
Abstract
The immunodominant staphylococcal antigen A (IsaA) is a potential target for active or passive immunization against the important human pathogen Staphylococcus aureus. Consistent with this view, monoclonal antibodies against IsaA were previously shown to be protective against S. aureus infections in mouse models. Further, patients with the genetic blistering disease epidermolysis bullosa (EB) displayed high IsaA-specific IgG levels that could potentially be protective. Yet, mice actively immunized with IsaA were not protected against S. aureus infection. The present study was aimed at explaining these differences in IsaA-specific immune responses. By epitope mapping, we show that the protective human monoclonal antibody (humAb) 1D9 recognizes a conserved 62-residue N-terminal domain of IsaA. The same region of IsaA is recognized by IgGs in EB patient sera. Further, we show by immunofluorescence microscopy that this N-terminal IsaA domain is exposed on the S. aureus cell surface. In contrast to the humAb 1D9 and IgGs from EB patients, the non-protective IgGs from mice immunized with IsaA were shown to predominantly bind the C-terminal domain of IsaA. Altogether, these observations focus attention on the N-terminal region of IsaA as a potential target for future immunization against S. aureus.
Similar content being viewed by others
Introduction
Staphylococcus aureus can cause a wide variety of diseases and has a strong tendency of developing resistance against multiple antibiotics1. Methicillin-Resistant S. aureus (MRSA)- associated infections are becoming increasingly harder to treat. Therefore, a renewed focus on the development of alternative means of treatment has arisen. Whereas many infectious diseases are nowadays controlled through vaccination, S. aureus immunity has proven hard to achieve with vaccines2,3,4,5. As an alternative to the active immunization against S. aureus with vaccines, passive immunization with monoclonal antibodies specifically targeting S. aureus is currently explored6,7,8,9,10,11.
Invariantly expressed cell surface-exposed proteins are attractive potential targets for immunization, due to their high accessibility to the human immune system12. This focused attention on the immunodominant S. aureus antigen A (IsaA). The IsaA protein was first described in the year 2000 as an antigen recognized by IgGs from patients with sepsis caused by MRSA13. Subsequent, proteomic analyses of the S. aureus exoproteome revealed that the IsaA protein was invariantly produced by all investigated isolates of this pathogen14, 15. More specifically, IsaA is a non-covalently cell wall attached protein that is both exposed to the cell surface and secreted13, 16, 17. The IsaA protein has a putative soluble lytic transglycosylase domain at the C-terminus, indicating a role in peptidoglycan turnover and cell wall hydrolysis17, 18. Interestingly, this C-terminal active site domain is exposed to the staphylococcal cell surface while the precise localization of the N-terminal domain was so far not known12, 18. Further, IsaA was localized to the septal region of dividing cells, suggesting its involvement in cell growth, separation and survival16. In addition to its potential role in cell division, IsaA was shown to be involved in biofilm formation17, 19, 20.
The immunodominant nature of IsaA was confirmed by different studies showing that both IgG1 and IgG4 against this protein are present in sera from a wide range of healthy human individuals and patients13, 21,22,23,24. Interestingly, healthy S. aureus carriers showed significantly higher IgG levels against IsaA than non-carriers22. Furthermore, patients with the genetic blistering disease epidermolysis bullosa (EB), who are highly colonized with S. aureus, displayed very high IgG responses against IsaA23. Notably, EB patients do not frequently suffer from S. aureus bacteremia, even though their skin barrier provides only limited protection against S. aureus. Although not conclusive, these observations did suggest that the anti-IsaA antibodies and other anti-staphylococcal antibodies could perhaps be involved in the protection of EB patients against serious S. aureus infections. Furthermore, the high immunogenicity of IsaA and its ubiquitous presence in S. aureus indicated that IsaA could be a suitable target for passive immunization. This view was supported by passive immunization experiments in animal studies. Firstly, a murine monoclonal antibody (mAb) targeting IsaA was shown to be protective in mouse models of catheter-related S. aureus infection7, and a humanized form of this mAb mediated bacterial killing in blood samples from high risk patients8. In another study, we showed that a fully human monoclonal antibody (humAb) against IsaA (designated 1D9) improved the survival of mice challenged with a clinical S. aureus isolate6.
Intriguingly, several active immunization studies in animals also revealed high immune responses against IsaA25,26,27,28. Nonetheless, no protection against S. aureus infection was observed in these immunized animals. This raised the question why certain monoclonal antibodies against IsaA, such as 1D9, are at least partially protective against S. aureus infection, while high anti-IsaA IgG levels raised by animal immunization were not protective. The present study was aimed at explaining this difference by assessing the IsaA epitopes recognized by the apparently protective and non-protective antibodies. Briefly, the results show that potentially protective antibodies recognize a different epitope in IsaA than the non-protective antibodies.
Materials and Methods
Bioinformatics
BLAST searches were performed with default search parameters (Program BLASTP 2.3.0+, refs 29 and 30; word size 6; expect value 10; hitlist size 1000; gapcosts 11,1; matrix BLOSUM62; filter string F; genetic code 1; window size 40; threshold 21; composition-based stats 2) against the NCBI Protein Reference Sequences Database (refseq_protein; posted date: December 7, 2015 10:23 AM). Pfam sequence searches (http://pfam.xfam.org/search) were performed with default options: use E-value 1.0 cut-off31. Multiple sequence alignments were performed using COBALT32 with default parameters (alignment parameters: gap penalties -11, -1; end-gap penalties -5, -1; CDD Parameters: Use RPS BLAST on; Blast E-value 0.003; find conserved columns and recompute on; query clustering parameters: use query clusters on; word size 4; max cluster distance 0.8; alphabet regular). Visualizations of HMM logos (representing both sequence alignments and profile hidden Markov models) were obtained using Skylign (http://skylign.org)33 with the following parameters: alphabet: AA; letter height: information content above background; alignment processing: HMM - remove mostly-empty columns. Predicted protein masses (kDa) and isoelectric points (pI) were obtained using the Expasy compute pI/Mw tool (web.expasy.org/compute_pi/). Data from Western blots were quantified by ImageJ software (available via http://rsbweb.nih.gov/ij/).
Bacterial strains and growth conditions
Bacterial strains and plasmids that were used in this study are listed in Table 1. Staphylococcal strains were grown at 37 °C, 250 rpm (0.87 × g) in Tryptone Soy Broth (TSB; Oxoid, Hampshire, UK). L. lactis strains were grown at 30 °C in M17 broth (Oxoid Limited) or on M17 plates with 1.5% agar and 0.5% glucose (w/v), supplemented with chloramphenicol (5 µg/ml) for plasmid selection. For in vivo studies the S. aureus clinical isolate P15 was grown in Brain Heart Infusion broth (BHI; Becton Dickinson, Breda, The Netherlands).
Construction of expression plasmids for the production of IsaA and its derivatives
PCR primers for the construction of IsaA protein-expressing plasmids are shown in Supplementary Table 1. DNA was isolated with the Genelute bacterial genomic DNA kit (Sigma-Aldrich, Zwijndrecht, The Netherlands). PCR was performed with the Phusion Hot Start II polymerase (Thermo Fisher Scientific, Wilmington, Delaware USA) using genomic DNA of S. aureus NCTC8325 as a template as described before34. The PCR fragments purified using the High Pure PCR purification kit (Analytic Jena, Jena, Germany) were cleft with BamHI and NotI restriction enzymes (New England Biolabs, Ipswich, USA) and ligated to BamHI/NotI-linearized vector DNA using T4 DNA Ligase (New England Biolabs). Of note, PCR products obtained with primer combinations including a reverse primer with a stop codon (F1/R2) were inserted into plasmids pNG4110, and PCR products obtained with reverse primers lacking a stop codon (F1/R1) were inserted into plasmid pNG4210. The resulting plasmids were transferred to electrocompetent L. lactis PA1001 as described before35. All plasmids were verified by sequencing (Eurofins MWG Operon, Ebersberg, Germany).
HumAb production and labeling
The humAb 1D9 directed against S. aureus IsaA was produced as described before6. Briefly, Expi293 cells were transiently transfected with plasmids encoding the H and K fragments of 1D9 (Expi293 Expression System, Life Technologies). Secreted 1D9 antibodies were isolated from the cell culture medium by Protein A column purification (HiTrap Protein A HP, GE Lifesciences) followed by desalting (HiTrap Desalting, GE Lifesciences) according to the manufacturer’s protocol. 1D9 F(ab’)2 nanobodies were derived from the complete 1D9 humAb using the FragIT kit (Genovis, Sweden) following the manufacturer’s protocol. The complete 1D9 antibody or the respective 1D9 F(ab’)2 nanobodies were labeled with IRDye 800CW (LI-COR Biosciences, Bad Homburg, Germany) by incubation with 20 µg of this dye per mg of protein in PBS (pH 8.5, 2 h, room temperature). The labeled antibodies were isolated and desalted as described above, and they were stored in the dark at 4 °C.
Protein expression, purification and detection of IsaA and its subdomains
The production and isolation of IsaA-His6 from E. coli BL21DE3 (pET24d::isaA::his 6) was performed as described previously6. Expression of IsaA and its derivatives in L. lactis PA1001 was induced with nisin (3 ng/ml, Sigma-Aldrich, St. Luis, MO) at an Optical Density at 600 nm (OD600) of ~0.5. After continued overnight incubation at 30 °C, cells were separated from the growth medium by centrifugation. Protein samples from both fractions were prepared as described before34. All proteins obtained were analyzed by Lithium Dodecyl Sulphate (LDS)-PAGE (NuPAGE gels, Life Technologies), and visualized either by protein staining (Simply BlueTM Safe Staining, Life Technologies) or by Western blotting on nitrocellulose membranes (Protan nitrocellulose transfer paper, Whatman, Germany). Immunodetection of particular proteins was achieved with the following antibodies or sera: mouse anti-His tag (Life Technologies), rabbit anti-IsaA (kindly provided by N. Sakata)16, sera from mice immunized with IsaA, rabbit anti-SceD (kindly provided by S. Foster)17 or the IRDye 800 CW-labeled 1D9 anti-IsaA humAb6. The binding of unlabeled antibodies was visualized with fluorescently labeled goat anti-human, goat anti-mouse or donkey anti-rabbit IRDye 800 CW-labeled secondary antibodies (LI-COR Biosciences, Lincoln, NE. USA), using the Odyssey Infrared Imaging System (LI-COR Biosciences).
Cell wall extraction and binding experiments
Crude cell wall extracts were obtained from overnight cultures of S. aureus MA12 ΔisaA or S. aureus Newman ΔspaΔsbi. To this end, cell pellets were resuspended in demineralized water with 0.1 µm glass beads (Biospec Products, Bartlesville, USA) and disrupted in a Precellys 24 homogenizer (Bertin Technologies, France), followed by centrifugation at 4000 rpm (1520 × g) for 6 min. Recovered supernatants were centrifuged for 15 min at 14000 rpm (18620 x g) and pellets containing crude cell wall extracts were resuspended in demineralized water and stored at 4 °C. Non-covalently bound cell wall proteins were extracted from S. aureus Newman ΔspaΔsbi cells by a 10 min incubation with 1 M potassium thiocyanate (KSCN) as described previously36.
Growth medium fractions of overnight nisin-induced cultures expressing IsaA protein fragments were precipitated with 10% TCA. The resulting protein pellets were resuspended in 4-fold diluted phosphate buffered saline with 0.1% Tween 20 (PBS-T), and proteins were analyzed by LDS-PAGE. For rebinding experiments, crude cell wall extracts were incubated with IsaA protein fragments (obtained as described above) at room temperature for 10 min and, thereafter, the binding of the IsaA fragments was assessed by LDS-PAGE and Western blotting.
Near Infrared Microscopy
Aliquots of 1 ml of bacterial cultures (OD600 of 1) in TSB were collected, washed twice with PBS, and resuspended in 1 ml of PBS. Next, ~1010 cells thus obtained were incubated with 0.65 mg/ml IRDye 800CW-labeled 1D9 or 1D9 F(ab’)2 for 1 h at room temperature. After washing 3 times with PBS, cells were spotted on poly-L-lysine-coated glass slides (Sigma Aldrich) and inspected using a Leica DM5500B epifluorescence microscope equipped with an 800 nm filter block. Images were captured with a Leica DFC365FX camera using a 63× objective (Leica Microsystems BV, The Netherlands).
Epitope mapping with peptide arrays
To determine which epitopes of S. aureus IsaA were recognized by the investigated anti-IsaA IgGs, libraries of linear 15-mer IsaA-specific peptides were synthesized with an overlap on a solid support (Pepscan), as previously described37, 38. Next, the peptide libraries were probed with heat-inactivated mouse sera, humAb 1D9, or rabbit anti-IsaA in a dilution of 1:1000. Bound IsaA-specific antibodies were detected with a secondary HRP-conjugated antibody (Southern Biotech, Birmingham, USA) and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) (Sigma-Aldrich). A charge-coupled device camera was used to record the absorbance at 405 nm.
Immunization of mice with IsaA
Specified pathogen-free female BALB/cBYJ mice were obtained from Charles River (Saint-Germain-sur-l’Arbresle, France). Mice were treated and selected as described previously27. Purified IsaA-His6 was emulsified 1:1 with TiterMax Gold adjuvant (Sigma-Aldrich). Mice were immunized subcutaneously in the flank with 100 μL formulated vaccine on days -28, -21, and -14 (25 μg of antigen). Control mice received 100 μL PBS emulsified with adjuvant. At day -1, blood was withdrawn from the tail artery. Sera were examined by ELISA using ELISA plates (Greiner Bio-One B.V, Alphen aan den Rijn, the Netherlands), coating, blocking, hybridization and detection procedures as described previously27. Immunized mice (n = 6 immunized mice, n = 11 placebo-immunized mice) were challenged on day 0 by intravenous inoculation of 100 μL of S. aureus isolate P (3 × 105 CFU) as described previously27. Discomfort and animal survival rate over 14 days after infection were monitored. For discomfort score, clinical signs of illness in each mouse were evaluated at least twice daily as described before27.
Ethics statements
Blood donations from EB patients were collected under the approval of the medical ethics committee of the University Medical Center Groningen (approval no. NL27471,042,09) upon written informed patient consent, and with adherence to the Helsinki Guidelines23. The required written informed consent was obtained from all EB patients and healthy volunteers included in the present studies. The Institutional Animal Care and Use Committee of the Erasmus University Medical Centre Rotterdam approved the present protocols for animal experiments (permit number: EMC2694). All animal experiments were performed in accordance with the rules laid down in the Dutch Animal Experimentation Act and the Directive 2010/63/EU on the protection of animals used for scientific purposes.
Results
IsaA contains 2 conserved domains
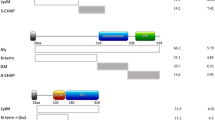
As a first approach towards the identification of antigenic epitopes in IsaA, we performed a bioinformatics analysis of the domain structure of this protein. Inspection of the amino acid sequence of IsaA using Pfam only showed the presence of the known soluble lytic transglycosylase domain (SLT, pfam01464) in the C-terminus (Fig. 1B). Subsequent BLASTP searches against the NCBI Protein Reference Sequences Database using the N-terminus of IsaA (residues 30–145) revealed another conserved domain ranging from residues 30 to 84. A following Position-Specific iterated BLAST search with this region of IsaA showed that this domain is exclusively present in the N-termini of 391 proteins, primarily the IsaA and SceD proteins from S. aureus and other staphylococcal species (Fig. 1A and Supplemental Fig. 1). In what follows, we refer to this N-terminal Conserved Domain as the NCD domain of IsaA.

Schematic representation of IsaA subdomains, their separate expression and immunodetection. (A) The N-terminal conserved domain (NCD) of IsaA (residues 30–84) as identified by BLASTP searches with the N-terminal region of IsaA (residues 30–145; see also Supplementary Fig. 1). (B) The mature IsaA, its N-terminal (IsaA-N, IsaA-N1, IsaA-N2) or C-terminal (IsaA-C, IsaA-C1, IsaA-C2) fragments were expressed in L. lactis PA1001 and secreted as N-terminal or C-terminal His-tag fusions using the lactococcal signal peptide of Usp45 (spu). The C-terminal part of IsaA contains a soluble lytic transglycosylase (SLT) domain (pfam01464). In the schematic representations of the expression vectors pNG4110 and pNG4210, used to produce IsaA or its subdomains, the positions of the cleavage sites for the restriction enzymes BamHI (B) and NotI (N), and the N-terminal or C-terminal His-tags (h 6) are indicated. Molecular weights (kDa) of IsaA and its subdomains and their isoelectric points (pI) are indicated. sp, IsaA signal peptide; *Stop codon provided within the expression vectors pNG4110 and pNG4210. (C) LDS-PAGE analysis of growth medium fractions of nisin-induced L. lactis cultures producing the C-terminally His-tagged IsaA protein or IsaA fragments. The His-tagged proteins are visualized either by Simply Blue gel-staining, or by Western blotting with the humAb 1D9–800CW, the polyclonal α-IsaA rabbit antibodies, or α-His-tag antibodies. Molecular weights (kDa) of the standard proteins are indicated on the left.
IsaA is non-covalently bound to the staphylococcal cell wall
To investigate the subcellular localization of IsaA, several experiments were performed. Fractionation and subsequent Western blotting analyses of S. aureus MA12, MA12ΔisaA and Newman ΔspaΔsbi showed that IsaA is both retained in the cell fraction and released into the culture medium (Fig. 2A). Furthermore, extraction of non-covalently cell wall-bound proteins from S. aureus Newman ΔspaΔsbi cells with KSCN resulted in the release of IsaA from the cells (Fig. 2A, lane labelled W). As KSCN is a chaotrope, the release of IsaA from the cells implies that this protein is associated with the cell wall through non-covalent cell wall interactions based on hydrogen bonds, van der Waals forces, or hydrophobic interactions (Fig. 2A). In contrast to IsaA, the IsaA paralogue SceD is mostly secreted into the growth medium by clinical S. aureus isolates that produce this protein (Fig. 2B). Of note, the full size 1D9–800CW antibody also binds to the IgG binding proteins Spa (protein A) and Sbi whereas, as expected, 1D9 F(ab’)2-800CW nanobodies do not bind to these proteins since they lack the Fc moiety needed for Spa- and Sbi-binding (Fig. 2A). Consistent with the Western blotting data, fluorescence microscopy showed that 1D9 F(ab)2-800CW binds to whole cells of the wild-type S. aureus MA12 strain, but not to cells of S. aureus MA12 ΔisaA (Fig. 3).

Cell-associated and secreted forms of IsaA and SceD. (A) S. aureus MA12, S. aureus MA12 ΔisaA and S. aureus Newman ΔspaΔsbi were grown overnight. Subsequently, cells (C) were separated from the growth medium (M) by centrifugation and proteins in the respective fractions were separated by LDS-PAGE. In addition, non-covalently cell wall-associated proteins (W) were extracted from S. aureus Newman ΔspaΔsbi and separated by LDS-PAGE. The presence of IsaA was subsequently visualized by Western blotting using humAb 1D9-800CW, hum Ab 1D9 F(ab)2-800CW or the polyclonal rabbit antibody α-IsaA. Molecular weights (kDa) of marker proteins are shown on the left and the positions of Protein A, Sbi and IsaA are indicated on the right. (B) Cellular (C) and secreted proteins in the growth medium (M) of the clinical S. aureus isolates (D,E,F,G and H) were separated by LDS-PAGE and detected by Western blotting using an α-SceD polyclonal antibody. Molecular weights (kDa) of marker proteins are shown on the left and the positions of protein A and SceD are indicated on the right.

Exposure of IsaA on the S. aureus cell surface. Phase contrast (left panels) and near-infrared fluorescence (right panels) microscopic images of overnight cultured S. aureus cells treated with 1D9 F(ab)2-800 CW. Top panels, S. aureus MA12 ΔisaA cells; bottom panels wild-type (wt) S. aureus MA12 cells.
To further investigate the potential of IsaA for cell wall binding, the mature protein and its N- and C-terminal fragments were expressed in L. lactis using vectors pNG4110 and pNG4210 as schematically represented in Fig. 1B. Specifically, this resulted in the nisin-inducible production of: the N-terminal conserved domain (N; residues 30–145), the N-terminal sub-domains N1 (residues 30–91) and N2 (residues 92–145), the C-terminal domain with the SLT active site domain, (C; residues 146–233), and the C-terminal fragments C1 (residues 146–189) and C2 (residues 190–233). These fragments were expressed either with a C-terminal His-tag or with an N-terminal His-tag. Further, all His-tagged fragments were attached to an N-terminal signal peptide, allowing their facile purification from the growth medium of L. lactis (Fig. 1C, simply blue staining panel and Western blot panel with α-His-tag antibodies). Of note, for unknown reasons the N and N2 fragments of IsaA had a lower mobility on the denaturing LDS-PAGE gels than expected based on their molecular weight. Analysis of the whole-cell and growth medium fractions of the L. lactis cultures expressing the different fragments or the full-size IsaA protein showed that mature IsaA and the N, N1 and C fragments can be detected both in the cells and the growth medium (Fig. 4A). The localization of these fragments was not affected by the presence of the His-tag at the N- or C-termini. In case of the N1 fragments most of the protein was present in the cells, suggesting that this IsaA moiety is of particular relevance for cell association. Conversely, relatively low amounts of the C fragment were localized to the cell fraction. Importantly, the N2, C1 and C2 fragments were exclusively detected in the growth medium fraction, suggesting that they have no important role in cell association (Fig. 4A).

Cell wall association of IsaA-derived protein fragments. (A) IsaA, its N-terminal (IsaA-N, IsaA-N1, IsaA-N2) and C-terminal (IsaA-C, IsaA-C1, IsaA-C2) fragments were expressed in L. lactis PA1001 with the Usp45 signal peptide for export from the cytoplasm and an N-terminal (←) or C-terminal (→) His-tag (see Fig. 1B). Next, cells (C) were separated from the growth medium (M) by centrifugation and the presence of full-size IsaA or its fragments in the respective fractions was analyzed by Western blotting using α-IsaA polyclonal rabbit antibodies. (B) The rebinding potential of secreted IsaA or IsaA-derived fragments to the S. aureus cell wall was assessed using full-size IsaA or IsaA-derived fragments (as described in A) and crude cell wall extracts from S. aureus MA12 ΔisaA as described in the Materials and Methods section. Upon incubation for 10 min, the cell walls were pelleted by centrifugation and proteins in the pelleted cell wall (W) and supernatant (S) fractions were analyzed by LDS-PAGE and Simply blue staining (upper panels) or Western blotting using α-IsaA polyclonal rabbit antibodies (lower panels). (C) As a control, IsaA was incubated without the crude cell wall extract from S. aureus MA12 ΔisaA and upon incubation centrifuged. Pellet and supernatant fractions were processed as described in (B). The molecular weights (kDa) of marker proteins are indicated on the left.
As the above cell association analysis was performed with the heterologous organism L. lactis, a cell wall rebinding analysis was performed using crude cell wall preparations of S. aureus MA12 ΔisaA to verify the cell wall binding capabilities of IsaA. This approach is based on the notion that non-covalently cell wall-bound proteins that have been released into the growth medium can in principle rebind to the S. aureus cell wall. In these experiments, we focused on S. aureus cell wall rebinding of the full-size IsaA, or the N or C fragments of IsaA, as purified from the growth medium of L. lactis. Upon incubation with the cell wall preparations, the cell walls were pelleted and the presence of IsaA (fragments) in the pellet and supernatant fractions was assessed by Western blotting. As shown in Fig. 4B, the full-size IsaA and the N fragment were capable of rebinding to the S. aureus cell wall, albeit that rebinding of the N fragment was less effective depending on the position of the His-tag. In contrast, the C fragment of IsaA did not rebind to the S. aureus cell wall. For control, the assay was performed also in the absence of cell wall extract. As presented in Fig. 4C, Simply Blue gel staining showed that hardly any IsaA was pelleted in the absence of cell wall extract, although a relatively small amount of pelleted IsaA was detectable by Western blotting. The latter probably represents a small amount of aggregated IsaA or IsaA bound to the tube. Altogether, the present observations show that IsaA has a modular structure composed of a conserved N-terminal domain and an active site domain in the C-terminus. Both domains are involved in effective binding of full-size IsaA to the S. aureus cell wall, albeit that the N-terminal domain can also bind to the cell wall in absence of the C-terminal domain.
HumAb 1D9 binds to the NCD domain of IsaA
The potentially different functions of the N and C domains of IsaA in cell wall binding and transglycosylase activity raised the question, which epitope(s) in IsaA need(s) to be recognized for a protective antibody response? As a first approach to answer this question, we assessed the binding site of our humAb 1D9 in IsaA using the different fragments of this protein expressed in L. lactis. As demonstrated by Western blotting, humAb 1D9 labeled with IRDye800CW binds effectively to the IsaA fragments N and N1, but not to the fragments N2, C, C1 and C2 (Fig. 1C). This shows that the epitope recognized by 1D9 resides within the first 62 residues of the mature IsaA protein. By contrast, all tested fragments of IsaA were bound by the polyclonal antibodies (α-IsaA) raised in a rabbit16 (Fig. 1C). To further narrow down the epitope recognized by 1D9, we applied the PepScan technology37, 38, where antibody binding to an array with linear 15-mer peptides representing the entire IsaA protein was tested. Unexpectedly, 1D9 did not bind to this array (not shown), while the polyclonal rabbit α-IsaA antibodies did bind to the array (Fig. 5). In fact, the profile of polyclonal α-IsaA binding to the peptide array mirrored the results obtained by Western blotting, where the N, N1, N2, C, C1 and C2 fragments of IsaA were all shown to bind the polyclonal antibodies (Fig. 1C). Taken together, these findings show that humAb 1D9 binds to an epitope within the conserved NCD domain of IsaA, and that this epitope is not represented in the array of linear 15-mer peptides. The latter finding suggests that 1D9 binding to IsaA relates to a feature with a particular secondary or tertiary structure.

Antibody reactivity against IsaA epitopes measured by pepscan-ELISA. Bar chart with relative fluorescence signals as detected upon binding of α-IsaA polyclonal rabbit antibodies to 15-mer peptides with 14 residue overlap. The peptides are ordered on the x-axis according to the position of their N-terminal residue in the overall IsaA amino acid sequence. The relative positions of the IsaA fragments N1, N2, C1 and C2 are indicated. The y-axis shows the OD (in AU).
Distinction of IsaA epitopes recognized by human sera and sera from IsaA-immunized mice
Our previous mouse immunization study with an octavalent anti-S. aureus vaccine including IsaA showed that antibodies against IsaA were not protective27. This previous conclusion was verified by immunizing mice with IsaA only, which showed that high IsaA-specific IgG titers did not protect mice against a challenge with the methicillin-sensitive S. aureus (MSSA) isolate P (Supplementary Figs 2 and 3). To determine whether the IsaA epitopes recognized by potentially protective IsaA-specific IgGs from six EB patients, and six non-protective IsaA-specific IgGs from immunized mice are similar or different, we employed a Western blotting approach. The results in Fig. 6 show that all tested sera of mice and men contained IgGs recognizing the full-size IsaA and the IsaA fragment N. All sera from mice contained IgGs recognizing fragment C, whereas this was significantly less common for the sera from EB patients. None of the tested sera contained IgGs recognizing fragment C2, and only two of the tested mouse sera contained IgGs recognizing fragment C1. Intriguingly, a distinguishing feature of the EB patient sera was that, with exception of the EB patient serum 51, all contained significant levels of IgGs against the N1 fragment (between ~22 to 41% of the total signal). Overall, this suggests that the IsaA-specific IgGs of EB patients preferentially recognize the N1 fragment of IsaA, similar to the protective humAb 1D9, whereas the non-protective IsaA-specific IgGs of immunized mice preferentially recognize the C fragment of IsaA (Fig. 6).

Differential binding of human and murine IgGs to IsaA-derived fragments. Proteins in the growth medium fractions of L. lactis PA1001 producing the full-size IsaA or N-terminal (N, N1, N2) or C-terminal (C, C1, C2) fragments of IsaA were separated by LDS-PAGE. Next, a Western blotting analysis was performed using sera from EB patients (EB01, EB09, EB11, EB15, EB51 and EB55) or sera from the mice immunized with IsaA (M31, M32, M33, M34, M35 and M36). The molecular weights (kDa) of marker proteins are indicated on the left. The intensities of detected bands were assessed with ImageJ, and the relative intensity for each band is indicated below each blot as the percentage of the total intensity determined for full-size IsaA plus the N-terminal (N, N1, N2) and C-terminal (C, C1, C2) fragments.
Discussion
The aim of this study was to localize immunogenic epitopes in the IsaA protein of S. aureus and to correlate the recognition of these epitopes by particular IgGs to protection against S. aureus infection. The results show that IsaA-specific human IgGs predominantly target a conserved N-terminal domain in IsaA, while IsaA-specific murine IgGs predominantly target the conserved C-terminal domain of IsaA.
Previous studies have shown that the C-terminal part of IsaA contains a soluble lytic transglycosylase (SLT) domain (Pfam 06737) that is conserved also in the SceD protein of S. aureus. This SLT domain is able to cleave peptidoglycan, and impacts on the clumping and separation of S. aureus cells17. In contrast, a possible function of the N-terminal region of IsaA was so far not known. Of note, our present data show that this N-terminal region contains a domain (NCD) that is well conserved in the IsaA and SceD proteins of a wide range of Staphylococcus species. Using proteomics approaches, it was previously shown that IsaA and SceD both are non-covalently cell wall-bound proteins18, 39,40,41. As it is known that most peptidoglycan hydrolases are composed of a catalytic site domain and a cell wall binding domain42, we hypothesized that the NCD domain could contribute to cell wall binding. Indeed, separate expression of the different regions of IsaA showed that the N-region, which includes the NCD domain, has affinity for cell walls of S. aureus and L. lactis. In contrast, the C-terminal region does apparently not bind cell walls by itself. Yet, the highest levels of cell wall binding were observed for the intact IsaA protein, indicating that both the N- and C-regions are involved in cell wall binding with a potentially more prominent contribution from the N-region. It should be noted, however, that a substantial portion of IsaA is secreted into the growth medium, and this is also the case for SceD. Consistent with the observed cell wall binding of IsaA, we did observe localization of this protein to the S. aureus cell surface using an immunofluorescence microscopy approach. Of note, this involved an IRDye 800CW-labeled F(ab)2 fragment of the humAb 1D9, which binds to the N1 subdomain of IsaA. Together, these observations imply that the N-domain of IsaA is exposed on the S. aureus cell surface, where it is accessible to IgG.
Intriguingly, our present results show that sera of patients with the genetic blistering disease EB contain IgGs that predominantly target the N-domain of IsaA. Chronic wounds of these patients are heavily colonized with S. aureus 43, 44, which was previously shown to elicit IgG levels against S. aureus proteins that were much higher than the IgG levels against the respective proteins in healthy volunteers23. The highest IgG levels were detected against the IsaA protein, leading to the view that these IgGs could be protective against S. aureus infections23. This view is supported by the fact that EB patients rarely suffer from invasive S. aureus infections despite the high-level colonization of their wounds45, and by the observation that monoclonal antibodies against IsaA (i.e. 1D9 and UK-66) can be protective against S. aureus infection in mouse models6,7,8. Of note, our present study also shows that that the protective humAb 1D9 also recognizes the N-domain of IsaA. In contrast to our findings with the sera of EB patients, the immunization of mice with purified IsaA did not elicit the production of IgGs that protect against S. aureus infection. In this case, the IsaA-specific IgGs predominantly recognized the C-terminal region of IsaA as shown in our present study. Together, these observations imply that IgGs against the C-terminal region of IsaA do not effectively protect against S. aureus infection. On the other hand, it is presently not possible to conclude that the high titers of anti-IsaA antibodies in EB patients, which bind to the N-terminal domain of IsaA, are indeed protective. For example, antibodies against other S. aureus components that are also present at high levels could contribute to the protection of EB patients against S. aureus bacteremia. In addition, it is also conceivable that differences in cell-medicated immunity contribute to the apparent protection of EB patients against S. aureus.
A key question that remains to be answered is why does there appear to be a bias towards IgGs specific for the N-terminal region of IsaA in EB patient sera, and a bias towards IgGs specific for the C-terminal region in immunized mice? Our present studies show that the N-terminal region of IsaA is exposed on the cell surface of S. aureus, which might suggest that this domain is better exposed to the human immune system than the C-terminal region of IsaA. Yet, our previous proteomics study, where S. aureus cells were shaved with trypsin that was immobilized on agarose beads, clearly showed that also the C-terminus of IsaA is exposed to the S. aureus cell surface12, 18. This makes a differential exposure of the N- and C-terminal regions of IsaA on the S. aureus cell surface a less likely explanation for the observed differences in the IgG specificities from mice and men. An alternative explanation could be that the immunogenicity of the N- and C-terminal domains of IsaA is different, depending on whether the immune system is challenged with IsaA attached to the S. aureus cell surface or with soluble IsaA. Conceivably, such a difference could relate to conformational differences in the respective domains when they are associated with the cell or secreted into the growth medium.
Conclusion
Altogether, our present study shows that the immune responses to IsaA are very different in the investigated mouse and human sera. In particular, the results seem to suggest that IgGs against the N-terminal domain of IsaA are potentially protective against S. aureus infection. This focuses attention on the N-terminal region of IsaA as a potential component in a future vaccine against the important human pathogen S. aureus. To validate this idea, protection studies need to be performed in an appropriate animal model, where the separated N- and C-terminal fragments of IsaA are used for active immunization.
References
Pozzi, C. et al. Vaccines for Staphylococcus aureus and Target Populations. Curr. Top. Microbiol. Immunol., doi:10.1007/82_2016_54 (2017).
Bagnoli, F. et al. Vaccine composition formulated with a novel TLR7-dependent adjuvant induces high and broad protection against Staphylococcus aureus. Proc. Natl. Acad. Sci. 112, 3680–3685 (2015).
Proctor, R. A. Recent developments for Staphylococcus aureus vaccines: clinical and basic science challenges. Eur. Cell. Mater. 30, 315–326 (2015).
Sause, W. E., Buckley, P. T., Strohl, W. R., Lynch, A. S. & Torres, V. J. Antibody-Based Biologics and Their Promise to Combat Staphylococcus aureus Infections. Trends Pharmacol. Sci. 37, 231–241 (2016).
Missiakas, D. & Schneewind, O. Staphylococcus aureus vaccines: Deviating from the carol. J. Exp. Med. 213, 1645 (2016).
van den Berg, S. et al. A human monoclonal antibody targeting the conserved staphylococcal antigen IsaA protects mice against Staphylococcus aureus bacteremia. Int. J. Med. Microbiol. 305, 55–64 (2015).
Lorenz, U. et al. Functional Antibodies Targeting IsaA of Staphylococcus aureus Augment Host Immune Response and Open New Perspectives for Antibacterial Therapy. Antimicrob. Agents Chemother. 55, 165–173 (2011).
Oesterreich, B. et al. Characterization of the biological anti-staphylococcal functionality of hUK-66 IgG1, a humanized monoclonal antibody as substantial component for an immunotherapeutic approach. Hum. Vaccines Immunother. 10, 926–937 (2014).
Stentzel, S., Gläser, R. & Bröker, B. M. Elucidating the anti-Staphylococcus aureus antibody response by immunoproteomics. PROTEOMICS – Clin. Appl. 10, 1011–1019 (2016).
Thammavongsa, V., Rauch, S., Kim, H. K., Missiakas, D. M. & Schneewind, O. Protein A-neutralizing monoclonal antibody protects neonatal mice against Staphylococcus aureus. Vaccine 33, 523–526 (2015).
Thomer, L. et al. Antibodies against a secreted product of Staphylococcus aureus trigger phagocytic killing. J. Exp. Med. 213, 293–301 (2016).
Dreisbach, A., van Dijl, J. M. & Buist, G. The cell surface proteome of Staphylococcus aureus. Proteomics 11, 3154–3168 (2011).
Lorenz, U. et al. Human antibody response during sepsis against targets expressed by methicillin resistant Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 29, 145–153 (2000).
Sibbald, M. J. J. B. et al. Mapping the Pathways to Staphylococcal Pathogenesis by Comparative Secretomics. Microbiol. Mol. Biol. Rev. 70, 755–788 (2006).
Ziebandt, A.-K. et al. Proteomics uncovers extreme heterogeneity in the Staphylococcus aureus exoproteome due to genomic plasticity and variant gene regulation. Proteomics 10, 1634–1644 (2010).
Sakata, N., Terakubo, S. & Mukai, T. Subcellular Location of the Soluble Lytic Transglycosylase Homologue in Staphylococcus aureus. Curr. Microbiol. 50, 47–51 (2005).
Stapleton, M. R. et al. Characterization of IsaA and SceD, Two Putative Lytic Transglycosylases of Staphylococcus aureus. J. Bacteriol. 189, 7316–7325 (2007).
Dreisbach, A. et al. Profiling the surfacome of Staphylococcus aureus. PROTEOMICS 10, 3082–3096 (2010).
Islam, N., Ross, J. M. & Marten, M. R. Proteome Analyses of Staphylococcus aureus Biofilm at Elevated Levels of NaCl. Clin. Microbiol. Los Angel. Calif 4, doi:10.4172/2327-5073.1000219 (2015).
Reijer, P. M. den et al. Detection of Alpha-Toxin and Other Virulence Factors in Biofilms of Staphylococcus aureus on Polystyrene and a Human Epidermal Model. PLOS ONE 11, e0145722, doi:10.1371/journal.pone.0145722 (2016).
Glasner, C. et al. Low anti-staphylococcal IgG responses in granulomatosis with polyangiitis patients despite long-term Staphylococcus aureus exposure. Sci. Rep. 5, 8188, doi:10.1038/srep08188 (2015).
Kloppot, P. et al. Microarray-based identification of human antibodies against Staphylococcus aureus antigens. PROTEOMICS – Clin. Appl. 9, 1003–1011 (2015).
van der Kooi-Pol, M. M. et al. High Anti-Staphylococcal Antibody Titers in Patients with Epidermolysis Bullosa Relate to Long-Term Colonization with Alternating Types of Staphylococcus aureus. J. Invest. Dermatol. 133, 847–850 (2013).
Swierstra, J. et al. IgG4 Subclass-Specific Responses to Staphylococcus aureus Antigens Shed New Light on Host-Pathogen Interaction. Infect. Immun. 83, 492–501 (2015).
van den Berg, S. et al. A multiplex assay for the quantification of antibody responses in Staphylococcus aureus infections in mice. J. Immunol. Methods 365, 142–148 (2011).
van den Berg, S., de Vogel, C. P., van Belkum, A. & Bakker-Woudenberg, I. A. J. M. Mild Staphylococcus aureus Skin Infection Improves the Course of Subsequent Endogenous S. aureus Bacteremia in Mice. PLoS ONE 10 (2015).
van den Berg, S. et al. Active Immunization with an Octa-Valent Staphylococcus aureus Antigen Mixture in Models of S. aureus Bacteremia and Skin Infection in Mice. PLoS ONE 10, e0116847, doi:10.1371/journal.pone.0116847 (2015).
Selle, M. et al. Global antibody response to Staphylococcus aureus live-cell vaccination. Sci. Rep. 6, 24754, doi:10.1038/srep24754 (2016).
Altschul, S. F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402 (1997).
Altschul, S. F. et al. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 272, 5101–5109 (2005).
Finn, R. D. et al. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 44, D279–D285 (2016).
Papadopoulos, J. S. & Agarwala, R. COBALT: constraint-based alignment tool for multiple protein sequences. Bioinformatics 23, 1073–1079 (2007).
Wheeler, T. J., Clements, J. & Finn, R. D. Skylign: a tool for creating informative, interactive logos representing sequence alignments and profile hidden Markov models. BMC Bioinformatics 15, 7 (2014).
Neef, J., Koedijk, D. G. A. M., Bosma, T., Dijl, J. Mvan & Buist, G. Efficient production of secreted staphylococcal antigens in a non-lysing and proteolytically reduced Lactococcus lactis strain. Appl. Microbiol. Biotechnol. 98, 10131–10141 (2014).
Leenhouts, K. & Venema, G. Lactococcal plasmid vectors. Plasmids Pract. Approach Oxf. Univ. Press Oxf. UK 65–94 (1993).
Sibbald, M. J. J. B. et al. Synthetic Effects of secG and secY2 Mutations on Exoproteome Biogenesis in Staphylococcus aureus. J. Bacteriol. 192, 3788–3800 (2010).
Timmerman, P. et al. Mapping of a discontinuous and highly conformational binding site on follicle stimulating hormone subunit-β (FSH-β) using domain ScanTM and Matrix ScanTM technology. Mol. Divers. 8, 61–77 (2004).
Timmerman, P., Puijk, W. C. & Meloen, R. H. Functional reconstruction and synthetic mimicry of a conformational epitope using CLIPSTM technology. J. Mol. Recognit. 20, 283–299 (2007).
Gatlin, C. L. et al. Proteomic profiling of cell envelope-associated proteins from Staphylococcus aureus. PROTEOMICS 6, 1530–1549 (2006).
Glowalla, E., Tosetti, B., Krönke, M. & Krut, O. Proteomics-Based Identification of Anchorless Cell Wall Proteins as Vaccine Candidates against Staphylococcus aureus. Infect. Immun. 77, 2719–2729 (2009).
Solis, N., Larsen, M. R. & Cordwell, S. J. Improved accuracy of cell surface shaving proteomics in Staphylococcus aureus using a false-positive control. PROTEOMICS 10, 2037–2049 (2010).
Visweswaran, G. R. R., Leenhouts, K., Roosmalen, Mvan, Kok, J. & Buist, G. Exploiting the peptidoglycan-binding motif, LysM, for medical and industrial applications. Appl. Microbiol. Biotechnol. 98, 4331–4345 (2014).
van der Kooi-Pol, M. M. et al. High genetic diversity of Staphylococcus aureus strains colonizing patients with epidermolysis bullosa. Exp. Dermatol. 21, 463–466 (2012).
van der Kooi-Pol, M. M. et al. Topography of Distinct Staphylococcus aureus Types in Chronic Wounds of Patients with Epidermolysis Bullosa. PLoS ONE 8, e67272 (2013).
van der Kooi-Pol, M. M., Duipmans, J. C., Jonkman, M. F. & van Dijl, J. M. Host–pathogen interactions in epidermolysis bullosa patients colonized with Staphylococcus aureus. Int. J. Med. Microbiol. 304, 195–203 (2014).
Bosma, T. et al. Novel Surface Display System for Proteins on Non-Genetically Modified Gram-Positive Bacteria. Appl. Environ. Microbiol. 72, 880–889 (2006).
Kreiswirth, B. N. et al. The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 305, 709–712 (1983).
Neef, J. et al. Versatile vector suite for the extracytoplasmic production and purification of heterologous His-tagged proteins in Lactococcus lactis. Appl. Microbiol. Biotechnol. 99, 9037–9048 (2015).
Acknowledgements
We thank the anonymous patients with EB from the Dutch Epidermolysis Bullosa Registry for blood donations, and Magda van der Kooi-Pol, José Duipmans and Marcel Jonkman for collecting the EB patient sera in a previous study. We thank Sjouke Piersma for his help in the Western data analysis, Simon Foster for providing anti-SceD antibodies, and N. Sakata for providing anti-IsaA polyclonal rabbit antibodies. This research was supported by the Top Institute Pharma projects T4-213 and T4-502. F.R.P. received a scholarship from CONACyT (169643) and was supported in parts by the Graduate School for Medical Sciences of the University of Groningen.
Author information
Authors and Affiliations
Contributions
D.G.A.M.K., F.R.P., J.W.B., I.A.J.M.B.-W., G.B. conceived and designed the experiments. D.G.A.M.K., F.R.P., H.H., S.v.d.B., J.W.B., C.K., R.C.P. performed the experiments. D.G.A.M.K., F.R.P., H.H., J.W.B., J.M.v.D., G.B. analyzed the data. D.G.A.M.K., F.R.P., S.v.d.B., J.W.B., I.A.J.M.B.-W., J.M.v.D., G.B. contributed reagents, materials and analysis tools. D.G.A.M.K., F.R.P., J.M.v.D., G.B. wrote the manuscript. All authors have reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
JWB is an employee of Pepscan Therapeutics BV. All other authors declare no conflicts of interest.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Koedijk, D.G.A.M., Pastrana, F.R., Hoekstra, H. et al. Differential epitope recognition in the immunodominant staphylococcal antigen A of Staphylococcus aureus by mouse versus human IgG antibodies. Sci Rep 7, 8141 (2017). https://doi.org/10.1038/s41598-017-08182-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-08182-9
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
