
Abstract
In recent years, several invasive woodborers (Coleoptera: Cerambycidae) have been found in Japan. Aromia bungii is a worldwide important pest of fruits and ornamental species of the genus Prunus. It invaded Japan in the early 2010s and now causes heavy damage to stone fruit trees. Anoplophora glabripennis and Apriona swainsoni are destructive pests of street, ornamental and horticultural trees. The first step in intercepting these beetles is to detect their presence early in their infestation, as accurate identification is crucial for their management. Ejected frass is a major sign of infestation and likely holds information on the insect. We focused on chemicals in both larvae and frass, and conducted a GC–MS analysis of these three invasive beetles and the native Anoplophora malasiaca. In all four species, 4 or 5 species-specific hydrocarbons were detected in both larvae and frass. These results indicate that analysis of hydrocarbons in frass could allow definitive detection of invasive wood-boring pests.
Similar content being viewed by others

Introduction
Invasive beetle species, especially in the family Cerambycidae, are important pests worldwide1. Their larvae bore deeply into trees, shrubs and wood products. Invasive species can easily find new hosts and establish new habitats in invaded areas. In recent years, three invasive longhorn beetles species have been found in Japan, but there is not yet enough ecological information on them, such as life cycle and host range. And no effective control methods have also been established yet. The first important step in their eradication is to detect their presence within trees early in their infestation. Then, their accurate identification is crucial for their management, such as the selection of suitable pesticides, to prevent more damage.
Aromia bungii (Faldermann) (Coleoptera: Cerambycidae), the red-necked longhorn beetle, was first recognized in Japan in the early 2010s2, and is now (March 2023) present in 13 of the 47 prefectures3,4,5. It is an important wood-boring pest of fruits and ornamental species of the genus Prunus, including the culturally significant cherry blossom, which attracts tourists from both Japan and overseas. Horticultural species, especially peaches, are also heavily damaged by this species3.
Anoplophora glabripennis (Motschulsky) (Coleoptera: Cerambycidae), the Asian long-horned beetle, a devastating pest, was confirmed in Japan in 2020 and 20216,7,8,9,10,11,12 and is already present in North America and Europe13,14. It was included in a list of the 100 of the World’s Worst Invasive Alien Species15. It poses a serious threat to landscape trees, such as maple (Acer), poplar (Populus), and willow (Salix), often killing host trees by vigorous larval feeding16. In invaded regions, aggressive containment programs have been carried out, such as removal and destruction of all trees with signs of infestation16,17. Japan is home to a native Anoplophora beetle, A. malasiaca (Thomson), the white-spotted longicorn beetle18. It is synonymized with Anoplophora chinensis19, although its taxonomy is confusing. Here, we use ‘A. malasiaca’ refer to the Japanese population. It is a serious pest of horticultural tree, such as citrus (Citrus), apple (Malus) and pear (Pyrus), and of street trees, such as oriental planetree (Platanus) and willow18,20. It has a very wide range of host plants, 108 species in 73 genera21. The invasive and native Anoplophora beetles look similar and share some host plants (e.g. Salix, Cercidiphyllum), most people could not distinguish them. It can be tricky to distinguish them for control, so the early detection method is eagerly awaited.
Apriona swainsoni (Hope) (Coleoptera: Cerambycidae) was confirmed in Fukushima prefecture in 202122. It is a serious pest of Styphnolobium japonicum in China23. In Japan, it frequently attacks street trees including S. japonicum, but especially does Maackia amurensis.
Species identification of beetle larvae is difficult because of their morphological similarities. For determination, larvae are often taken out from the tree and reared in the laboratory until adulthood, which can take several months24. Moreover, the collection of larvae from trees can injure the host tree as well as the larvae25. On the other hand, frass ejection from trees infested by pest insects is often observed in the field. Frass includes woody materials and sometimes faeces of infesting insects.
Invasive beetles can be identified from their genetic information. Analysis of frass by real-time PCR confirmed the presence of A. bungii25 and another wood-boring pest26. Although this method guarantees accuracy of species identification, the stability of genetic information in the field is not assured under the tough conditions of rain and sunlight. To develop better methods for identifying infesting beetles under tough field conditions, we focused on the hydrocarbons in their frass. Among possible chemical components in frass, hydrocarbons are synthesized by insects and are chemically more stable than polar compounds and DNA. Insect cuticular hydrocarbons (CHCs) have roles in maintaining the insect’s water balance and acting as signalling molecules for mate recognition and chemical communication27,28,29. CHCs of many adult insects are reported to be species specific30. In contrast, only a few reports of larval CHCs were found: one on blowfly larvae31, one on Tribolium confusum (Coleoptera, Tenebrionidae)32, and some on parasitic lepidopteran larvae which penetrate ant nests33. These reports describe age-dependent changes in larval CHCs (blowfly), kairomonal activity (T. confusum), or chemical camouflage.
Here, we compared profiles of CHCs of larvae and frass of three invasive and one native beetles and confirmed these commonalities. Based on these comparisons, we could reveal that analysis of hydrocarbons in frass could contribute to detecting beetle species in trees. Furthermore, we demonstrate the validity of our method by using frass samples of A. bungii. We discuss the possible use of hydrocarbon analysis for the identification of infesting species.
Results
Common hydrocarbons detected in larvae and frass of four beetle species
Table 1 lists hydrocarbons commonly detected in frass and larvae of each species, with the Kovát’s Index and molecular or fragment ions used for their identification. Double-bond positions of unsaturated hydrocarbons were determined only in A. bungii. Some hydrocarbons were detected in multiple species; for example, n-tricosane was detected in A. bungii, A. glabripennis and A. swainsoni. All four species have species specific hydrocarbons: for example, 6,9-C25:2 and 6,9-C27:2 in A. bungii; C27:1 in A. glabripennis; C28, 4Me-C28 and C29 in A. malasiaca; and C23:1 and C25:1 in A. swainsoni.
Aromia bungii
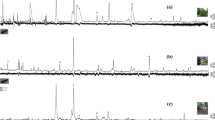
Both A. bungii larvae and frass showed the same five hydrocarbons (Table 1; Fig. 1, numbered peaks): three were saturated (C23, C24, C25) and two were unsaturated (diene: 6,9-C25:2, 6,9-C27:2). C23 and C25:2 were always detected at strong intensity.

Total ion chromatogram of GC/MS profiles of hydrocarbons of (a) larvae and (b) frass of A. bungii from plum tree. C23, n-tricosane; C24, n-tetracosane; 6,9-C25,2, 6,9-pentacosadiene; C25, n-pentacosane; 6,9-C27,2; 6,9-heptacosadiene.
Anoplophora glabripennis
Both A. glabripennis larvae and frass showed the same five hydrocarbons (Table 1; Fig. 2, numbered peaks): four were saturated (C23, C24, C25, C27) and one was unsaturated (monoene: C27:1). C23 and C25 were always detected at strong intensity.

Total ion chromatogram of GC/MS profiles of hydrocarbons of (a) larvae and (b) frass of A. glabripennis from willow tree. C23, n-tricosane; C24, n-tetracosane; C25, n-pentacosane; C27,1, heptacosene; C27, n-heptacosane.
Anoplophora malasiaca
Both A. malasiaca larvae and frass showed the same four hydrocarbons (Table 1; Fig. 3, numbered peaks): three were saturated (C27, C28, C29) and one was a monomethyl (4Me-C28). 4Me-C28 and C29 were always detected at strong intensity.

Total ion chromatogram of GC/MS profiles of hydrocarbons of (a) larvae and (b) frass of A. malasiaca from willow tree. C27, n-heptacosane; C28, n-octacosane; 4Me-C28, 4-methyloctacosane; C29, n-nonacosane.
Apriona swainsoni
Both A. swainsoni larvae and frass showed the same five hydrocarbons (Table 1; Fig. 4, numbered peaks): three were saturated (C23, C25, C27) and two were unsaturated (monoene: C23:1, C25:1). C25 and C25:1 were always detected at strong intensity.

Total ion chromatogram of GC/MS profiles of hydrocarbons of (a) larvae and (b) frass of A. swainsoni from M. amurensis. C23,1, tricosene; C23, n-tricosane; C25,1, pentacosene; C25, n-pentacosane; C27, n-heptacosane.
Comparison of detected hydrocarbons among 4 different beetle species
Every beetle’s larva and frass had some hydrocarbons (Table 1). Table 2 summarizes the hydrocarbons detected in each species.
Hydrocarbon analysis of field-collected frass of Aromia bungii from two tree species
The field-collected frass of A. bungii larvae (over 2-month-old larvae) showed the same profile as that under laboratory conditions (Figs. 1a, 5a,b). The same five hydrocarbons were detected from the frass of both tree species (Fig. 5a, cherry tree; b, peach tree, numbered peaks).

Total ion chromatogram of GC/MS profiles of hydrocarbons of A. bungii frass collected from (a) cherry tree in the field and (b) peach tree in the laboratory. Peak numbers indicate the same hydrocarbon components shown in Fig. 1.
Hydrocarbon analysis of sawdust and frass from plum tree infested by A. bungii
Larval-specific hydrocarbons were not detected in the sawdust extract (Fig. 6a). Only frass showed the five characteristic hydrocarbons (Fig. 6b, arrows).

Total ion chromatogram of GC/MS profiles of hydrocarbons of (a) sawdust and (b) frass made by A. bungii. All samples were collected from plum trees. Peak numbers indicate the same hydrocarbon components shown in Fig. 1.
Time-course hydrocarbon analysis of Aromia bungii frass
Weekly analysis of A. bungii frass showed that the hydrocarbon profiles were unchanged over 5 weeks after larvae hatching (Fig. 7a, week 1; b, week 3; c, week 5). As a reference data, field-collected frass ejected by over-wintered, 9-month larvae included same species-specific hydrocarbons (Fig. 7d). Aromia bungii-specific hydrocarbon profiles were detected through the larval period.

Total ion chromatograms of GC/MS profiles of hydrocarbons of A. bungii frass ejected by larvae of different age. Each chromatogram shows the frass hydrocarbon profiles of larvae which after hatching (a) week 1; (b) week 3; (c) week 5; (d) around week 36 (supposed to overwintered, 9-month-old larvae). Peak numbers indicate the same hydrocarbon components shown in Fig. 1.
Discussion
We developed a new method to detect three invasive and one native beetle species from frass ejected from infested trees. Extraction of hydrocarbons and GC–MS analysis correctly identified the beetle species in the trees. Every sample showed clear total ion chromatograms, and all detected peaks could be chemically characterized. Four or five hydrocarbons common to both larvae and frass were consistently detected within species. As reported of other insect species28, CHC profiles of tested larvae differed among species. We also got the same hydrocarbon profiles of A. bungii frass from laboratory (Fig. 1) and field samples (Fig. 5). This result reveals that our method could be applied to both laboratory and field samples.
CHCs of insects consist of a complex mixture of straight-chain (saturated), unsaturated, and methyl-branched components with over 20 carbon atoms28. In A. bungii, A. malasiaca and A. glabripennis, chemical profiles of CHCs of adults and their larvae are not common [34 (cf. our data), 35,36]. There is no information on the roles of larval CHCs of these beetles in chemical communication, but some may have kairomonal activity against parasitoids. Dastarcus helophoroides (Coleoptera: Bothrideridae) is a larval predator of cerambycid forest tree pests, and its mass release is one option for controlling beetles37. Volatile chemicals from the host tree or insects play a key role in host searching from a distance37, although, commonly detected CHCs in this study are slightly volatile and almost non-volatile, there is a possibility to have close-distant or contact kairomonal activity.
Currently, the most accurate method to identify infesting beetle species is to collect larvae and rear them to adulthood, because beetle larvae look very similar38. However, this method damages trees. Instead, if ejected frass is present, hydrocarbon analysis could be used to identify them without further damage to the tree. We confirmed the validity of our method with A. bungii because of sample availability. The results of the chemical analysis of A. bungii frass from plum and peach (Figs. 1b, 5b), plum (Fig. 6b), and cherry trees (Fig. 5a) show that our method can work with different tree species. Even within their ‘first week of feeding or just after hatch, frass was seen. Only 1 mg equivalent of frass extract injected into the GC/MS could determine the profiles of its typical hydrocarbons. Thus, our method could be applied very early in larval infestation; the earlier an infestation can be detected, the earlier it can be treated, avoiding further damage. This is crucial to reducing the damage by wood-boring beetles to economically, culturally or historically important trees. Throughout our experiments, larvae-frass common hydrocarbons were stably detected from early stage (1–5 weeks, Fig. 7a–c), 2-month over (Fig. 5a,b) and 9-month over (Fig. 7d). As is known in other insects, CHCs can change over the larval stage31. If the same tendency existed in the beetles, our method could allow us to estimate larval age or size. The entire larval stage changes of their CHCs composition should be investigated in our future work.
We could detect hydrocarbons in almost all A. bungii and A. swainsoni frass samples. In the case of A. glabripennis and A. malasiaca, however, the frass sometimes did not include detectable amounts of hydrocarbons. In our interpretation, the difference may be caused by larval behaviour. Aromia bungii and species of Apriona (including A. swainsoni) continually make excretory holes for ejecting their frass39,40. During our preliminary experiment, Aromia larvae used their body segments to eject the frass. This active ejection of frass may transfer their CHCs to the frass. On the other hand, this behaviour has not been reported in Anoplophora larvae and we haven’t observed it. We could detect A. malasiaca-specific hydrocarbons in frass from willow twigs (Fig. 3), but not in other trials using fresh or live trees. One possibility is that when feeding on spatially restricted dry material (such as cut twigs), A. malasiaca larvae have to maintain their living and feeding space by ejecting frass. As a result of that behaviour, their CHCs might often be undetectable.
Many other sympatric insects eject frass in the same way as our beetles. To improve risk management of trees worldwide, we should accumulate examples of insect cuticular and frass hydrocarbon profiles not only of Coleopteran but also of other frass-ejecting insect species.
Materials and methods
Collection and rearing of adult beetles
Aromia bungii
Adult A. bungii were collected by hand from peach groves in Sano city, Tochigi Prefecture, Japan, in late June 2022.
Anoplophora glabripennis and A. malasiaca
Adult A. glabripennis and A. malasiaca were collected by hand from street trees such as Salix spp. (willow) and Cercidiphyllum japonicum in Tsukuba, Omitama and Ishioka cities, Ibaraki Prefecture, in June and July 2022.
Apriona swainsoni
Adult A. swainsoni were collected by hand from Maackia amurensis in Tenei village and Koriyama city, Fukushima Prefecture, in August 2022. Some emerged from host plants (M. amurensis) cut back in Koriyama city in May 2022.
Aromia bungii, A. glabripennis and A. malasiaca beetles were individually reared in clear plastic cups (ca. 11 cm diam. × 9.5 cm height) at 24 °C under a 15L:9D photoperiod, illuminated by fluorescent lamps. All females mated at least once in our laboratory. A. bungii were fed artificial diet for rhinoceros beetle (Fujikon Co. Ltd, Osaka, Japan); and A. glabripennis and A. malasiaca were fed willow and citrus twigs, renewed every 5 days. Apriona swainsoni were reared in meshed cages (22 cm × 42 cm × 42 cm), together with two or three branches (30 cm long, 10 cm diameter) of M. amurensis for feeding and oviposition, renewed every 5 days.
Frass collection
Aromia bungii
In the laboratory, small branches of plum or peach (5 cm diam., 10 cm long) were partially wrapped in 1-cm-wide Parafilm (Bemis Flexible Packaging, Chicago, IL, USA), and females of A. bungii were housed on them for 2 days. Eggs laid between the bark and the Parafilm were collected and laid on wet filter paper (9 cm diam., Toyo Roshi Kaisha, Ltd, Tokyo, Japan). Eggs on filter paper were placed together in a plastic Petri dish (9 cm diam. × 2 cm height) at 24 °C under a 15L:9D photoperiod, illuminated by fluorescent lamps for 10 days. Newly hatched larvae were placed on new twigs of plum. After a few weeks, larvae began to eject frass, which was then collected by forceps into glass vials weekly for 5 weeks. Frass was also collected from peach groves and cherry trees in Sano and Ashikaga city on October 4th, 2022, Tochigi prefecture, where we collected the A. bungii. These field collected frass were expected to be ejected by over 2-month larvae of A. bungii. Frass of over-wintered larvae also collected on May 25th, 2022 from woods bring back from peach groves in Sano city in December 2021 and maintained in the laboratory (9-month over larvae).
Anoplophora glabripennis and A. malasiaca
Females of each species were individually kept in clear plastic cups (ca. 11 cm diam. × 9.5 cm height) together with host twigs, mainly willow (3 cm diam., 5 cm long). Eggs laid under the bark were collected and laid on wet filter paper. Eggs on filter paper were placed together in a plastic Petri dish as above for 10 days. Newly hatched larvae were placed on new plum or willow twigs. Ejected frass was collected as above in our laboratory (A. glabripennis and A. malasiaca, three samples) and from two frass samples were collected from Cercidiphyllum japonicum in Tsukuba from the field (A. glabripennis).
Apriona swainsoni
Males and females of A. swainsoni were kept together in small mesh cages (28 cm × 30 cm × 18 cm). Females laid eggs into Maackia Amurensis branches. The branches were maintained as above. After a few weeks, larvae began to eject frass, which was collected as above in our laboratory (two samples) in 2022. In 2023, three larvae and frass were collected from the infested wood cut in Fukushima prefecture.
Extraction and purification of hydrocarbons from frass and larvae
All frass was freeze-dried for 16 h at − 30 °C before extraction. Dried samples were weighed and then extracted in 5–8 mL of n-hexane (HPLC grade, Fujifilm-Wako, Osaka, Japan) for 5 min. Sample sizes of each frass were 10 for A. bungii, 5 for A. glabripennis and A. swainsoni, 3 for A. malasiaca.
Larvae were placed on filter paper for 20 min to void faeces and then frozen (− 30 °C). Each larva was put into a glass vial (5–12 mL) and extracted with n-hexane for 5 min. Sample sizes of each larva were 5 for A. bungii, 5 for A. glabripennis, 3 for A. malasiaca, and 5 for A. swainsoni.
Hexane extracts were filtered and poured through a silica gel column (Wakogel C-300, 0.5 g, Fujifilm-Wako). The final volume of extracts after concentration depended on frass volume or larval size, but 0.001 g equivalent of frass sample or 0.01 larva equivalent dissolved in 1 µL n-hexane was injected into the GC–MS.
Comparison of hydrocarbons in frass and larvae on plum tree in the laboratory
Females of A. bungii were introduced into a meshed cage with a plum tree in a pot (2 years old, 100 cm high) and allowed to lay freely for 2 days. After 2–3 weeks, hatched larvae started to eject frass. At the same time, sawdust was collected by hand-sawing for analysis to confirm the components of the host plant. Frass and sawdust were extracted as above.
Gas chromatography/mass spectrometry (GC/MS) analyses
GC/MS analyses were performed on an Agilent 7890N GC system (Agilent, Santa Clara, CA, USA) interfaced to a JEOL JMS-T100GC time-of-flight mass spectrometer (JEOL, Tokyo, Japan) in EI mode at 70 eV at 220 °C. Samples were injected in splitless mode at 220 °C for 1 min, with helium as the carrier gas in constant flow mode (1.1 mL/min). The capillary column, a DB-5MS (30 m × 0.25 mm ID × 0.25 μm film thickness; Agilent), was linked to an MS, and the GC oven temperature was held at 40 °C for 1 min, increased from 40 to 250 °C at 10 °C/min and then held at the final temperature for 10 min.
Identification of hydrocarbons in larvae and frass
Hydrocarbons were identified by the mass spectrum and the Kovát index of each peak41. A reference hydrocarbon mixture that included saturated, even-numbered hydrocarbons from C12 to C32 was analysed as above, with reference to KIs of standard compounds. Only A. bungii yielded diene components in sufficient amount for derivatization to decide the position of two double bonds. Diene components were partially reduced with hydrazine and epoxidated by m-chloroperoxybenzoic acid. The resultant epoxide mixture was analysed by GC–MS, and the position of the double bonds was determined42.
Data availability
The data presented in this study are available on request from the corresponding author.
References
Brockerhoff, E. G., Jones, D. C., Kimberley, M. O., Suckling, D. M. & Donaldson, T. Nationwide survey for invasive wood-boring and bark beetles (Coleoptera) using traps baited with pheromones and kairomones. For. Ecol. Manag. 228, 234–240 (2006).
Kano, M., Nonaka, T., Kiriyama, S. & Iwata, R. Aromia bungii (Coleoptera: Cerambycidae), an invasive cerambycid, found at Soka, Saitama Pref., Japan, infesting cherry trees, Cerasus ×yedoensis ‘Somei-yoshino’. For. Pests. 63, 101–105 (2014) (in Japanese).
Shoda-Kagaya, E. Red necked longhorn beetle Aromia bungii damaged areas in Japan and Rosaceae tree protection methods: Tips for controlling. Plant Prot. 74(5), 293–296 (2020) (in Japanese).
Kanagawa prefecture. First recorded red necked longhorn beetle Aromia bungii investation in Kanagawa prefecture. https://www.pref.kanagawa.jp/documents/77637/kubiaka-nougi.pdf (2021). (in Japanese).
Hyogo prefecture. First recorded red necked longhorn beetle Aromia bungii investation in Hyogo prefecture. https://web.pref.hyogo.lg.jp/press/20220629_10646.html (2022). (in Japanese).
Kanada, Y. & Kidono, H. The Asian longhorned beetle of Aichi prefecture. Gekkan-Mushi 608, 53–54 (2021) (in Japanese).
Nishiura, K., Shinozaki, S., Suzuki, H. & Fuda, S. Collection of the Asian longhorned beetle in Tsukuba City, Ibaraki prefecture. Gekkan-Mushi 608, 53 (2021) (in Japanese).
Saitama Prefecture. Beware of the Asian longhorned beetle. https://www.pref.saitama.lg.jp/a0907/tuyahadakamikiri.html (accessed 12 Feb 2022) (in Japanese).
Sato, H. & Nishiura, K. Collection of the Asian longhorned beetle in Shirakawa City, Fukushima Prefecture. Gekkan-Mushi 608, 54 (2021) (in Japanese).
Yanagi, T., Nagahata, Y., Yoshino, K. & Akita, K. A new record of Anoplophora glabripennis (Motschulsky, 1853) (Coleoptera, Cerambycidae), with newly host plants recorded Cercidiphyllum japonica and Cerasus ×yedoensis from Miyagi pref, Japan. Gekkan-Mushi 609, 23–27 (2021) (in Japanese).
Iwata, T. & Kiriyama, S. First records of Anoplophora glabripennis (Motschulsky, 1853) in Toyama prefecture, including a collected record of 2010. Gekkan-Mushi 611, 34–36 (2022) (in Japanese).
Shimono, M., Nagano, A. & Hirayama, H. A new record of Anoplophora glabripennis (Motschulsky, 1853) in Yamaguchi prefecture. Gekkan-Mushi 613, 56 (2022) (in Japanese).
European and Mediterranean Plant Protection Organization (EPPO). EPPO A2 List of pests recommended for regulation as quarantine pests-version 2022-09. https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed 21 Dec 2022).
Animal and Plant Health Inspection Service (APHIS). Asian Longhorned Beetle—Quarantines. https://www.aphis.usda.gov/aphis/resources/pests-diseases/asian-longhorned-beetle (accessed 21 Dec 2022).
Lowe, S., Browne, M., Boudjelas, S., de Poorter, M. 100 of the World’s Worst Invasive Alien Species. A selection from the Global Invasive Species Database. Published by The Invasive Species Specialist Group (ISSG) a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN). 2000. Updated and reprinted version: November 2004.
Haack, R., Herard, F., Sun, J. & Turgeon, J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 55, 521–546 (2010).
Hérard, F. et al. Anoplophora spp. in Europe: Infestations and management process. Bull. OEPP/EPPO Bull. 36, 470–474 (2006).
Ohbayashi, N. Genus Anoplophora Hope, 1839. In An Illustrated Guide to Identification of Longicorn Beetles of Japan (eds Ohbayashi, N. et al.) 583–584 (Tokai University Press, 1992) (in Japanese).
Lingafelter, S. W. & Hoebeke, E. R. Revision of Anoplophora (Coleoptera: Cerambycidae) 236 (Entomological Society of Washington, 2002).
Kojima, K., Nakamura, S. Food Plants of Cerambycid Beetles (Cerambydae, Coleoptera) in Japan 128–133 (Hiba Society of Natural History Press, 1986) (in Japanese).
Sjöman, H., Östberg, J. & Nilsson, J. Review of host trees for wood-boring pests Anoplophora glabripennis and Anoplophora chinensis: An urban forest perspective. Arboricult. Urban For. 40, 143–164 (2014).
Muto, S., Yosii, H. & Tsutsumi, T. Preliminary report on distribution, infestation, and feeding plants of the alian longhorn beetle Apriona swainsoni swainsoni (Hope, 1840) (Coleoptera: Cerambycidae, Lamiinae) discovered from Fukushima Prefecture, Japan. Jp. J. Entomol. (New Series) 25(1), 18–24 (2022) (in Japanese).
Liu, H. et al. Pest risk assessment of Dendroctonus valens, Hyphantria cunea and Apriona swainsoni in Beijing. Front. For. China 3, 328–335 (2006).
Kitajima, H. Identification and rearing of beetles infesting trees. Tree For. Health Chiba Branch Annu. Rep. 9, 20–26 (2018) (in Japanese).
Rizzo, R. et al. Identification of the red-necked longhorn beetle Aromia bungii (Faldermann, 1832) (Coleoptera: Cerambycidae) with real-time PCR on frass. Sustainability 12, 6041 (2020).
Rizzo, R. et al. TaqMan probe assays on different biological samples for the identification of three ambrosia beetle species, Xylosandrus compactus (Eichoff), X. crassiusculus (Motschulsky) and X. germanus (Blandford) (coleoptera Curculionidae Scolytinae). 3 Biotech 11, 259 (2021).
Chung, H. & Carroll, S. B. Wax, sex and the origin of species: Dual roles of insect cuticular hydrocarbons in adaptation and mating. BioEssays 37, 822–830 (2015).
Blomquist, G. J., Bagnères, A-G. Chemistry, biochemistry, and physiology. In Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology. 3–206 (Cambridge University Press, 2010).
Ginzel, M. D., Blomquist, G. J. Insect hydrocarbons: Biochemistry and chemical ecology. In Extracellular matrices in arthropods (eds. Cohen, E., Moussian, B). 221–252 (Springer, 2016).
Blomquist, G. J., Tittiger, C., Jurenka, R. Cuticular hydrocarbons and pheromones of arthropods. In Hydrocarbons, Oils and Lipids: Diversity, Origin, Chemistry and Fate, 213–244 (Springer, 2020).
Zhu, G. H., Ye, G. Y., Hu, C., Xu, X. H. & Li, K. Development changes of cuticular hydrocarbons in Chrysomya rufifacies larvae: Potential for determining larval age. Med. Vet. Entomol. 20, 438–444 (2006).
Awater-Salendo, S., Voigt, D., Hilker, M. & Fürstenau, B. Cuticular hydrocarbon trails released by host larvae lose their kairomonal activity for parasitoids by solidification. J. Chem. Ecol. 47, 998–1013 (2021).
Akino, T., Knapp, J. J., Thomas, J. A. & Elms, G. W. Chemical mimicry and host specificity in the butterfly Maculinea rebeli, a social parasite of Myrmica and colonies. Proc. R. Soc. Lond. B 266, 1419–1426 (1999).
Di Palma, A., Pistillo, M., Griffo, R. Garonna, A. P., Germinara, G. S. Scanning electron microscopy of the antennal sensilla and their secretion analysis in adults of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae). Insects 10, 88 (2019).
Zhang, A. et al. Evidence for contact sex recognition pheromone of the Asian longhorned beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae). Naturwissenschaften 90, 410–413 (2003).
Fukaya, M. et al. Hydrocarbon components in contact sex pheromone of the white-spotted longicorn beetle, Anoplophora malasiaca (Thomson) (Coleoptera: Cerambycidae) and pheromonal activity of synthetic hydrocarbons. Entomol. Sci. 3, 211–218 (2000).
Li, L. R., Balakrishnan, K., You, Q. L. & Schütz, S. EAG response and behavioral orientation of Dastarcus helophoroides (Fairmaire) (Coleoptera: Bothrideridae) to synthetic host-associated volatiles. PLoS ONE 12, e0190067 (2017).
Haruyama, N. et al. Peach orchard damages caused by the invasive red-necked longhorn beetle Aromia bungii (Faldermann) in Tochigi Prefecture. Ann. Rep. Kanto Pl. Prot. Soc. 66, 106–109 (2019) (in Japanese).
Shoda-Kagaya, E. Invasion of the red-necked longicorn beetle, Aromia bungii: Damages of Rosaceae trees and practical control methods. Tree For. Health 22, 68–72 (2018) (in Japanese).
EPPO. Pest Risk Analysis for Apriona germari, A. japonica, A. cinerea (EPPO, 2013).
Kovát, E. Gas chromatographic characterization of organic substances in the retention index system. Adv. Chromatogr. 1, 229–247 (1965).
Yamaoka, R., Fukami, H. & Ishii, S. Isolation and identification of the female sex pheromone of the potato tuberworm moth, Phthorimaea operculella (Zeller). Agric. Biol. Chem. 40, 1971–1977 (1976).
Acknowledgements
This research was partially supported by a grant (JPJ007097) from the Project of the Bio-oriented Technology Research Advancement Institution (Research program on development of innovative technology 04015C1). We thank Nami Uechi of NARO and Shinsuke Sato of the Horticultural Research Institute, Ibaraki Agricultural Center, for collecting frass samples, Anoplophora beetles and host trees; Etsuko Kagaya of the Forest Research and Management Organization for providing host trees; Yukari Anzai of the Japan Tree Doctors Association for providing A. swainsoni and a log of infested Maackia amurensis; and Naoto Haruyama of Tochigi Prefectural Agricultural Experiment Station for providing logs of damaged peach trees from peach orchards in Sano city. We also thank Ms. Kazue Murata for her assistance with taking care of the beetles. N. F.-T. also thank the late Ryohei Yamaoka, professor emeritus of Kyoto Institute of Technology, for the inspiration to start this study.
Author information
Authors and Affiliations
Contributions
N.F.-T. contributed to the study design and carried out the analyses and characterization. N.F.-T. and H.Y. contributed to sample collection, discussed the results and wrote the manuscript. Both reviewed and approved the final version with consent for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.








Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fujiwara-Tsujii, N., Yasui, H. Detection of invasive and native beetle species within trees by chemical analysis of frass. Sci Rep 13, 11837 (2023). https://doi.org/10.1038/s41598-023-38835-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-38835-x
This article is cited by
-
Biology, impact, management and potential distribution of Aromia bungii, a major threat to fruit crops around the world
Journal of Pest Science (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}