
Abstract
Despite being fundamental to multiple biological processes, phosphorus (P) availability in marine environments is often growth-limiting, with generally low surface concentrations. Picocyanobacteria strains encode a putative ABC-type phosphite/phosphate/phosphonate transporter, phnDCE, thought to provide access to an alternative phosphorus pool. This, however, is paradoxical given most picocyanobacterial strains lack known phosphite degradation or carbon-phosphate lyase pathway to utilise alternate phosphorus pools. To understand the function of the PhnDCE transport system and its ecological consequences, we characterised the PhnD1 binding proteins from four distinct marine Synechococcus isolates (CC9311, CC9605, MITS9220, and WH8102). We show the Synechococcus PhnD1 proteins selectively bind phosphorus compounds with a stronger affinity for phosphite than for phosphate or methyl phosphonate. However, based on our comprehensive ligand screening and growth experiments showing Synechococcus strains WH8102 and MITS9220 cannot utilise phosphite or methylphosphonate as a sole phosphorus source, we hypothesise that the picocyanobacterial PhnDCE transporter is a constitutively expressed, medium-affinity phosphate transporter, and the measured affinity of PhnD1 to phosphite or methyl phosphonate is fortuitous. Our MITS9220_PhnD1 structure explains the comparatively lower affinity of picocyanobacterial PhnD1 for phosphate, resulting from a more limited H-bond network. We propose two possible physiological roles for PhnD1. First, it could function in phospholipid recycling, working together with the predicted phospholipase, TesA, and alkaline phosphatase. Second, by having multiple transporters for P (PhnDCE and Pst), picocyanobacteria could balance the need for rapid transport during transient episodes of higher P availability in the environment, with the need for efficient P utilisation in typical phosphate-deplete conditions.
Similar content being viewed by others

Introduction
Phosphorus (P) is an essential biological building block, integral for processes such as energy use (ATP), cell structure (phospholipids), and storage of genetic information in nucleic acids [1]. Dissolved P is also vital to the biogeochemistry of marine environments. The availability of the primary bioavailable form of P in its most oxidised state (+5), the inorganic phosphate ion (Pi), significantly influences the growth, abundance, and diversity of the most abundant photosynthetic microorganisms on Earth, marine picocyanobacteria of the genera Prochlorococcus and Synechococcus [2,3,4]. Low-nanomolar concentrations of Pi have been reported in various marine environments, which could limit picocyanobacterial growth in these areas [5].
Marine picocyanobacteria have a well-characterised response to Pi limitation [5,6,7], leading to significant alterations in gene expression, particularly of the phosphate regulation (Pho regulon) network. The nature and organisation of the Pho regulon is highly variable, even between related picocyanobacteria [7], corresponding to different adaptation strategies between different ecotypes [4, 6]. Genes encoding the high-affinity multicomponent periplasmic binding protein-dependent Pi specific transporter (Pst) are shown to be significantly up-regulated under Pi starvation in marine picocyanobacteria [7]. Marine picocyanobacterial genomes lack genes encoding the low-affinity constitutively expressed single-component Pi symporter system (Pit) found in E. coli [4, 8, 9]. It is hypothesised that Pi uptake via the Pit system could be energetically more favourable due to the symport with protons at 1:1 stoichiometry as opposed to Pi uptake via the Pst system that potentially requires hydrolysis of two ATP per substrate transported [10].
Picocyanobacterial strains also encode a well-conserved predicted ABC-type phosphite/phosphate/phosphonate transporter, PhnD1CE, which is thought to provide access to an alternative P pool such as organic phosphonates, Pn (P valence +3) or the inorganic reduced P compound phosphite, Pt (P valence +3) [5]. The ability to uptake alternate P sources could provide significant competitive advantages under Pi-depleted conditions. Physiological studies on picocyanobacteria, however, demonstrate variability in the expression of the PhnD1CE transport system under P stress. While some picocyanobacterial strains induce the expression of PhnD1CE transporter in a P-deficient media as shown for Synechococcus WH8102 [11], in some Prochlorococcus strains, it is not induced under P limitation [12] or can be constitutively expressed [11]. Several picocyanobacterial strains also encode an additional PhnCD2E transporter that clusters distinctly on protein phylogenetic trees [2, 3].
As the periplasmic substrate-binding component of the two predicted picocyanobacterial Pn transporters, PhnD1 and PhnD2, are typically co-expressed with the PhnC and PhnE components, the binding proteins have been used as proxies to investigate the substrate specificities of the respective transporter to understand their role in P acquisition [2, 3, 13]. Biophysical and structural data on PhnD1 and PhnD2, isolated from Prochlorococcus MIT9301, show diverse substrate specificity. While Prochlorococcus MIT9301_PhnD1 shows a high affinity for Pt with a binding affinity (KD) in the sub-micromolar range and comparatively weaker affinity to Pi and methylphosphonate (MPn), Prochlorococcus PhnD2 show a stronger affinity for MPn followed by Pt [2, 3, 13]. Unlike the Prochlorococcus PhnD proteins, there is a distinct lack of understanding regarding the specificity range of PhnD proteins within Synechococcus strains.
For various Synechococcus isolates, low P quotas and high uptake rates underscore the intense P competition in nutrient deplete oligotrophic environments [5, 14]. Low available P concentrations exert an intense selective pressure that influences the repertoire of P acquisition mechanisms of the related Prochlorococcus strains [12], most of which are also shared by Synechococcus isolates [5]. Several genes responsible for P acquisition have been acquired horizontally [12], indicating acquisition strategies between different strains likely reflect local P availability.
Synechococcus strains from clade I (e.g. CC9311) and IV generally co-occur in coastal and/or temperate mesotrophic open ocean waters, largely above 30°N and below 30°S [15, 16], alongside a broad nitrate and phosphate concentration range (0.03 to 14.5 µM and 0.2 to 1.2 µM, respectively) [5]. In contrast, Synechococcus clade II strains (e.g. CC9605) are abundant in coastal/continental shelves strictly in the subtropical/tropical latitudes (between 30°N and 30°S) [15,16,17,18]. Synechococcus clade III isolates (e.g. WH8102) do not show any latitudinal preference but are restricted to limited nitrate and phosphate concentrations [15], whereas clades CRD1 (e.g. MITS9220) and CRD2 are most successful in low Fe waters [19]. We examined PhnD1 from four marine Synechococcus strains (CC9311, CC9605, WH8102 and MITS9220) isolated from diverse environmental niches to characterise their corresponding ligand binding preferences and to explore the relationship between PhnD1 ligand specificity range and ecological niches.
Results and discussion
PhnD1 proteins in picocyanobacteria are lineage partitioned
The predicted phosphonate-binding protein (PhnD1) under study is found within the genome of all 97 sequenced picocyanobacteria strains in the Cyanorak database, comprising cluster CK_860 [20]. While the gene encoding PhnD1 is highly conserved among all picocyanobacterial genomes, our phylogenetic tree reveals it is distinctly partitioned by lineage, with Prochlorococcus representatives group separately from Synechococcus isolates and each clustering into clade-level groups (Fig. 1A). This highlights that the PhnD1 gene has been retained across evolutionary pressures and likely propagated by vertical transfer rather than horizontal gene transfer events. The latter is unlike the predicted duplicate copies, PhnD2 (cluster CK_6203; also annotated as PtxB [2, 3]) present in only a small number of strains (Prochlorococcus HLII/LLIV and Synechococcus clade II isolates) and a single PhnD3 (cluster CK_56876; only found in Prochlorococcus MIT9314) that cluster remotely from the PhnD1 genes. The low conservation of PhnD2 and PhnD3 indicates these are likely laterally transferred or results of lineage-specific gene duplications among ecotypes in similar environmental niches.

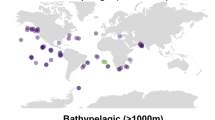
A The amino acid sequences of Synechococcus (Syn) and Prochlorococcus (Pro) PhnD1 (CK_860), PhnD2 (CK_6203) and PhnD3 (CK_56876), as well as previously characterised non-picocyanobacterial PhnD proteins were aligned using MAFFT [40], and the phylogenetic tree inferred using IQ-Tree [41]. The final phylogenetic tree was visualised using iTOL [42]. The four Synechococcus PhnD1 proteins (CC9311, CC9605, WH8102 and CRD1a) characterised in this study are highlighted on the tree. Genes are numbered according to their Cyanorak cluster designation [20]. B The environmental abundance for Prochlorococcus MIT9301_PhnD1 (top-left) and MIT9301_PhnD2 (bottom-left) picocyanobacterial homologues extracted from the Tara Oceans MetaT dataset [57]. The transcript abundance is plotted for surface waters, with a circle size corresponding to the measured abundance at a particular sampling site. Sampling sites are denoted by an ‘X’. The corresponding bubble plot (right) for the identified transcripts across sampling depths (SRF, surface waters; DCM, deep chlorophyll maximum; MES, mesopelagic zone; MIX, marine epipelagic mixed layers;) is depicted as a function of measured phosphate concentration. The Krona plot in the inset shows the taxonomic distribution of MIT9301_PhnD1, and MIT9301_PhnD2 homologues selected to analyse the metatranscriptome abundance.
To evaluate the distribution and expression of PhnD1 and PhnD2 in marine picocyanobacterial populations, we investigated the metagenomes and metatranscriptomes available from the Ocean Microbial Reference Gene Catalogue (OM-RGC) [21, 22]. For comparison, we provide distribution maps for Synechococcus and Prochlorococcus lineage-specific genes (Supplementary Fig. S1A, B). This analysis revealed that picocyanobacterial PhnD1 transcripts (Fig. 1B) and genes (Supplementary Fig. S1C, D) are not only enriched in mostly Pi-replete waters but also some Pi-deplete areas across several Pacific Ocean, Atlantic Ocean and Indian Ocean sites (surface water as well as the deep chlorophyll maximum zone). In contrast, picocyanobacterial PhnD2 homologues were most abundant in Pi-limited surface waters in the North-western Atlantic Ocean, the Mediterranean Sea and the Gulf of Mexico.
Previous studies show that Prochlorococcus MIT9301, which encodes both phnD1 and phnD2, can use Pt or MPn as the sole P source to support its growth [2, 3, 23]. The ability of Prochlorococcus MIT9301 to grow on Pt or MPn is attributed to the predicted phosphonate oxidative pathway encoded by the putative phnY (cluster CK_55307) and phnZ (cluster CK_7402) genes [23, 24] and a NAD-dependent phosphite dehydrogenase ptxD (cluster CK_56808), respectively. These predicted Pn and Pt utilisation genes are co-located with the phnC2D2E2 (also annotated as ptxABC) transport system in some picocyanobacterial strains. Based on these observations, we hypothesise that, by extension, all picocyanobacterial isolates encoding the phnC2D2E2 and the adjacent phnY, phnZ and ptxD genes have the potential to metabolise Pt or MPn, providing an ecological advantage to survive in a Pi-depleted environment. Our comparative genome analysis reveals Prochlorococcus and Synechococcus genomes sequenced to date, however, lack the phnY, phnZ and ptxD genes, suggesting these strains may not be able to utilise Pt or MPn.
Based on the in vitro studies showing a very high affinity of Prochlorococcus MIT9301 PhnD1 to Pt [3, 13], it is predicted that the transporter encoded by phnD1C1E1 could be a phosphite transporter. However, studies show that Prochlorococcus strains MIT9313 and MED4 (lacking the phnC2D2E2 transport system and the adjacent metabolising genes) are unable to grow and utilise Pt (or MPn) and instead are only able to utilise Pi for their growth [2, 3, 23]. Other studies examining the P-starvation response in picocyanobacteria indicate none of the components of the phnD1C1E1 transport system showed any significant upregulation in P-deplete conditions, as opposed to the strong induction of genes previously implicated in P-scavenging, such as the high-affinity pstS gene [7, 12].
We examined the phnD1C1E1 genome context and find a highly conserved gene (tesA; cluster CK_171), encoding a predicted lysophospholipase L1-like esterase, co-located in almost all picocyanobacterial sequenced genomes. While the function of the periplasmic TesA in picocyanobacteria is unclear, it has been previously proposed to be part of a complex enzymatic system responsible for phospholipid membrane homeostasis in P. aeruginosa [25]. Picocyanobacteria (Prochlorococcus MED4 and Synechococcus WH8102, WH7803, WH5701) are shown to have the ability to substitute sulfolipids for phospholipids in their membrane to economise P utilisation under P-limitation [26]. The same study also showed that while the “substitute lipids” are dominant in the P-limited cultures, the phospholipid substitution occurred in all cases, including in the P-replete conditions, although to a much smaller extent. The genes required for sulfolipid biosynthesis, sqdB (cluster CK_123) and sqdX (cluster CK_333), are conserved in all sequenced picocyanobacterial genomes and not just those abundant in the P-deplete conditions. It remains unclear if the conservation of sulfolipid biosynthesis genes (sqdB, sqdX) and the co-localisation of the phnD1C1E1 transporter with conserved tesA are correlated and what physiological advantage in terms of P utilisation this would provide to the marine picocyanobacterial strains. It is possible that TesA could play a role in breaking down phospholipids found in picocyanobacteria, such as the prevalent phosphatidylglycerol (PG). While the specific breakdown products of PG in cyanobacteria are currently unknown, in yeast, PG can be degraded to diacylglycerol and glycerol-3-phosphate (G3P) [27]. In this case, alkaline phosphatase could then cleave the phosphoryl group from the glycerol-3-phosphate, producing glycerol and inorganic phosphate (Pi), which could be taken up by the PhnDCE or Pst transport system, setting the stage for a constant phosphorus supply irrespective of the environmental conditions.
Synechococcus PhnD1 proteins show a preferential binding affinity for phosphite, phosphate and methylphosphonate
Several studies have examined the P-binding specificities for different PhnD proteins in Prochlorococcus [2, 3, 13]. This is the first study to report the substrate specificity range of Synechococcus PhnD1 proteins. We expressed and purified four Synechococcus PhnD1 proteins (CC9311_PhnD1, CC9605_PhnD1, MITS9220_PhnD1 and WH8102_PhnD1; Supplementary Fig. S2) and evaluated their potential ligand binding preferences by differential scanning fluorimetry (DSF). This was done by measuring the increase in melting temperature (TM) of the protein in the presence of potential ligands. The first-pass ligand screen encompassed compounds (Supplementary Table S1), including a diverse range of biologically active small molecules, trace metals, and common metabolic nutrients (C, N, S, P sources) as well as inorganic phosphate (Pi), phosphite (Pt), methylphosphonate (MPn), and hypophosphite (HPt). This comprehensive DSF screen reveals (Fig. 2A) all four PhnD1 proteins have higher thermal stability (ΔTM 5–23 °C) only in cocktail conditions comprising P sources. Among the single P-sources tested, the most significant shift in the TM is observed in the presence of Pt (ΔTM 17–23 °C), followed by Pi (ΔTM 3–8 °C). A second DSF screen comprising 60 distinct individual P-sources were tested for CC9605_PhnD1 protein, including those representing the hydrophilic polar headgroups of various phospholipids such as 3-phospho glyceric acid, phosphoryl choline, inositol phosphate and phospho-serine (Supplementary Fig. S3). With the exception of carbamyl phosphate and phospho glycolic acid, which may act as a phosphate proxy due to their comparatively smaller size, no other phosphorous sources showed a marked increase in the thermal stability, indicating they are unlikely to be PhnD1 substrates.

A DSF thermal melt assay was used to screen ligands for CC9311_PhnD1 (yellow), CC9605_PhnD1 (blue), MITS9220_PhnD1 (red) and WH8102_PhnD1 (green) proteins in the presence of a range of cocktail solutions (Silver Bullets 96-well screen) containing unique biologically relevant potential substrates as well as with various P sources (Pi, Pt, MPn and HPt). These data show that significant change in the melting temperature (ΔTM) is conferred only in cocktail conditions comprising P sources. B The measured binding affinity of the four Synechococcus PhnD1 proteins calculated based on the isothermal analysis of the DSF data (Supplementary Figs. S4–S7) upon incremental addition of the respective ligand; Pi (red), Pt (green), MPn (blue) or HPt (orange) is depicted. Reported affinities were computed using the Python package [29] and plotted on GraphPad Prism.
A fluorometric isothermal approach [28, 29] was further utilised to determine the binding dissociation constants (KD) of the four Synechococcus PhnD1 proteins in the presence of Pi, Pt, MPn, and HPt (Fig. 2B). This approach involves measuring incremental changes in the thermal stability (Supplementary Figs. S4–S7) caused by the partial formation of protein-ligand complexes, measured by an extrinsic dye [28, 29]. All four Synechococcus PhnD1 proteins displayed an affinity for at least two P sources: Pi and Pt (Fig. 2B); however, the specific binding affinities for these differ between each strain, as summarised in Table 1. Of the four proteins under study, MITS9220_PhnD1 displayed a comparatively stronger affinity for Pt and Pi. The derived KD value for Pt is at least an order of magnitude stronger than Pi for each PhnD1 protein, indicating a markedly stronger affinity for Pt. This is intriguing, given the fact that most strains of picocyanobacteria lack known Pt utilisation proteins.
Except for the CC9311_PhnD1, isolated from the strain found in the mesotrophic marine settings, three Synechococcus PhnD1 proteins also show a measurable affinity for MPn: CC9605_PhnD1 (17 ± 3 μM), WH8102_PhnD1 (41 ± 5 μM), and MITS9220_PhnD1 (47 ± 9 μM). While the binding affinity of these PhnD1 proteins for MPn is comparatively weaker than Pi and Pt, the measured binding constants are comparable to that known for the Prochlorococcus MIT9301 PhnD1 homologue (Pm_PhnD1, KD = 39–108 μM) (21). As organic phosphonates arise from cellular components, such as phospholipids, nucleic acids, amino acids, and polysaccharides, this broad substrate affinity observed for some Synechococcus isolates could potentially reflect an ability to access more labile forms of phosphorus [3, 30] rather than solely inorganic sources, which are generally sparingly soluble. As discussed earlier, none of the four Synechococcus strains under study (CC9311, CC9605, WH8102, MITS9220) harbours currently known putative Pt or MPn utilisation genes (ptxD, phnY/phnZ or C-P lyase) in their genome. Therefore, the affinity of Synechococcus PhnD1 proteins to Pt or MPn, analogous to the Prochlorococcus PhnD1 [3, 13], may be incidental, requiring further investigations into the structural mechanism of P selectivity for picocyanobacterial PhnD1 proteins.
The crystal structure of MITS9220_PhnD1 in complex with phosphate reveals an extensive hydrogen bond network
While crystal structures of PhnD1 in complex with Pt and MPn from Prochlorococcus are available, there are no crystal structures of picocyanobacterial PhnD1 protein in complex with Pi. Therefore, to understand the molecular mechanism of Pi selectivity, we determined a high-resolution crystal structure (2.0 Å) of Synechococcus MITS9220_PhnD1 with bound Pi, resulting in a closed ligand-bound complex (Fig. 3). Data collection and final refinement statistics for the crystal structure are outlined in Table 2. The overall structure of MITS9220_PhnD1 is similar to that of other class II substrate binding proteins (SBPs) [31] and specifically to cluster-F SBPs [32, 33], with two α/β domains separated by a longer central hinge region (8–10 amino acids). The extended hinge region in cluster-F SBPs is believed to provide greater flexibility between the ligand-bound (closed) and unbound (open) conformations of cluster-F SBPs [33]. This hinge region encompasses a buried ligand-binding cavity and includes a Pi molecule within the MITS9220_PhnD1 structure copurified with the protein. The MITS9220_PhnD1 structure displays an unusual asymmetric distribution of electrostatic surface potential (Fig. 3B), where the front is completely enveloped by positive charge, attracting several Cl- anions from the crystallisation buffer. While the presence of Cl- on the surface might be a crystallisation artifact, the strong positively charged front may have functional significance in facilitating picocyanobacterial PhnD1 association with the predominantly negatively charged phospholipid membrane surface.

A Block diagram representation (top) of the primary structure of MITS9220_PhnD1 showing the two lobes and the amino acids that determine the border of each domain. A cartoon representation (bottom-left) of the overall fold of MITS9220_PhnD1 highlighting the two α/β domains; lobe 1 (violet) and lobe 2 (pink), with Pi drawn as spheres (red). Also included in the representation are ten Cl− ions (green spheres) and 3 EDO (magenta sticks) molecules sequestered from the crystallisation condition. A detailed view of the ligand-binding pocket (bottom-right), outlining the interactions between Pi and MITS9220_PhnD1. The interacting sidechains and Pi are shown as sticks and coloured as per the respective domains, with the protein backbone shown as a cartoon. The single buried water is shown as a red sphere. B The electrostatic surface potential of the front (left) and back (right) of the MITS_9220 PhnD1 structure, with positive (blue) and negative charge (red) shown. C The protein sequence conservation mapping of picocyanobacterial PhnD1 homologues on MITS9220_PhnD1 structure (left) and ligand-binding pocket (right). The areas with high conservation (purple) and low conservation (green) are colour-coded.
Even though clear electron density is observed in the substrate binding site, which appears to have a tetrahedral geometry consistent with the Pi molecule in the binding pocket (Fig. 3), all ligands under study (Pi, Pt, and MPn) have similar chemical structures. Therefore, careful assignment of Pi to the positive difference Fourier electron density was ensured by considering van der Waals contacts and hydrogen bonds in the most appropriate chemical orientation and comparing refinements with the Pi replaced by Pt and MPn (Supplementary Fig. S8). Additionally, we also collected X-ray data above (2550 eV) and below (2400 eV) the sulphur edge, which assisted in dismissing the remote possibility of a sulphate ion instead being sequestered within the binding cavity (Supplementary Fig. S9).
Within the crystal structure of MITS9220_PhnD1, the four Pi oxygens are bound by a total of 10 direct hydrogen bonds, forming an extensive H-bond network to the main chain and sidechains of residues distributed between the two domains (Fig. 3A). These include a tyrosine (Y44) at the beginning of α2; an -STS- motif (S124, T125, S126) in a loop region joining β6 and α4; a histidine (H156) at the beginning of α6; an H-acceptor aspartic acid (D203) and a capping tyrosine residue (Y204) found just before β10. A single water molecule buried deep within the cavity also contributes to this hydrogen bond network. This water molecule is further stabilised by intramolecular hydrogen bonds to residues on domain 1 (T65) and domain 2 (S123), contributing to the interactions that connect the two domains and occluding the binding pocket from the solvent. Similarly, the capping Y204 sidechain is engaged in hydrogen bonds to residues D11 (domain 1) and N174 (domain 2), contributing to the two-domain interactions.
Sequence analysis shows strong conservation of all key MITS9220_PhnD1 Pi binding residues among most picocyanobacteria PhnD1 homologues (Fig. 3C), suggesting an analogous Pi binding mechanism in these isolates. The only exception is a substitution of aspartic acid (D203) to a functional homologue asparagine residue in the three Cyanobium strains, all Prochlorococcus LL1 strains (MIT0901, NatL1A, NatL2A, PAC1), and most Synechococcus clade I strains (Syn MV1R-18, Syn20, Pros 9-1, WH8016, WH8020) available on the Cyanorak database. In the absence of the biophysical data, it is unclear if the Asp to Asn substitution at the binding site impacts the P specificity of PhnD1 in these isolates, given the essential role of D203 as an H-bond acceptor.
MITS9220_PhnD1 structural comparison reveals the molecular basis of broad P specificity and medium Pi affinity
A search for structural homologues using the Dali server [34] reveals many structures similar to MITS9220_PhnD1 bound to Pi (Table 3). Among them, the structure of Pm_PhnD1 (56% sequence identity) in complex with Pt and MPn is most similar to MITS9220_PhnD1, with an r.m.s.d. of 1.1 Å. The structure of Pm_PhnD2 in complex with Pt, sequence identity of 30% to MITS9220_PhnD1, is the second closest structural homologue (r.m.s.d 1.7 Å). In comparison, the E. coli PhnD structure differs markedly with an r.m.s.d. of 2.4 Å, whereas the high-affinity Pi-binding PstS protein in E. coli aligns distantly with an r.m.s.d. of 5.1 Å. The overall fold of Synechococcus MITS9220_PhnD1 (with Pi) structure and the close structural homologues from Prochlorococcus MIT9301 [13] are very similar (Fig. 4A). The chemical structure of Pi, Pt, and MPn only differs at the R1 position substituted by OH, H and CH3, respectively. In all these structures, the R1 position of the respective P-ligand is capped by a tyrosine residue (Fig. 4B–D), leading to a smaller binding pocket for picocyanobacterial PhnD1 and PhnD2 proteins. In contrast, the E. coli PhnD (Ec_PhnD) crystal structure shows two H-bond acceptors, D205 and E177, in the active site, resulting in a comparatively larger binding cavity that can accommodate bulkier phosphonates, such as 2-aminoethylphosphonate (2-AEP) (Fig. 4E). The structural comparisons thus reveal the significance of the capping tyrosine residue as a steric barrier and rationalise why picocyanobacterial strains in the past were unable to bind and metabolise large complex Pn, such as 2-AEP [2, 23] but instead only use simple phosphonates such as MPn as a sole P source in strains encoding the potential Pn utilisation genes (phnY/phnZ) [23].

A The overlay of the crystal structure of Synechococcus MITS9220_PhnD1 in complex with Pi (blue; PDB 7S6G), Prochlorococcus MIT9301_PhnD1 (green; PDB 5LQ5) and MPn (purple; PDB 5LQ8), Prochlorococcus MIT9301_PhnD2 (yellow; PDB 5LV1). The ligand binding cavity (grey surface) with side-chain and ligand interactions is depicted for (B) Synechococcus MITS9220_PhnD1 in complex with Pi, (C) Prochlorococcus MIT9301_PhnD1 and MPn, (D) Prochlorococcus MIT9301_PhnD2, (E) E. coli PhnD in complex with 2-AEP (grey; PDB 3P7I) and (F) E. coli PstS in complex with Pi (pink; PDB 2ABH).
As discussed earlier, the Pi in MITS9220_PhnD1 structure is co-ordinated by ten hydrogen bonds (Fig. 4B), where the three oxygen atoms are engaged in multiple H-bonds, three each with O1, O3 and O4 in a trigonal geometry. The O2 atom (representing the R1 group) is, however, stabilised by a single H-bond formed by the capping residue Y204 (Supplementary Table S3). In contrast, the crystal structures of the high-affinity Pi-specific binding protein PstS, for example, in E. coli (PDB 2ABH), show an extensive hydrogen bond network [35, 36] consisting of 14 H-bonds (Fig. 4F). In addition to the three oxygen atoms O1, O3 and O4 engaging in three H-bonds each, the O2 atom in the case of E. coli PstS is stabilised by five H-bonds in a pentagonal geometry, explaining the high-affinity and specificity of PstS to Pi. The comparison of the binding pocket of the medium and high-affinity Pi binding proteins, MITS9220_PhnD1 and E. coli PstS, respectively, highlights the central role of the H-bond network in determining the specificity and binding affinity of the P-ligands. Our structural comparisons thus provide molecular insights into the potential fortuitous binding of Pt or MPn and the comparatively lower affinity of picocyanobacterial PhnD1 to Pi.
Low-affinity substrate-binding proteins are better suited for rapid substrate turnover, while high-affinity substrate-binding proteins are well suited for substrate scavenging at low concentrations. By having multiple nutrient transporters with different binding affinities and transport rates, cells can mitigate a “rate-affinity trade-off” and ensure the efficient transport of nutrients under different conditions [37]. In picocyanobacteria, the high-affinity Pst transporter could therefore be used to maintain a steady supply of Pi under phosphate-deplete conditions, while the comparatively low-affinity Phn transporter with potentially fast transport rates could be used to import large amounts of Pi when it is transiently available. For example, in times of upwelling, riverine inputs or Pi bursts from plankton or viral lysis. This would thus allow the picocyanobacterial cells to balance the need for rapid transport of nutrients under occasional conditions of plenty, with the need to efficiently utilise available P resources under typical phosphate-deplete conditions.
Synechococcus strains MITS9220 and WH8102 do not utilise Pt and MPn for growth as the sole P source
To test if Synechococcus strains not encoding phnD2 and the adjacent Pt/Pn utilisation genes can use alternative P sources other than Pi, we examined their growth in culture conditions containing either Pt or MPn as the sole P source. We specifically selected Synechococcus strain MITS9220 (clade CRD1a representative) associated with mesotrophic environments and strain WH8102 (clade III representative) found in oligotrophic waters, to understand growth physiology for isolates from environments with known differences in Pi availability.
We show both Synechococcus strains, MITS9220 and WH8102, when grown in the presence of either Pt or MPn as the sole P source, do not exhibit a clearly defined exponential growth (Fig. 5), as opposed to the growth seen in the presence of Pi. It is possible that the Synechococcus strains tested may have lost their ability to utilise Pt or MPn due to prolonged exposure to Pi-rich media in laboratory cultures. However, this scenario seems unlikely as the genomes of these strains lack currently known Pt or MPn metabolising genes (ptxD, phnY/phnZ or C-P lyase). A picocyanobacterial strain with these genes (e.g. Prochlorococcus MIT9301) is perfectly able to metabolise Pt and MPn as the sole P source to support its growth [2, 23], despite being in the laboratory cultures for an equivalent amount of time.

Synechococcus MITS9220 (A) and WH8102 (B) do not support well-defined exponential growth in the presence of Pt or MPn as a sole P source. The cultures were grown in PCR-S11 medium containing Pi (blue), Pt (orange), and MPn (green) in addition to the no P control (grey). The cell density was measured at regular intervals using a CytoFLEX S flow cytometer. The standard deviation of the mean cell density from triplicate cultures is represented as error bars.
For Synechococcus MITS9220, beyond day 3, all non-phosphate culture conditions exhibited a steady decline in cell density, reaching an undetectable cell density after day 15, upon which the experiment was terminated for this strain (Fig. 5A). In contrast, while WH8102 did not show exponential growth, it was able to persist for much longer, with detectable cell densities measured for all culture conditions for the entire duration of the experiment (49 days; Fig. 5B). It is unclear from our growth data if the observed persistence of WH8102 in P-deplete conditions is a result of the strain’s capability to persist in nutrient-deplete oligotrophic conditions or if they perhaps can support biotic (via unknown enzymatic pathways) or abiotic conversion of Pt or Pn to Pi. A recent study in E. coli demonstrated that in the absence of the two known canonical Pi transport-related genes that were sequentially deleted, the Phn uptake system could support growth with Pi as the sole P source [38].
Conclusions
All picocyanobacteria strains encode a well-conserved predicted ABC transporter, PhnDCE, that has long been thought to provide a competitive advantage in nutrient deplete conditions. We (for Synechococcus isolates) and others (for Prochlorococcus isolates) have successfully shown that the PhnD1 protein has a high-affinity for Pt and a medium-affinity for Pi and MPn. However, most picocyanobacterial strains lack known phosphite degradation or C-P lyase pathways to metabolise Pt or MPn. Our findings show that the PhnD1 expression and abundance in the global oceans is not influenced by the phosphate concentration. We further demonstrate the inability of several picocyanobacterial strains (including Synechococcus WH8102 and MITS9220 in this study) to grow on Pt or MPn as a sole P source. Taken together, these findings suggest that the PhnDCE system may function as a constitutive Pi transporter.
None of the Synechococcus PhnD1 proteins under investigation displayed a measurable affinity for hypophosphite, implying the requirement for minimally three covalently bound oxygen atoms in a trigonal arrangement to engage specific protein sidechains. Our structure of MITS9220_PhnD1 in complex with Pi shows an extensive H-bond network with Pi. However, they are fewer than the hydrogen bond interactions in the high-affinity PstS protein with Pi (e.g. in E. coli). This explains the lower measured affinity of picocyanobacterial PhnD1 proteins to Pi with a broader substrate range, which may be fortuitous.
We propose two potential scenarios that explain the critical role of PhnD1 in the environment. First, it is likely that PhnD1 aids in the recycling of phospholipid polar headgroups via a predicted phospholipase encoded by tesA, which is located adjacent to the phnDCE genes. TesA could potentially hydrolyse the commonly found picocyanobacterial phospholipid (PG) to diacylglycerol and glycerol-3-phosphate. Subsequently, the periplasmic alkaline phosphatase would release Pi, which can then be taken up by the PhnD1CE (or Pst) transporter, providing a mechanism for recycling P from the phospholipid bilayer. The second scenario is based on the rate-affinity trade-off that cells mitigate by having multiple transporters for the same nutrient. A medium-affinity Pi transporter such as the PhnD1CE, with potentially fast transport rates, could help picocyanobacterial cells to acquire large amounts of Pi when it is transiently available. This would enable picocyanobacterial cells to balance their requirement for quick transport during sporadic abundance with efficient utilisation of P resources (via Pst transport system) under typical phosphate-deplete conditions.
Materials and methods
Genomic and Phylogenetic analyses
Phylogenetic analysis of gene cluster CK_860 annotated as PhnD1 (phosphate/phosphonate-binding proteins) within the Cyanorak database (www.sb-roscoff.fr/cyanorak) [6, 20], was performed using a modified method given by Wilding et al. [39]. Briefly, the 97 orthologous PhnD1 sequences, as well as PhnD2 (CK_6203, 10 sequences) and PhnD3 (CK_56876, 1 sequence), were used to compute a multiple sequence alignment using the L-iNS-I option of MAFFT [40]. The phylogenetic tree was inferred using IQ-Tree [41], using the -TESTONLY option, found to be WAG + G4. The final phylogenetic tree was generated from the inferred and visualised using iTOL [42].
The geographic distribution and expression of PhnD1 and PhnD2 homologues in marine picocyanobacterial populations were analysed using the metagenomes and metatranscriptomes available from the Ocean Microbial Reference Gene Catalogue (OM-RGC) [21, 22]. The protein sequence of Prochlorococcus MIT9301_PhnD1 and MIT9301_PhnD2 were used to search against the ‘OM_RGC_v2_metaG’ and ‘OM_RGC_v2_metaT’ catalogue using the default parameters. To limit our analyses to picocyanobacterial sequences, we chose a high significance E-threshold (>e−75) and ensured that the homologue matches (with sequence identity >70%) for the PhnD1 and PhnD2 sequences did not overlap. Plots were viewed using P as the environmental variable of interest. Additionally, for comparison, we provide environmental abundance maps for Prochlorococcus and Synechococcus lineage-specific genes. The protein sequence of a predicted chlorophyll b synthase, PcCAO (CK_2331), specific to Prochlorococcus strains and a predicted phycocyanin lyase, CpcS (CK_1523), specific to Synechococcus strains were used to search against the ‘OM_RGC_v2_metaG’ catalogue using the default parameters with a high significance E-threshold (>e−75).
Recombinant expression and protein purification
The protein sequences of CC9311_01670, CC9605_01294, MITS9220_01173, and WH8102_01170, annotated to encode PhnD1, were analysed using the SignalP 4.0 [43].
Following truncation of the N-terminal signal peptide (Supplementary Table S3), the genes were PCR-amplified from respective genomic DNA (extracted using the CTAB/phenol-chloroform method) incorporating vector-specific (pOPINF) [44] overhang regions for heterologous expression in E. coli. Ligation-independent cloning (Clontech) [45] into the pOPINF vector was carried out using KpnI and HindIII restriction sites to incorporate an N-terminal hexahistidine tag with a 3C protease cleavage site.
Following plasmid transformation in E. coli, all target proteins were expressed to high density using the autoinduction method [46] in a 1 L culture medium. While CC9311_PhnD1, MITS9220_PhnD1, and WH8102_PhnD1 proteins displayed the highest expression in an E. coli Lemo strain (BL21 derivative, NE Biolabs), CC9605_PhnD1 achieved higher expression in BL21 derivative strain (Rosetta 2, Novagen). All four PhnD1 proteins were purified using IMAC, as described earlier [47]. Protein-containing fractions were pooled and desalted using size exclusion chromatography (SEC) with a Superdex HiLoad 200 16/600 column (GE Healthcare) equilibrated in buffer containing HEPES (50 mM, pH 7.4), NaCl (300 mM), and glycerol (5% v/v). All buffers contained the reducing agent tris(2-carboxyethyl)phosphine (TCEP, 0.5 mM).
For CC9605_PhnD1, MITS9220_PhnD1, and WH8102_PhnD1 proteins, SEC traces show a single, well-defined peak corresponding to a monomeric state at expected molecular size (~30 kDa). However, for the CC9311_PhnD1 protein, multiple peaks were observed during SEC. Isolation of pure CC9311_PhnD1 occurred by fractionating the peak corresponding to the monomeric species, allowing >90% purity.
PhnD1 ligand screening and determination of binding affinity to P sources
Ligand screening was performed using differential scanning fluorimetry (DSF) [48] with SYPRO Orange dye (Invitrogen) used to monitor fluorescence at 590 nm following excitation at 485 nm. DSF measures the thermal stability of the protein-ligand complexes, whereby the temperature at which the protein denatures (melting temperature TM) is shifted to a higher temperature in the presence of a stabilising ligand. The compounds that stabilise the protein the most can then be identified as putative ligands for further binding affinity analysis. Compounds tested incorporated cocktails of common protein stabilising ligands (HR2-096 Silver Bullets, Hampton Research) and single-molecule P sources: phosphate, phosphite, hypophosphite, and methyl phosphonate (each as sodium salts). Each condition was tested in triplicate, with each plate containing a control well with no additive. Thermal melt curves were analysed using the analysis template provided [48] and fitting of Boltzmann distribution (GraphPad Prism) to determine mid-point thermal melt temperatures. Cocktail conditions leading to ≥5 °C in melting temperature (TM) were considered a significant increase and repeated in triplicate. The thermal melting curve for all four PhnD proteins, CC9311_PhnD, CC9605_PhnD, MITS9220_PhnD and WH8102_PhnD, showed a single transition point with a melting temperature (TM) of 47 °C, 32 °C, 46 °C, and 35 °C, respectively.
To elucidate the affinity of PhnD1 to P, the DSF assay was modified and carried out with increasing ligand concentrations as previously described [47], leading to an incremental shift in observed melting temperatures. Data were processed using the provided Python package [29]. Results were plotted and checked for consistency by independently determining EC50 values (GraphPad Prism). The standard error of derived KD values was determined by comparing the deviation in fitting three individual replicates. It was, in all cases, smaller than 10% of the determined affinity measurement.
MITS9220_PhnD1 crystallisation and structure determination
All four Synechococcus PhnD1 purified protein products were subjected to sparse matrix crystallisation screening at 20 °C by sitting drop vapour diffusion method by mixing 0.1 μL of protein (10 mg/mL) with 0.1 μL of the reservoir. However, only MITS9220_PhnD1 showed good diffraction-quality crystals grown in 0.1 M Tris pH 8.5, 25% PEG 3350. Before plunge freezing in liquid nitrogen, MITS9220_PhnD1 crystals were cryo-protected in the mother liquor with an additional 30% (v/v) ethylene glycol. The crystals were further transferred into the vacuum vessel using an adapted cryotransfer system (Leica VCT100).
The data were collected at the Diamond Light Source I23 beamline [49], equipped with the semi-cylindrical Pilatus 12 M (Dectris AG, Switzerland) detector, at four different wavelengths, 2.7552, 3.0996, 4.8621 and 5.1666 Å. Data were processed using XDS [50] and merged using XSCALE. The structure of MITS9220_PhnD1 was solved by experimental phasing using the SAD dataset collected at 3.0996 Å wavelength using the CRANK2 pipeline [51]. After density modification, automated model building within the CRANK2 pipeline produced an initial model comprising a single copy. Several rounds of manual model building and refinement were carried out in COOT [52]and Refmac5 [53] before validating the final model with Molprobity [54]. The final coordinates contain residues 1–270 of the SignalP truncated mature sequence in one chain, with one defined phosphate site, nine chloride anions and three EDO molecules. Final refinement statistics are given in Table 2, and coordinates are deposited in the PDB (7S6G). Anomalous difference Fourier from long-wavelength data was generated using ANODE [55]. The positions of anomalous peaks higher than 4.0σ as output by ANODE from datasets both above (2.755 Å) and below the Ca-edge (3.0996 Å) were inspected in COOT [52] to discount these peaks as calcium. The peaks at 3.0996 Å combined with their absence at 4.8621 Å are indicative of chloride ions. To ascertain whether a phosphate or sulphate was present in the ligand site, anomalous difference Fourier maps were compared from data collected above (4.8621 Å) and below (5.1666 Å) the sulphur absorption edge. The strong peak present in both datasets suggests that the ligand contains phosphorus (Supplementary Fig. S8).
Synechococcus MITS9220 and WH8102 growth in the presence of alternate P sources
Synechococcus MITS9220 and WH8102 cultures were grown at 22 °C, with 40 µM photons m−2 s−1 continuous illumination, in an orbital shaker at 100 rpm in acid-washed polycarbonate flasks using Red Sea salt-based PCR-S11 medium [56]. For treatment conditions containing alternative P compounds, 50 µM NaH2PO4 was replaced with 50 µM phosphite (Na2HPO3·5H2O) or methylphosphonate. In P-deplete media, an equal volume of water was added instead of a P compound. Experimental cultures were started from mid-logarithmic cultures grown in phosphate-containing PCR-S11 media. A small inoculum (~100 µL) was used to minimise the transfer of phosphate from stock cultures. Four biological replicates were tested for each experimental condition. Growth was regularly monitored via cell counts using a CytoFLEX S flow cytometer. Cells were identified by chlorophyll and phycoerythrin fluorescence the following excitation using a blue laser (488 nm). Phosphate concentration in all treatment conditions was monitored regularly for the duration of the experiments using a Phosphate Colorimetric kit following the manufacturer’s instructions (Sigma-Aldrich) with a detection limit of 0.5 µM.
Data availability
The structure coordinates for MITS9220_PhnD1 are deposited in the Protein Data Bank (accession code 7S6G). All other study data is included in the article and/or supporting information files.
References
White A, Dyhrman S. The marine phosphorus cycle. Front Microbiol. 2013;4:105.
Martinez A, Osburne MS, Sharma AK, DeLong EF, Chisholm SW. Phosphite utilization by the marine picocyanobacterium Prochlorococcus MIT9301. Environ Microbiol. 2012;14:1363–77.
Feingersch R, Philosof A, Mejuch T, Glaser F, Alalouf O, Shoham Y, et al. Potential for phosphite and phosphonate utilization by Prochlorococcus. ISME J. 2012;6:827–34.
Moore LR, Ostrowski M, Scanlan DJ, Feren K, Sweetsir T. Ecotypic variation in phosphorus-acquisition mechanisms within marine picocyanobacteria. Aquat Micro Ecol. 2005;39:257–69.
Scanlan DJ, Ostrowski M, Mazard S, Dufresne A, Garczarek L, Hess WR, et al. Ecological genomics of marine picocyanobacteria. Microbiol Mol Biol Rev. 2009;73:249–99.
Doré H, Farrant GK, Guyet U, Haguait J, Humily F, Ratin M, et al. Evolutionary mechanisms of long-term genome diversification associated with niche partitioning in marine picocyanobacteria. Front Microbiol. 2020;11:567431.
Tetu SG, Brahamsha B, Johnson DA, Tai V, Phillippy K, Palenik B, et al. Microarray analysis of phosphate regulation in the marine cyanobacterium Synechococcus sp. WH8102. ISME J. 2009;3:835–49.
Santos-Beneit F. The Pho regulon: a huge regulatory network in bacteria. Front Microbiol. 2015;6:402.
Wanner BL, Chang BD. The phoBR operon in Escherichia coli K-12. J Bacteriol. 1987;169:5569–74.
Gebhard S, Ekanayaka N, Cook GM. The low-affinity phosphate transporter PitA is dispensable for in vitro growth of Mycobacterium smegmatis. BMC Microbiol. 2009;9:254.
Ilikchyan IN, McKay RM, Zehr JP, Dyhrman ST, Bullerjahn GS. Detection and expression of the phosphonate transporter gene phnD in marine and freshwater picocyanobacteria. Environ Microbiol. 2009;11:1314–24.
Martiny AC, Coleman ML, Chisholm SW. Phosphate acquisition genes in Prochlorococcus ecotypes: evidence for genome-wide adaptation. Proc Natl Acad Sci USA. 2006;103:12552–7.
Bisson C, Adams NBP, Stevenson B, Brindley AA, Polyviou D, Bibby TS, et al. The molecular basis of phosphite and hypophosphite recognition by ABC-transporters. Nat Commun. 2017;8:1746.
Zubkov MV, Mary I, Woodward EM, Warwick PE, Fuchs BM, Scanlan DJ, et al. Microbial control of phosphate in the nutrient-depleted North Atlantic subtropical gyre. Environ Microbiol. 2007;9:2079–89.
Zwirglmaier K, Jardillier L, Ostrowski M, Mazard S, Garczarek L, Vaulot D, et al. Global phylogeography of marine Synechococcus and Prochlorococcus reveals a distinct partitioning of lineages among oceanic biomes. Environ Microbiol. 2008;10:147–61.
Zwirglmaier K, Heywood JL, Chamberlain K, Woodward EM, Zubkov MV, Scanlan DJ. Basin-scale distribution patterns of picocyanobacterial lineages in the Atlantic Ocean. Environ Microbiol. 2007;9:1278–90.
Ahlgren NA, Rocap G. Culture isolation and culture-independent clone libraries reveal new marine Synechococcus ecotypes with distinctive light and N physiologies. Appl Environ Microbiol. 2006;72:7193–204.
Fuller NJ, Marie D, Partensky F, Vaulot D, Post AF, Scanlan DJ. Clade-specific 16S ribosomal DNA oligonucleotides reveal the predominance of a single marine Synechococcus clade throughout a stratified water column in the Red Sea. Appl Environ Microbiol. 2003;69:2430–43.
Sohm JA, Ahlgren NA, Thomson ZJ, Williams C, Moffett JW, Saito MA, et al. Co-occurring Synechococcus ecotypes occupy four major oceanic regimes defined by temperature, macronutrients and iron. ISME J. 2016;10:333–45.
Garczarek L, Guyet U, Dore H, Farrant GK, Hoebeke M, Brillet-Gueguen L, et al. Cyanorak v2.1: a scalable information system dedicated to the visualization and expert curation of marine and brackish picocyanobacteria genomes. Nucleic Acids Res. 2021;49:D667–D76.
Villar E, Vannier T, Vernette C, Lescot M, Cuenca M, Alexandre A, et al. The Ocean Gene Atlas: Exploring the biogeography of plankton genes online. Nucleic Acids Res. 2018;46:W289–95.
Sunagawa S, Coelho LP, Chaffron S, Kultima JR, Labadie K, Salazar G, et al. Ocean plankton. Struct Funct Glob Ocean Microbiome Sci. 2015;348:1261359.
Sosa OA, Casey JR, Karl DM. Methylphosphonate oxidation in Prochlorococcus strain MIT9301 supports phosphate acquisition, formate excretion, and carbon assimilation into purines. Appl Environ Microbiol. 2019;85:e00289-19.
Martinez A, Tyson GW, Delong EF. Widespread known and novel phosphonate utilization pathways in marine bacteria revealed by functional screening and metagenomic analyses. Environ Microbiol. 2010;12:222–38.
Kovacic F, Granzin J, Wilhelm S, Kojic-Prodic B, Batra-Safferling R, Jaeger KE. Structural and functional characterisation of TesA - a novel lysophospholipase A from Pseudomonas aeruginosa. PLoS One. 2013;8:e69125.
Van Mooy BA, Fredricks HF, Pedler BE, Dyhrman ST, Karl DM, Koblizek M, et al. Phytoplankton in the ocean use non-phosphorus lipids in response to phosphorus scarcity. Nature. 2009;458:69–72.
Simockova M, Holic R, Tahotna D, Patton-Vogt J, Griac P. Yeast Pgc1p (YPL206c) controls the amount of phosphatidylglycerol via a phospholipase C-type degradation mechanism. J Biol Chem. 2008;283:17107–15.
Niebling S, Burastero O, Burgi J, Gunther C, Defelipe LA, Sander S, et al. FoldAffinity: binding affinities from nDSF experiments. Sci Rep. 2021;11:9572.
Bai N, Roder H, Dickson A, Karanicolas J. Isothermal analysis of ThermoFluor data can readily provide quantitative binding affinities. Sci Rep. 2019;9:2650.
Gomez-Garcia MR, Davison M, Blain-Hartnung M, Grossman AR, Bhaya D. Alternative pathways for phosphonate metabolism in thermophilic cyanobacteria from microbial mats. ISME J. 2011;5:141–9.
Fukami-Kobayashi K, Tateno Y, Nishikawa K. Domain dislocation: a change of core structure in periplasmic binding proteins in their evolutionary history. J Mol Biol. 1999;286:279–90.
Scheepers GH, Lycklama ANJA, Poolman B. An updated structural classification of substrate-binding proteins. FEBS Lett. 2016;590:4393–401.
Berntsson RP, Smits SH, Schmitt L, Slotboom DJ, Poolman B. A structural classification of substrate-binding proteins. FEBS Lett. 2010;584:2606–17.
Holm L. Dali server: structural unification of protein families. Nucleic Acids Res. 2022;50:W210–5.
Yao N, Ledvina PS, Choudhary A, Quiocho FA. Modulation of a salt link does not affect binding of phosphate to its specific active transport receptor. Biochemistry. 1996;35:2079–85.
Luecke H, Quiocho FA. High specificity of a phosphate transport protein determined by hydrogen bonds. Nature. 1990;347:402–6.
Montano-Gutierrez LF, Correia K, Swain PS. Multiple nutrient transporters enable cells to mitigate a rate-affinity tradeoff. PLoS Comput Biol. 2022;18:e1010060.
Stasi R, Neves HI, Spira B. Phosphate uptake by the phosphonate transport system PhnCDE. BMC Microbiol. 2019;19:79.
Wilding M, Peat TS, Kalyaanamoorthy S, Newman J, Scott C, Jermiin LS. Reverse engineering: transaminase biocatalyst development using ancestral sequence reconstruction. Green Chem. 2017;19:5375–80.
Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30:772–80.
Nguyen LT, Schmidt HA, von Haeseler A, Minh BQ. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol. 2015;32:268–74.
Letunic I, Bork P. Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47:W256–W9.
Petersen TN, Brunak S, von Heijne G, Nielsen H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods. 2011;8:785–6.
Berrow NS, Alderton D, Sainsbury S, Nettleship J, Assenberg R, Rahman N, et al. A versatile ligation-independent cloning method suitable for high-throughput expression screening applications. Nucleic Acids Res. 2007;35:e45.
Aslanidis C, de Jong PJ. Ligation-independent cloning of PCR products (LIC-PCR). Nucleic Acids Res. 1990;18:6069–74.
Studier FW. Protein production by auto-induction in high density shaking cultures. Protein Expr Purif. 2005;41:207–34.
Ford BA, Ranjit P, Mabbutt BC, Paulsen IT, Shah BS. ProX from marine Synechococcus spp. show a sole preference for glycine-betaine with differential affinity between ecotypes. Environ Microbiol. 2022;24:6071–85.
Niesen FH, Berglund H, Vedadi M. The use of differential scanning fluorimetry to detect ligand interactions that promote protein stability. Nat Protoc. 2007;2:2212–21.
Wagner A, Duman R, Henderson K, Mykhaylyk V. In-vacuum long-wavelength macromolecular crystallography. Acta Crystallogr D Struct Biol. 2016;72:430–9.
Kabsch W. XDS. Acta Crystallogr D Biol Crystallogr. 2010;66:125–32.
Pannu NS, Waterreus WJ, Skubak P, Sikharulidze I, Abrahams JP, de Graaff RA. Recent advances in the CRANK software suite for experimental phasing. Acta Crystallogr D Biol Crystallogr. 2011;67:331–7.
Emsley P, Cowtan K. Coot: model-building tools for molecular graphics. Acta Crystallogr D Biol Crystallogr. 2004;60:2126–32.
Murshudov GN, Skubak P, Lebedev AA, Pannu NS, Steiner RA, Nicholls RA, et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr D Biol Crystallogr. 2011;67:355–67.
Chen VB, Arendall WB 3rd, Headd JJ, Keedy DA, Immormino RM, Kapral GJ, et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr D Biol Crystallogr. 2010;66:12–21.
Thorn A, Sheldrick GM. ANODE: anomalous and heavy-atom density calculation. J Appl Crystallogr. 2011;44:1285–7.
Rippka R, Coursin T, Hess W, Lichtle C, Scanlan DJ, Palinska KA, et al. Prochlorococcus marinus Chisholm et al. 1992 subsp. pastoris subsp. nov. strain PCC 9511, the first axenic chlorophyll a2/b2-containing cyanobacterium (Oxyphotobacteria). Int J Syst Evol Microbiol. 2000;50:1833–47.
Sunagawa S, Acinas SG, Bork P, Bowler C, Acinas SG, Babin M, et al. Tara Oceans: towards global ocean ecosystems biology. Nat Rev Microbiol 2020;18:428–45.
Belisle BS, Avila Paz AA, Carpenter AR, Cormier TC, Lewis AJ, Menin LS, et al. Genome sequences of Synechococcus sp. Strain MITS9220 and cocultured cyanophage SynMITS9220M01. Microbiol Resour Announc. 2020;9:e00481-20.
Ahlgren NA, Belisle BS, Lee MD. Genomic mosaicism underlies the adaptation of marine Synechococcus ecotypes to distinct oceanic iron niches. Environ Microbiol. 2020;22:1801–15.
Rocap G, Distel DL, Waterbury JB, Chisholm SW. Resolution of Prochlorococcus and Synechococcus ecotypes by using 16S-23S ribosomal DNA internal transcribed spacer sequences. Appl Environ Microbiol. 2002;68:1180–91.
Rizk SS, Cuneo MJ, Hellinga HW. Identification of cognate ligands for the Escherichia coli phnD protein product and engineering of a reagentless fluorescent biosensor for phosphonates. Protein Sci. 2006;15:1745–51.
Alicea I, Marvin JS, Miklos AE, Ellington AD, Looger LL, Schreiter ER. Structure of the Escherichia coli phosphonate binding protein PhnD and rationally optimized phosphonate biosensors. J Mol Biol. 2011;414:356–69.
Poddar N, Badilla C, Maghool S, Osborne TH, Santini JM, Maher MJ. Structural and functional investigation of the periplasmic Arsenate-binding protein ArrX from Chrysiogenes arsenatis. Biochemistry 2021;60:465–76.
Acknowledgements
This research was supported, in part, by the Australian Research Council Discovery grant (DP200102944) and the Australian Research Council Centre of Excellence in Synthetic Biology (CE200100029) to ITP; BAF acknowledges receipt of an Australian Government Research Training Program (RTP) Scholarship. The authors thank Dr Lisa Moore and Dr Martin Ostrowski for insightful discussions during the course of the research project.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Contributions
BSS and ITP designed the experiments and directed the study. BSS, BAF and DV performed laboratory work. BSS and HM performed crystallisation; BSS, HM, CO, and VM collected x-ray data sets and determined protein structure. RJO contributed to the experimental workflows. BSS analysed data; BSS, BAF and ITP wrote the paper. ITP contributed to the funding. All authors contributed to the revisions.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shah, B.S., Ford, B.A., Varkey, D. et al. Marine picocyanobacterial PhnD1 shows specificity for various phosphorus sources but likely represents a constitutive inorganic phosphate transporter. ISME J 17, 1040–1051 (2023). https://doi.org/10.1038/s41396-023-01417-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-023-01417-w
