 with isotopic enhancement at C-2, C-4, C-6 and C-7 of 1.3% (Supplementary Data S5), with a lower global incorporation and 13C NMR signal enhancement than observed in [1-13C]acetate feeding experiment. The sample was too diluted and the carbonyl signal was not observed.
Discussion
Given that the biosynthesis of secondary metabolites of harvestmen had never been studied before, we used Morgan's suggestion2 of an aceto-propiogenin origin for aliphatic ketones structurally similar to 1 as a first approach. To test this hypothesis, we selected 13C NMR spectroscopy and 13C-labeled precursors (acetate and propionate), which allowed the exact position of the unit incorporation into the intact molecule of 1. Iporangaia pustulosa has a defense secretion with 1 as major constituent, allowing 13C NMR analysis of the crude exudate, with the additional advantage of surviving feeding experiments in the laboratory.
Feeding Iporangaia individuals on [13C3]propionate yielded 1 labeled at C-1, C-2, C-5, C-6 and C-7 (Table 1). The 13C enrichment increased the intensities of the satellites and reduced the central signal of the singly labeled isotopomers. The doublets at C-7 and C-5 and double doublet at C-6 have characteristic 13C-13C coupling constants, revealing the intact incorporation of the [13C3]propionate precursor at these positions (Fig. 2, Table 2). Satellite signals (doublets) were also enhanced at C-1 and C-2 indicating the incorporation of a second [13C3]propionate unit and loss of one labeled carbon. The 13C enrichment at C-1 and C-2 (~15%) was less effective than at C-5, C-6 and C-7 (~44%), suggesting alternative C3 units for initiating (propionate) and extending (methylmalonate) the polyketide chain of 1. In the same way, [4-13C]methylmalonate incorporation yielded labelling at C-1 and C-7, in agreement with two C3 units incorporated into the polyketide chain of 1. C-3 and C-4 showed no 13C incorporation from C3 labeled precursors, suggesting that these two carbons arise from an acetate unit. In fact, when [1-13C]acetate was incorporated into the diet, the C-3 signal increased 3% (Table 1) and [1-13C]glucose incorporation enriched C-4 in 1.3% (Table 1). This last result was expected as [1-13C]glucose is first metabolized producing two acetate units, one unlabelled and one labelled at C-2 after a TCA, cycle yielding a weaker enrichment.
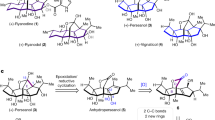
This labeling pattern indicates that biosynthesis of 1 occurs via polyketide synthases (PKS) notwithstanding the lack of evidence of their presence in arthropods (see review in ref. 13). On the basis of the PKS mode of action14 we propose assembling to initiate with a propionyl-CoA (C3), which is extended by a malonyl-CoA (C2) and a methylmalonyl-CoA (C3) (Fig. 3). Hydrolysis of the thioester and decarboxylation yields the saturated ketone precursor of 1 (Fig. 3c). The introduction of the terminal double bond of 1 can be assigned to an unusual desaturase (Fig. 3d), but this mechanism was not assessed in our experiments. Known desaturases in insects do not insert double bonds at the terminal nor at α, β-carbonyl positions2. A saturated analog of 1 (4-methyl-3-hexanone) was isolated from the ant Harpegnathos saltator and also revealed a mixed acetate-propionate origin by heavy isotope labeling15. Among arachnids, the storage mite Chortoglyphus arcuatus was shown to adds one acetate and four propionate units to produce the aggregation pheromone (4R,6R,8R)-4,6,8-trimethyldecan-2-one (chortolure)16.

Proposed biosynthetic pathway for hepten-3-one (1) from the harvestman Iporangaia pustulosa.
(a) Condensation of propionyl-CoA with malonate. (b) Condensation with methylmalonyl-CoA. (c) Reduction, dehydration, reduction, thioester hydrolysis and decarboxylation. (d) Dehydrogenation of the saturated ketone.
A closer examination of the 13C NMR spectra from labeling experiments with [4-13C]methylmalonate, [1-13C]acetate and [1-13C]glucose revealed intriguing labeling patterns relative to the C3 units (Fig. 4). The unexpected labeling of 1 at C-6, by adding [4-13C]methylmalonate to the diet of Iporangaia individuals suggests the existence of a methylmalonyl-CoA mutase. The isotopic enhancement at positions C-6 (1.5%) and C-7 (2.3%) complies with the methylmalonyl-CoA–succinyl-CoA interconversion, leading to [2-13C] and [3-13C]propionyl-CoA biosynthesis as consequence of the equilibrium succinyl-CoA-succinate17 (Fig. 5a1). Stronger enrichment at C-7 indicates the incorporation of [3-13C]propionyl-CoA and the enrichment at C-1 (1.9%) is provided by direct incorporation of [4-13C]methylmalonyl-CoA as the second extender unit (Fig. 5c). Moreover, labeling of 1 at propionate units with [1-13C]acetate incorporation (Fig. 4) suggests the conversion of acetate into propionate/methylmalonate. The presence of methylmalonyl-CoA would yield [1-13C]methylmalonyl-CoA (Fig. 5c), undetectable due to the loss of labeled carbon, as depicted in Figure 3c. Methylmalonyl-CoA mutase was previously reported in polyketide biosynthesis of insects18,19,20 and other arthropods21, suggesting that 1 from I. pustulosa possesses similar biosynthetic origin.

Labeling pattern of 1 by feeding Iporangaia pustulosa individuals with 13C labeled precursors.
Colored ball indicate 13C labeled positions. Green signals means expected and magenta means unexpected labeling. The ball sizes correspond to 13C enrichment (%).

Proposed biosynthesis of 1 from the harvestman Iporangaia pustulosa based on [4-13C]methylmalonate feeding experiment and observed labelling pattern.
Isotopomers of 1 and precursor are not indicated separately.
Enrichment at C-5 (1.6%) and C-6 (1.3%) are similar with the [1-13C]acetate feeding, suggesting that both labelings arise from the same pool of starting units, either [1-13C] or [2-13C]propionyl-CoA. The biosynthesis of the C3 unit from C2 via α-oxidation and decarboxylation of butyrate has been reported for pheromone biosynthesis of Carpophilus beetles (Nitidulidae)22, however this mechanism is not in accordance with the observed labeling pattern. Chan et al.23 reviewed the sources of propionate in several microorganisms and, in the crotonyl-CoA carboxylase/reductase pathway, the acetate is converted into propionate after condensation of two acetyl-CoA units yielding a butyrate24. The presence of a crotonyl-CoA mutase pathway in I. pustulosa would explain [1-13C]acetate labeling of 1 at C-5 and C-6, which corresponds to [1-13C] and [3-13C]propionate incorporation (Fig. 6a). Enrichment at C-1 of 1 by feeding on [1-13C]acetate corresponds to the incorporation of a [3-13C]methylmalonyl-CoA extender unit, whose origin can be linked to methionine and/or threonine catabolism via acetate (Fig. 6c).

Proposed biosynthesis of 1 from the harvestman Iporangaia pustulosa based on [1-13C]acetate feeding experiment and observed labelling pattern.
Isotopomers of 1 and precursors are not indicated separately.
[4-13C]methylmalonate incorporation enriches positions C-6 and C-7 of 1, corresponding to a C3 starter unit while the C3 extender unit labels only C-1 and not C-2, (Fig. 5). This labeling pattern indicates the existence of parallel propionate routes in I. pustulosa for the C3 units as starter and extender to be loaded during the polyketide chain assembly. Analyzing [1-13C]acetate incorporation, the labeling at C-6 and non-labeling at C-2 (Fig. 6) are further evidences of these parallel route. Additionally, the [13C3]propionate 13C enrichment at C-1 and C-2 (15.25 and 16.27%, respectively) corresponding to the extender unit is weaker than at C-5, C-6 and C-7 (39.18%, 48.63% and 45.06%, respectively), the starter unit, which reinforces the hypothesis of parallel propionate route (Table 2).
Feeding on [4-13C]methylmalonate yields no labeling at C-3 nor at C-4 of 1, corresponding to the acetate portion of 1 polyketide chain. Consequently, we can say that harvestman propionate catabolism occurs via succinate, contrary to the alternative biosynthetic pathway via 3-hydroxypropionate to acetate for insects25,26. Our results also indicate that propionate in harvestmen may arise from three alternative biosynthetic pathways (succinyl-CoA, crotonyl-CoA and methionine and/or threonine catabolism) (Fig. 7). Bacterial symbionts are known to participate in the biosynthesis of arthropod secondary metabolites27,28 and may be an alternative source of C3 units for the biosynthesis of harvestman defense molecules like 1.

Propionate metabolism in the harvestman Iporangaia pustulosa.
Given that the production of vinyl ketones is a synapomorphy of the clade composed of the subfamilies Gonyleptinae, Hernandariinae, Sodreaninae, Progonyleptoidellinae and Caelopyginae10, the biosynthetic pathway described here is probably similar in other harvestman species belonging to this major group of gonyleptids. Moreover, given that the production of vinyl ketones is derived from an ancient state of the benzoquinone production in gonyleptid harvestmen10, our next challenge is to investigate whether distantly related species partially share the biosynthetic pathways of these two major classes of metabolites. Previous studies with tenebrionid beetles have shown that two 1,4-benzoquinones also reported in the scent gland secretion of several gonyleptid species (2-methyl-1,4-benzoquinone and 2-ethyl-1,4-benzoquinone) are derived from a pathway using acetate and propionate as precursors, suggesting a polyketide origin29,30. Therefore, similar precursors for both benzoquinones and vinyl ketones could help to explain evolutionary transitions among gonyleptid species.
Methods
Harvestmen
Individuals of Iporangaia pustulosa were collected from a large fragment of Atlantic forest in the state of São Paulo, southeastern Brazil. The collection of the animals in the field and the experiments in the laboratory complied with the current laws of the Brazilian government. Before beginning the experiment, a dorso-ventral pressure was applied on all individuals to empty the gland sacs. The pool of harvestmen was divided into four groups and fed on: 1) canned sardines with 5% w/w [1-13C]sodium acetate (Cambridge Isotope Laboratories, CIL) (n = 14 individuals); 2) canned sardines with 5% w/w [4-13C]sodium methylmalonate (Supplementary Method S7) (n = 11 individuals); 3) canned sardines with 1% w/w [1-13C]glucose (CIL) (n = 10 individuals) and 4) [13C3] propionate (Aldrich) (n = 28 individuals). The control group was the exudate extracted before initiating each experiment (n = 30 individuals). The experiment was set up over a period of 60 days (for groups 1 to 3) and 30 days for group 4, with feeding renewal every 48 h. The number of individuals analyzed in each experimental group is the total that survived for the whole period of feeding (minimum of 10 individuals). The gland exudates were collected with dewaxed cotton wool and extracted from the cotton wool with deuterochloroform or deuterobenzene.
NMR analyses of 1
13C NMR signal assignment of 1 has been described in the literature31. Previous assignments were refined with 2D-NMR spectra (1H, 13C HSQC) (Supplementary Data S9) of the synthetic standard9. NMR spectra of 1 were acquired with either a Bruker Avance III 11 tesla (for [1-13C]acetate, [4-13C]methylmalonate and [13C3]propionate groups) or a Varian Inova spectrometer 11 tesla ([1-13C]glucose group), both operating at 125.75 MHz, 25°C, acquisition time 0.55 s and collecting about 40,000 scans, taking care to have equal scan numbers for samples belonging to the same experiment (sample and control).
References
Eisner, T., Eisner, M. & Seigler, M. Secret Weapons: Defenses of Insects, Spiders, Scorpions and Other Many-Legged Creatures, (Belknap Press of Harvard University Press, 2005).
Morgan, E. D. Biosynthesis in Insects - Advanced Edition, (RSC, Cambridge 2010).
Gnaspini, P. & Hara, M. R. Defense Mechanisms., in: Pinto-da-Rocha, R., Machado, G. & Giribet, G. (Eds.), Harvestmen: the Biology of Opiliones, 374–399 (Harvard University Press, 2007).
Machado, G., Bonato, V. & Oliveira, P. S. Alarm communication: a new function for the scent gland secretion in harvestmen (Arachnida: Opiliones). Naturwissenschaften, 89, 357–360 (2002).
Duffield, R. M., Olubajo, O., Wheeler, J. W. & Shear, W. A. Alkylphenols in the defensive secretion of the nearctic opilionid. Stygnomma spinifera (Arachnida: Opiliones). J. Chem. Ecol. 7, 445–452 (1981).
Eisner, T., Rossini, C., González, A. & Eisner, M. Chemical defense of an opilionid (Acanthopachylus aculeatus). J. Exp. Biol. 207, 1313–1321 (2004).
Machado, G., Carrera, P. C., Pomini, A. M. & Marsaioli, A. J. Chemical defense in harvestmen (Arachnida, Opiliones): do benzoquinone secretions deter invertebrate and vertebrate predators? J. Chem. Ecol. 31, 2519–2539 (2005).
Hara, M. R., Cavalheiro, A. J., Gnaspini, P. & Santos, D. Y. A. C. A comparative analysis of the chemical nature of defensive secretions of Gonyleptidae (Arachnida: Opiliones: Laniatores). Biochem. Syst. Ecol. 33, 1210–1225 (2005).
Rocha, D. F. O., Hamilton, K., Gonçalves, C. C. S., Machado, G. & Marsaioli, A. J. 6-Alkyl-3,4-dihydro-2H-pyrans: chemical secretion compounds in neotropical harvestmen. J. Nat. Prod. 74, 658–663 (2011).
Caetano, D. S. & Machado, G. The ecological tale of Gonyleptidae (Arachnida, Opiliones) evolution: phylogeny of a Neotropical lineage of armoured harvestmen using ecological, behavioural and chemical characters. Cladistics 10.1111/cla.12009 (2013).
Schneider, B. Nuclear magnetic resonance spectroscopy in biosynthetic studies. Prog. Nucl. Magn. Reson. Spectrosc. 51, 155–198 (2007).
Maier, W., Shneider, B. & Strack, D. Biosynthesis of sesquiterpenoid cyclohexenone derivatives in mycorrhizal barley roots proceeds via glyceraldehyde 3-phosphate/pyruvate pathway. Tetrahedron Lett. 39, 521–524 (1998).
Pankewitz, F. & Hilker, M. Polyketides in insects: ecological role of these widespread chemicals and evolutionary aspects of their biogenesis. Biol. Rev. 83, 209–226 (2008).
Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. 48, 4688–4716 (2009).
Jarvis, A. P., Liebig, J., Hölldobler, B. & Oldham, N. J. Biosynthesis of the insect pheromone (S)-4-methyl-3-heptanone. Chem. Commun. 1196–1197 (2004).
Schulz, S., Fuhlendorff, J., Steidle, J. L. M., Collatz, J. & Franz, J. T. Identification and biosynthesis of an aggregation pheromone of the storage mite Chortoglyphus arcuatus. ChemBioChem 5, 1500–1507 (2004).
Amagai, K., Kudo, F. & Eguchi, T. Biosynthetic pathway of macrolactam polyketide antibiotic cremimycin. Tetrahedron 67, 8559–8563 (2011).
Halarnkar, P. P. & Blomquist, G. J. Comparative aspects of propionate metabolism. Comp. Biochem. Physiol. 92B, 227–231 (1989).
Blomquist, G. J., Halankar, P. P. & Dwyer, L. A. Propionate and methyl malonate metabolism in insects., in: ACS Symposium Series, Bioregulators for Pest Control, Vol. 276, 245–253 (ACS, Washington D.C. (1985).
Wakayama, E. D., Dillwith, J. W., Howard, R. W. & Blomquist, G. J. Vitamin B12 levels in selected insects. Insect Biochem. 14, 175–179 (1984).
Halarnkar, P. P., Chambers, J. D., Wakayama, E. J. & Blomquist, G. J. Vitamin B12 levels and propionate metabolism in selected non-insect arthropods and other invertebrates. Comp. Biochem. Physiol. 88B, 869–873 (1987).
Petroski, R. J., Bartelt, R. J. & Weisleder, D. Biosynthesis of (2E,4E,6E)-5-Ethyl-3-Methyl-2,4,6-nonatriene: the aggregation pheromone of Carpophilus freemani (Coleoptera: Nitidulidae). Insect Biochem. Mol. Biol. 24, 69–78 (1994).
Chan, Y. A., Podevels, A. M., Kevany, B. M. & Thomas, M. G. Biosynthesis of polyketide synthase extender units. Nat. Prod. Rep. 26, 90–114 (2009).
Erb, T. J. et al. Synthesis of C5-dicarboxylic acids from C2-units involving crotonyl-CoA carboxylase/reductase: The ethylmalonyl-CoA pathway. Proc. Natl. Acad. Sci. U. S. A. 104, 10631–10636 (2007).
Halarnkar, P. P., Nelson, J. H., Heisler, C. R. & Blomquist, G. J. Metabolism of propionate to acetate in the cockroach Periplaneta americana. Archiv. Biochem. Biophys. 236, 526–534 (1985).
Halarnkar, P. P., Chambers, J. D. & Blomquist, G. J. Metabolism of propionate to acetate in nine insect species. Comp. Biochem. Physiol. 84B, 469–472 (1986).
Crawford, J. M. & Clardy, J. Bacterial symbionts and natural products. Chem. Commun. 47, 7559–7566 (2011).
Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 26, 338–362 (2009).
Meinwald, J., Koch, K. F., Rogers, J. E., Jr & Eisner, T. Biosynthesis of arthropod secretions. III. Synthesis of simple p-benzoquinones in a beetle (Eleodes longicollis). J. Am. Chem. Soc. 88, 1590–1592 (1966).
Blum, M. S. Biosynthesis of arthropod exocrine compounds. Annu. Rev. Entomol. 32, 381–413 (1987).
Vavrecka von, M. & Hesse, M. Synthesis of monomorin I, a trail pheromone of the pharao ant (Monomorium pharaonis). Helv. Chim. Acta 74, 438–444 (1991).
Acknowledgements
The authors acknowledge Prof. C. H. Collins (Chemistry Institute/Unicamp) for revising the text and CNPq, Petrobrás and Fundação de Amparo à Pesquisa do Estado de São Paulo (GM 02/00381-0; 08/06604-7) for financial support.
Author information
Authors and Affiliations
Contributions
D.F.O.R., F.C.W. and A.J.M. are the chemists responsible for the experimental design and data interpretation. G.M. is the biologist responsible for the identification and collection of the individuals and furnished technical support to raise Iporangaia pustulosa individuals in the laboratory.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
suplementary infoormation
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Rocha, D., Wouters, F., Machado, G. et al. First Biosynthetic pathway of 1-hepten-3-one in Iporangaia pustulosa (Opiliones). Sci Rep 3, 3156 (2013). https://doi.org/10.1038/srep03156
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03156
This article is cited by
-
Chemical and evolutionary analysis of the scent gland secretions of two species of Gonyleptes Kirby, 1819 (Arachnida: Opiliones: Laniatores)
Chemoecology (2023)
-
On the enigmatic scent glands of dyspnoan harvestmen (Arachnida, Opiliones): first evidence for the production of volatile secretions
Chemoecology (2014)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
