
Abstract
Hematopoietic stem cell transplant (HSCT) treats or cures a variety of hematological and inherited disorders. Unfortunately, patients who undergo HSCT are susceptible to infections by a wide array of opportunistic pathogens. Pseudomonas aeruginosa bacteria can have life-threatening effects in HSCT patients by causing lung pathology that has been linked to high levels of the potent pro-inflammatory cytokine, interleukin-1β (IL-1β). Using a murine bone marrow transplant (BMT) model, we show that overexpression of prostaglandin E2 (PGE2) post-BMT signals via EP2 or EP4 to induce cyclic adenosine monophosphate (cAMP), which activates protein kinase A or the exchange protein activated by cAMP (Epac) to induce cAMP response element binding–dependent transcription of IL-1β leading to exacerbated lung injury in BMT mice. Induction of IL-1β by PGE2 is time and dose dependent. Interestingly, IL-1β processing post-P. aeruginosa infection occurs via the enzymatic activity of either caspase-1 or caspase-8. Furthermore, PGE2 can limit autophagy-mediated killing of P. aeruginosa in alveolar macrophages, yet autophagy does not have a role in PGE2-mediated upregulation of IL-1β. Reducing PGE2 levels with indomethacin improved bacterial clearance and reduced IL-1β-mediated acute lung injury in P. aeruginosa–infected BMT mice.
Similar content being viewed by others

Introduction
Hematopoietic stem cell transplant (HSCT) utilizes stem cells derived from bone marrow, umbilical cord blood, or peripheral blood to treat or cure a variety of hematological disorders.1 This procedure has become a standard of care with >18,000 HSCT performed every year in the United States alone.1 HSCT requires a conditioning regimen (e.g., total body irradiation and/or chemotherapy) to allow the patient’s hematopoietic system to be replaced by donor cells. Depending on disease, the patient can undergo autologous (self-donation of stem cells) or allogeneic (stem cells from a human leukocyte antigen–matched donor) transplants. Unfortunately, patients who undergo HSCT become immunosuppressed and susceptible to infections by opportunistic pathogens.2 Around 60% of HSCT patients develop pulmonary complications, many of them due to bacterial infections, which correlate with high mortality and morbidity.1, 2, 3 Infections post-HSCT are independent of conditioning regimen and type of transplant as both autologous and allogenic patients are highly susceptible. Infections can take place in the preengraftment phase (before 30 days posttransplant) as well as after immune reconstitution (after 30 days posttransplant).3 Although infections can be caused by viral, bacterial, or fungal organisms, bacterial infections are increasingly problematic due to the rise in drug-resistant bacteria. Bacterial infections post-HSCT can occur in 51.3% of HSCT patients.2 These infections, including infections by the Gram-negative bacterial pathogen, Pseudomonas aeruginosa, cause life-threatening complications.2
P. aeruginosa is an opportunistic pathogen that normally infects immunocompromised individuals, such as HSCT patients.2, 4, 5, 6, 7 It is a leading nosocomial pathogen, and it is the most frequently isolated Gram-negative bacteria in the intensive care unit.8 This pathogen causes urinary tract infection, hospital-acquired pneumonia, and bacteremia in burn patients.9 It is also the predominant cause of morbidity and mortality in cystic fibrosis patients. There has been a significant increase in research effort studying P. aeruginosa due to the difficulty of treating infected patients, as 26% of its isolates are resistant to antibiotics and disinfectants.8 Understanding how to modulate P. aeruginosa infections in an antibiotic-independent method is likely to have positive impacts on the outcome of infected patients. Although many antimicrobial pathways have been linked to P. aeruginosa clearance, induction of inflammasome activation and interleukin-1β (IL-1β) secretion have pathogenic roles during P. aeruginosa infection.5
The inflammasome is a multi-protein complex expressed mainly in immune cells and activated by pathogenic stimuli. Its activation leads to secretion of two potent inflammatory cytokines, IL-1β and IL-18. Their secretion pathways are complex and partially unknown but have been well established to require two signals. Signal one leads to upregulation of immature pro-cytokines and is mediated by Toll-like receptor stimulation by pathogen-associated molecular patterns. Gram-negative bacteria such as P. aeruginosa can trigger signal one by stimulation of Toll-like receptors 4 and 5 via lipopolysaccharide (LPS) and flagella, respectively.10, 11, 12 Signal two can be triggered by a wide array of pathogenic stimuli that culminate in aggregation of inflammasome components (e.g., caspase-1 or NOD-like receptor proteins) and result in secretion of mature IL-1β and IL-18. P. aeruginosa can induce signal two via recognition of type III secretion proteins. Although a protective mechanism against many pathogens, overproduction of IL-1β has been associated with auto-inflammatory syndromes such as gout and periodic fever syndromes, such as Familial Mediterranean Fever and cryopyrin-associated periodic fever syndromes.13 Therapies targeting IL-1β signaling have shown better outcomes in cryopyrin-associated periodic fever syndrome patients.14 Moreover, asbestos and silica inhalation can cause IL-1β-dependent pulmonary fibrosis mediated by alveolar macrophage (AMs).15 In the lung, P. aeruginosa can induce IL-1β secretion by AMs.5 Interestingly, depleting AMs prior to P. aeruginosa infection leads to significantly lower levels of IL-1β in the lung, improving survival.5 Apart from IL-1β secretion, P. aeruginosa infection has also been shown to be regulated by prostaglandin E2 (PGE2).6, 7, 16
PGE2 is a lipid mediator derived from arachidonic acid by the enzymatic activity of cyclooxygenase (COX) and PGE synthases that signals through four different G-protein-coupled plasma membrane receptors of the E-prostnoid (EP) family termed EP1, EP2, EP3, and EP4. Each receptor activates different intracellular pathways. Stimulation of EP1 receptor increases intracellular calcium and activation of protein kinase C, which stimulates the transcription factors, nuclear factor of activated T cells and nuclear factor-κB.17 EP2 and EP4 receptors are stimulators of adenylyl cyclase (AC) and phosphoinositide 3-kinase, respectively. AC mediates conversion of adenosine triphosphate (ATP) to cyclic adenosine monophosphate (cAMP) leading to activation of protein kinase A (PKA) and the transcription factor cAMP response element binding (CREB). The EP3 receptor is a regulator of the EP2–EP4 signaling pathway as its activation leads to inhibition of AC. P. aeruginosa infection increases the levels of inducible COX2 leading to high levels of PGE2 and inhibition of COX2, with subsequent diminished production of PGE2, can lead to a better outcome in P. aeruginosa–infected mice.7 Interestingly, HSCT patients possess higher levels of PGE2 in blood and bronchoalveolar lavage (BAL) when compared with healthy non-transplanted individuals.18, 19
PGE2 is dysregulated in HSCT patients18 and suppression of PGE2 confers protection against P. aeruginosa infection in murine HSCT models.6 However, other roles of PGE2 in the context of HSCT and pulmonary P. aeruginosa infection remain elusive. To study bacterial lung infections post-HSCT, we use a mouse model of syngeneic bone marrow transplant (BMT). Total body irradiation is used as a conditioning regimen.6, 20, 21, 22 Experiments are performed in the postengraftment period, 5 weeks post-BMT when lung leukocytes are composed of 82% donor cells and splenic leukocytes are 95% donor cells.20 We previously reported that BMT mice are deficient in phagocytosis and killing of P. aeruginosa,6, 23 correlating with observations in humans.2 In the present study, we compare the cytokine profile and lung tissue injury of control and BMT mice after P. aeruginosa infection. We also tested the direct relationship of PGE2 with P. aeruginosa–induced IL-1β and examined the effect that PGE2 stimulation had on autophagy, a main mechanism of P. aeruginosa clearance by AMs that has also been linked to inflammasome regulation.24, 25
Results
BMT mice are deficient in clearing P. aeruginosa infection and experience exacerbated lung tissue injury
BMT mice are deficient in clearing a P. aeruginosa PAO-1 infection compared with healthy control mice 24 h postinfection (Figure 1a), confirming published data.6 Deficiency in clearing bacteria is not due to low numbers of immune cells in the alveolar compartment as there are no differences in cell numbers or percentages of monocyte/macrophages and lymphocytes in the BAL (see Supplementary Figure S1A,B online) between groups. We also noted higher levels of albumin in the BAL fluid (BALF) of BMT mice compared with control mice (Figure 1b), suggesting more severe pulmonary injury (Figure 1c). Acute lung injury (ALI) is a leading cause of death in the intensive care unit, characterized by accumulation of leukocytes, protein leakage, and epithelial injury. ALI has been linked to high levels of IL-1β in the lung.26 Thus we measured the levels of several pro-inflammatory cytokines. We found higher levels of IL-1β but not other pro-inflammatory cytokines (IL-6, IL-12, tumor necrosis factor (TNF)-α; data not shown) in BALF from infected BMT mice compared with control mice (Figure 2a). We tested the levels of PGE2 in BALF and detected higher levels of PGE2 in BMT mice infected with PA01 compared with infected control mice. Moreover, we noticed that mice expressing higher levels of PGE2 have direct and significant correlations between levels of IL-1β and tissue injury but not with other cytokines, such as TNF-α (Figure 2c–f). Previous research has suggested a role for PGE2 in IL-1β induction.27, 28 Thus exacerbated levels of IL-1β post-P. aeruginosa infection might mediate lung tissue injury and be dependent on PGE2. Additionally, P. aeruginosa–mediated IL-1β release has been shown to be dependent on AMs.5, 26 Thus we directed our attention to AMs in BMT mice.

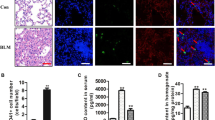
Bone marrow transplant (BMT) mice are deficient in clearing Pseudomonas aeruginosa infection and experience exacerbated lung tissue injury. (a) Colony-forming units (CFUs) were counted 24 h after infecting non-transplanted control and BMT mice (C57BL/6J→C57BL/6J) with 5 × 105 CFUs of P. aeruginosa (PA01). (n=5 control; n=5 BMT). (b) Albumin measurements from bronchoalveolar lavage fluid (BALF) from PA01-infected control and BMT mice. (n=7 control; n=6 BMT) (c) Hematoxylin and eosin stain of lungs from saline or PA01-infected control and BMT mice; images taken at × 40 magnification (representative of n=3 control; n=3 BMT). Statistics are Student’s t-test between comparative groups. *P<0.05. Data are representative of at least two independent experiments.

Pseudomonas aeruginosa infection induces higher levels of interleukin (IL)-1β and prostaglandin E2 (PGE2) post–bone marrow transplant (post-BMT) correlating with increased lung injury. (a) IL-1β and (b) PGE2 measurements from the bronchoalveolar lavage fluid (BALF) of control and BMT mice 24 h after infection with PA01; measurements were carried out by enzyme-linked immunosorbent assay (n=7 control; n=6 BMT); (c) correlation between albumin and IL-1β in BALF 24 h post-PAO1; (d) correlation between albumin and PGE2 in BALF 24 h post-PAO1; (e) correlation between PGE2 and IL-1β in BALF 24 h post-PAO1; (f) correlation between PGE2 and tumor necrosis factor (TNF)-α in BALF 24 h post-PAO1. In all correlations, closed symbols represent control mice, whereas open circles are BMT mice. R=Pearson correlation coefficient; *P<0.05.
Pro-inflammatory AMs in BMT mice account for higher levels of IL-1β in response to P. aeruginosa infection
To determine the sources of IL-1β and PGE2 in the lung, we compared mRNA levels from AMs obtained by BALF to lung interstitium. Transcripts for the rate-limiting enzymes in PGE2 synthesis, COX1 and COX2, were higher in AMs from BMT mice compared with control (Figure 3a). This was unique to AMs but not to interstitial samples. In addition, AMs from BMT mice have higher levels of IL-1β transcripts compared with control cells (Figure 3a). Consistent with our past studies, overnight culture of AMs from BMT mice secreted higher levels of PGE2 compared with cells from control mice (Figure 3b). To assess whether PGE2 can have an effect on IL-1β secretion, we pretreated AMs with or without PGE2 prior to PA01 infection, in vitro. We detected higher levels of IL-1β in supernatants of AMs pretreated with PGE2 and infected with PAO1 compared with AMs infected with PA01 alone (Figure 3c). Moreover, to determine whether higher bacterial burden in BMT mice was responsible for higher levels of IL-1β, we induced ALI with LPS instillation in control and BMT mice. We detected higher IL-1β but not IL-6 or TNF-α in BALF from BMT mice compared with control mice post-LPS instillation (Figure 3d–f).

Alveolar macrophages (AMs) in bone marrow transplant (BMT) mice account for higher interleukin (IL)-1β release post-P. aeruginosa in response to prostaglandin E2 (PGE2). (a) Quantitative reverse transcriptase–PCR measurement of relative gene expression of Cox-1, Cox-2, and IL-1β from AMs and interstitial lung from uninfected control and BMT mice normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH; n=3 control; n=3 BMT per group). (b) PGE2 measurements by enzyme-linked immunosorbent assay (ELISA) from overnight culture of untreated AMs from control and BMT mice (n=4 control; n=4 BMT). (c) IL-1β measurements by ELISA from AMs infected or not in vitro with PA01 (multiplicity of infection: 10), treated or not with 100 nM of PGE2 (n=3). (d) IL-1β, (e) tumor necrosis factor (TNF)-α, and (f) IL-6 measurements by ELISA from bronchoalveolar lavage fluid of lipopolysacccharide (LPS; 50 μg)-treated control and BMT mice (n=10 control, n=10 BMT). Statistics are Student’s t-test between comparative groups. *P<0.05; **P<0.01; ****P<0.0001; ND, not detected; NS, non-significant. Data are representative of at least two independent experiments.
PGE2 induces higher levels of IL-1β upon pathogenic stimuli
To assess whether PGE2 could induce IL-1β in macrophages other than AMs, we prepared bone marrow–derived macrophages (BMDMs) from healthy mice and pretreated them with PGE2 prior to PA01 infection, in vitro. Similar to AMs, we detected higher IL-1β, but not IL-10, in supernatants of PGE2- stimulated cells compared with non-treated cells post-PA01 (Figure 4a,b). In addition, to detect whether PGE2-mediated higher levels of IL-1β with different stimuli, we pretreated BMDMs with and without PGE2 and stimulated cells with LPS and/or heat-killed PA01. In all cases, PGE2 pretreatment increased the levels of IL-1β compared with non-treated cells (Figure 4c). A canonical method to achieve IL-1β secretion is to activate the NOD-like receptor family, pyrin domain containing 3; nucleotide-binding leucine-rich repeat-containing pyrin receptor 3 inflammasome by stimulation of the pannexin-1 channel and the purinergic P2X7 by LPS priming and ATP treatment.25, 28, 29, 30 We also detected higher IL-1β in BMDMs pretreated with PGE2 and treated with LPS and ATP when compared with treatment with LPS and ATP alone (Figure 4d). Furthermore, we noted higher levels of IL-1β in BMDMs pretreated with PGE2 and infected with methicillin-resistant Staphylococcus aureus (MRSA) or Streptococcus pneumoniae compared with infected BMDMs not treated with PGE2 (see Supplementary Figure S2B). MRSA and S. pneumoniae are also main pathogens that infect immunosuppressed individuals.6, 19, 31, 32

Prostaglandin E2 (PGE2) increases interleukin (IL)-1β in bone marrow–derived macrophages (BMDMs) upon pathogenic stimulation. (a, b) IL-1β and IL-10 measurements from supernatants of PA01-infected (multiplicity of infection (MOI): 10) BMDMs treated or not with 100 nM PGE2. (c) IL-1β measurements from supernatant of BMDMs treated with lipopolysaccharide (LPS) derived from P. aeruginosa (500 ng ml−1), heat-killed PA01 (MOI:10), and (d) adenosine triphosphate (ATP; 1 mM) with LPS with or without 100 nM of PGE2. Supernatant was taken 2 h poststimulation. One-way analysis of variance with Bonferroni’s posttest. *P<0.05, **P<0.01, ****P<0.0001; NS, non-significant. Data are representative of three independent experiments.
PGE2-mediated increase in IL-1β is dependent on activation of transcription factor CREB via increased levels of cAMP dependent on EP2 and EP4 signaling
We next probed PGE2 signaling pathways. We stimulated PGE2 receptors using pharmacological agonists for the EP2, EP3, and EP4 receptors.17 A selective EP1 agonist is not available. We found that prestimulating BMDMs with EP2 and EP4 agonists lead to higher levels of IL-1β post-P. aeruginosa infection (Figure 5a). EP3 stimulation did not lead to higher IL-1β (data not shown). EP2 and EP4 receptors activate AC and increase cAMP from cytosolic ATP. We pretreated BMDMs with the AC stimulator forskolin prior to PA01 infection and observed higher levels of IL-1β in forskolin-pretreated BMDMs (Figure 5b). Moreover, as increased levels of cAMP lead to the activation of PKA and Epac (exchange protein directly activated by cAMP), we used agonists for the activation of these two proteins.33 We found higher levels of IL-1β in supernatant of BMDMs prestimulated with PKA and Epac agonists prior to PA01 infection (Figure 5c). PKA and Epac activation lead to activation of the transcription factor CREB, thus we used a CREB inhibitor during PGE2 stimulation prior to PA01 infection. We decreased the levels of PGE2-mediated IL-1β using the CREB inhibitor (Figure 5d). Thus PGE2 increases IL-1β by activation of the transcription factor CREB via stimulation of EP2 and/or EP4 receptors.

Prostaglandin E2 (PGE2) mediated increase in interleukin (IL)-1β is dependent on activation of transcription factor CREB (cyclic adenosine monophosphate (cAMP) response element binding) by increasing the levels of cAMP dependent on EP2 and EP4 signaling. IL-1β measurements from supernatant of bone marrow–derived macrophages (BMDMs) treated or not with PAO1 (multiplicity of infection (MOI): 10) with or without (a) EP2 agonist (1 μM, Butaprost), EP4 agonist (500 nM, ONO-AE1-329), (b) forskolin (25 μM), PGE2 (100 nM), (c) protein kinase A (PKA) agonist (50 μM, 6-BNZ-cAMP), and Epac agonist (50 μM, 8-pcpt-2′-OM-cAMP). (d) IL-1β protein measurement from supernatants of BMDMs treated or not with 100 nM of PGE2, CREB inhibitor (100 μM, Naphthol AS-E phosphate) and infected or not with PA01 (MOI:10). In all cases, supernatant was taken 2 h after infection. One-way analysis of variance with Bonferroni’s posttest. *P<0.05; ***P<0.001; ****P<0.0001; NS, non-significant. Data are representative of two independent experiments.
PGE2-dependent IL-1β release post- P. aeruginosa infection can be mediated by canonical or non-cannonical inflammasomes and is independent of autophagy inhibition
Mycobacterium tuberculosis and Candida albicans have been shown to induce IL-1β by a non-canonical inflammasome pathway dependent on caspase-8 in macrophages.34, 35 Although a role of caspase-8-mediated IL-1β processing post-P. aeruginosa infection has not been proposed, research has shown that P. aeruginosa–infected caspase-1-deficient mice can secrete normal levels of IL-1β compared with control mice.36 Thus other proteases might have a role in processing pro-IL-1β post-P. aeruginosa infection. We stimulated macrophages with or without caspase-1 and caspase-8 inhibitors. Interestingly, in both cases, IL-1β secretion was inhibited with or without the effects of PGE2 (Figure 6a,b). Stimulating macrophages with LPS and ATP in the presence or absence of caspase-1 and/or caspase-8 inhibitors confirmed the specificity of these inhibitors (see Supplementary Figure S3). AMs can clear P. aeruginosa infection by autophagy and IL-1β release can be negatively regulated by autophagy.24, 25 Autophagy can be induced by serum starvation; thus we tested the effects of PGE2 on autophagy-enhanced clearance of P. aeruginosa carried out under conditions of serum deprivation. Our results indicate PGE2 can inhibit autophagy-dependent clearance in AMs (Figure 7a). To detect whether there is a direct effect of PGE2 on autophagy, we treated BMDMs with different concentrations of PGE2 and detected the levels of autophagy-related proteins, LC3 and P62. We detected downregulation of LC3 and accumulation of P62 when we treated AMs with PGE2. These changes are characteristic of autophagy inhibition (Figure 7b and Supplementary Figure S4). In addition, we induced autophagy by serum starvation with or without PGE2 stimulation and determined that PGE2 inhibits the levels of autophagy-related protein, ATG5, by western blotting and quantitative reverse transcriptase–PCR (Figure 7c). Although we found PGE2 inhibits autophagy, autophagy-deficient BMDMs were still able to upregulate IL-1β release when PGE2 was present, suggesting that PGE2 inhibition of autophagy is not required for processing and upregulation of IL-1β (Figure 7d).

PAO1 can use cannonical or non-cannonical inflammasomes to secrete interleukin (IL)-1β. IL-1β measurements from supernatant of bone marrow–derived macrophages treated or not with (a) Caspase 8 inhibitor (10 nM) or Caspase 1 inhibitor (10 nM) with or without prostaglandin E2 (PGE2; 100 nM). Supernatant was taken 2 h after infection. One-way analysis of variance with Bonferroni’s posttest. **P<0.01; ****P<0.0001; NS, non-significant. Data are representative of two independent experiments.

Inhibition of autophagy impacts bacterial killing but not interleukin (IL)-1β release. (a) Alveolar macrophages (AMs) were cultured in complete media or were nutrient starved in Earle’s Balanced Salt Solution (EBSS) (#1) or Hanks’ Balanced Salt solution (HBSS) (#2) in the presence or absence of prostaglandin E2 (PGE2; 100 nM) for 2 h. (b) AMs were collected from control mice and stimulated for 1 h in the presence of serum-free media alone with or without a dose response of PGE2 added (50–500 nM) for 1 h. Cell lysates were then analyzed for the levels of LC3 and p62 by western blotting compared with glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Blot shown is representative of three experiments. (c) AMs were subjected to culture in complete media or were serum starved for 1 h in the presence or absence of 100 nM PGE2 before RNA was prepared and analyzed for the expression of the autophagy gene ATG5 relative to β-actin by quantitative reverse transcriptase–PCR (n=4); (bottom) cell lysates were also taken from samples assessed for the levels of ATG5 and β-actin by western blotting. (d) Bone marrow–derived macrophages (BMDMs) from autophagy-deficient LC3−/− mice were stimulated with 100 nM PGE2, PAO1 (multiplicity of infection: 10) or the combination for 2 h before supernatants were collected and measured for IL-1β by enzyme-linked immunosorbent assay (n=4); Data are representative of two independent experiments and statistics were measured by one-way analysis of variance with Bonferroni post hoc test. *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001; ND, not detected; NS, non-significant.
PGE2 elevates the levels of IL-1β transcripts by EP2 and EP4 stimulation in human and mouse cells
Considering that neither autophagy inhibition nor inflammasome activation explain why PGE2 boosted IL-1β release, we next studied the transcriptional effects of PGE2 on IL-1β. We treated BMDMs with PGE2 in a dose- and time-dependent manner and detected significant increases in IL-1β transcripts as soon as 2 h post-PGE2 stimulation (Figure 8a,b). Furthermore, we confirmed that elevated levels of IL-1β transcripts by PGE2 stimulation are dependent on increasing levels of cAMP by stimulation of EP2 and/or EP4 receptors but not by EP3 stimulation (Figure 8c). Moreover, we confirmed our findings in human peripheral monocytes and human AMs (Figure 9).

Prostaglandin E2 (PGE2) elevates the levels of interleukin (IL)-1β transcripts by EP2 and EP4 stimulation. (a) Bone marrow–derived macrophages (BMDMs) were treated or not with 100 nM PGE2 for 1, 2, 4, and 8 h before RNA was prepared and analyzed for the expression of IL-1β gene. (b) BMDMs were treated or not with 10, 100, and 500 nM of PGE2 during 4 h before RNA was prepared and analyzed for the expression of IL-1β. (c) BMDMs were treated with EP2 agonist (1 μM, Butaprost), EP3 agonist (10 nM, Sulprostone), EP4 agonist (500 nM, ONO-AE1-329), and forskolin (25 μM) before RNA was prepared and analyzed. All RNA data were normalized to the expression levels of glyceraldehyde 3-phosphate dehydrogenase. Data are representative of two independent experiments and statistics were measured by one-way analysis of variance with Bonferroni post hoc test. *P<0.05; ****P<0.0001; NS, non-significant.

Human alveolar macrophages (AMs) and peripheral monocytes upregulate interleukin (IL)-1β under prostaglandin E2 (PGE2) stimulation. IL-1β relative expression from 100 nM PGE2 stimulated (a) human peripheral monocytes and (b) human AMs normalized to glyceraldehyde 3-phosphate dehydrogenase. (c) IL-1β protein measurements from supernatant of PGE2-treated or not peripheral macrophages during or not PA01 infection. Statistics are Student’s t-test between comparative groups. *P<0.05; **P<0.01; ****P<0.0001; ND, not detected.
Decreasing levels of PGE2 reduces P. aeruginosa-mediated lung tissue injury
Increased levels of IL-1β post-P. aeruginosa infection aggravate lung tissue injury.4, 5 Therapeutic strategies using caspase-1 inhibitors reduced severity of IL-1β pulmonary injury.26 However, other research has shown that inhibiting IL-1β signaling has no impact on bacterial burden or immune cell recruitment, with minimal effects on lung injury.36 As we have shown that PGE2 elevation in BMT mice aggravates lung injury, we tested the effect that PGE2 inhibition had on lung injury outcomes. We instilled P. aeruginosa to control and BMT mice with or without administration of the COX inhibitor, indomethacin. We noted reduced IL-1β in the BALF as well as lower levels of protein leakage, suggesting that inhibition of PGE2 can decrease levels of ALI post-P. aeruginosa infection (Figure 10).

Decreasing levels of prostaglandin E2 (PGE2) by indomethacin treatment leads to decreased interleukin (IL)-1β in the lung after P. aeruginosa infection in bone marrow transplant (BMT) mice. (a) IL-1β and (b) albumin measurements from the bronchoalveolar lavage fluid (BALF) of control and BMT mice treated or not with indomethacin (1.2 mg kg−1) for 24 h after PA01 infection; measurements were carried out by enzyme-linked immunosorbent assay. (n=14 control; n=14 BMT; n=14 BMT treated with indomethacin); (c) PA01 colony-forming unit (CFU) measurement in BAL from infected mice after 24 h. (n=10 control; n=10 BMT; n=10 BMT treated with indomethacin). Data are representative of at least two independent experiments. Statistics were measured by one-way analysis of variance with Bonferroni post hoc test. *P<0.05; **P<0.01; ***P<0.001; NS, non-significant.
Discussion
In February 2017, the World Health Organization published a report containing a list of 12 bacterial pathogens for which new therapeutic strategies are urgently needed (http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/). P. aeruginosa was considered to be of critical importance. P. aeruginosa is an opportunistic pathogen that causes minimal pathogenicity in healthy individuals but major mortality and morbidity in HSCT patients.2, 6, 19, 31, 37, 38 Susceptibility to this pathogen has been reported to occur even after immune reconstitution following HSCT.2, 6, 31 HSCT patients have high levels of PGE2 in the blood and BALF when compared with healthy individuals.18, 19, 20 We have previously reported that elevated PGE2 is causally related to the impaired ability of AMs from BMT mice to phagocytize and kill bacteria.20, 23, 39 The purpose of the current study was to determine whether PGE2 production post-BMT was responsible for enhanced lung injury postinfection as well, and if so, by what mechanism. We now know that inflammasome-dependent IL-1β secretion is induced by P. aeruginosa, and better health outcomes have been reported by inhibiting IL-1β signaling.4, 5, 12, 26, 36 In addition, recent articles have linked PGE2 signaling with IL-1β regulation.27, 28, 40 However, reports have shown contradictory effects of PGE2 on IL-1β release.27, 28 Thus, in this study, we focused on the PGE2-mediated effects on IL-1β response to P. aeruginosa infection by AMs from BMT mice in our quest to determine new therapeutic strategies.
A dose of 2 × 106 colony-forming unit (CFU) P. aeruginosa instilled into untransplanted mice causes a moderate infection.41 Yet, BMT mice have difficulty clearing this bacterial dose and 100% of them die within 48 h.23 Interestingly, the susceptibility to P. aeruginosa in BMT mice is seen even after immune reconstitution.6, 23, 42 Here we show evidence that BMT mice have severe vascular and epithelial leakage, indicating a more severe ALI post-P. aeruginosa infection. ALI can be caused by exacerbated levels of pro-inflammatory cytokines. Thus we searched for upregulated levels of different pro-inflammatory cytokines (IL-12, IL-6, TNF-α, and IL-1β) and found that only IL-1β was significantly higher in BMT mice when compared with untransplanted mice postinfection. In addition, we noticed a direct correlation between IL-1β, PGE2, and albumin levels in the BALF but not a correlation with other pro-inflammatory cytokines, such as TNF-α.
Because we observed exacerbated levels of IL-1β post-P. aeruginosa infection in BMT mice, we analyzed the capability of AMs from BMT mice to secrete IL-1β. Interestingly, AMs from BMT mice have higher levels of IL-1β transcripts compared with control mice, correlating with higher levels of COX1 and COX2 transcripts. Furthermore, stimulating AMs with PGE2 prior to infection with P. aeruginosa also leads to higher levels of IL-1β compared with untreated AMs. These findings show that higher levels of PGE2 in AMs from BMT mice might influence the higher levels of IL-1β post-P. aeruginosa.
As higher amounts of bacterial burden in BMT mice might influence the levels of IL-1β, we adopted a bacterial-free model of ALI dependent on LPS instillation.43 We were able to get exacerbated levels of IL-1β, but not IL-6 and TNF-α, in BMT mice compared with untransplanted mice. In addition, in an in vitro model where we controlled the stimulants, we detected that PGE2 could increase secretion of IL-1β in BMDMs by a wide array of pathogenic stimuli, such as LPS, live and heat killed P. aeruginosa, MRSA infection, and S. pneumoniae infection. All stimuli induce increased IL-1β after previous exposure to PGE2 in macrophages. Thus PGE2 signaling acts as signal one for IL-1β secretion. Interestingly, dual stimulation of macrophages with PGE2 and another signal one stimulant, LPS, can lead to increased IL-1β secretion. This phenomenon is likely explained by the moderate activation of caspase-1 (signal two) in macrophages by LPS stimulation as previously noted.44
Recent reports have shown an opposite effect of PGE2 regulation on IL-1β27 than our results. We compared their PGE2-stimulation method, based on 5 min of stimulation by PGE2, side by side with our stimulation method, based on 4 h prestimulation with PGE2. As expected, we were able to obtain exacerbated levels of IL-1β after 4 h of PGE2 stimulation, whereas 5 min of stimulation had no effect on IL-1β regulation (see Supplementary Figure S5). Thus these comparative results suggest that the kinetics of PGE2 exposure is important. However, in the setting of HCST, it is important to remember that PGE2 levels are chronically elevated,18, 19 and thus in this setting, PGE2 is promoting IL-1β.
PGE2 signaling is mediated by four members of the G-coupled protein receptor family termed EP1–EP4.17 We determined that stimulating the EP2 and EP4 receptors in macrophages prior to P. aeruginosa infection leads to higher levels of IL-1β when compared with non-stimulated macrophages. Furthermore, as EP2/EP4 receptors share the ability to activate PKA and Epac, we activated PKA or Epac with the use of intracellular agonists prior to P. aeruginosa infection and detected higher levels of IL-1β when compared with non-stimulated macrophages. We were able to abolish IL-1β secretion in P. aeruginosa–infected mice by stimulating macrophages with an inhibitor for the CREB transcription factor. These data indicate that the PGE2-mediated increase in IL-1β is due to CREB transcription factor activation by EP2/EP4-mediated stimulation of PKA and/or Epac. Although we present evidence that CREB is mediating PGE2-dependent elevation of IL-1β, we do not discard the possibility that other transcription factors such as nuclear factor-κB28 may also have a role. Furthermore, while both EP2 and EP4 can mediate the transcriptional effect of PGE2 in macrophages from control mice, in the setting of BMT, we have previously shown that levels of EP2 are upregulated, while EP4 is slightly downregulated on AMs and an EP2 antagonist, AH6809, was able to mimic effects of indomethacin on AM phagocytosis.39
Researchers have established autophagy as a main mechanism of P. aeruginosa clearance24 as well as IL-1β regulation.10, 25 We tested the effects of PGE2 in the regulation of autophagy and determined that PGE2 can inhibit autophagy-induced clearance of P. aeruginosa. Therefore, these data suggested that PGE2-mediated IL-1β increase might be due to autophagy inhibition. However, we detected high levels of IL-1β protein in autophagy-deficient macrophages after PGE2 administration and infection. Thus the mechanism of PGE2-mediated increase in IL-1β production cannot be attributed to impaired clearance of inflammasome components as a result of defective autophagy.25 Although we previously demonstrated that PGE2 was associated with impaired autophagy in neutrophils,19 this is the first description of its ability to limit autophagy in macrophages. Thus PGE2 may be a common negative regulator of autophagic flux in other cell types as well. Future work will be needed to understand what impact this negative regulation may have on processes, such as epithelial repair.
In macrophages, IL-1β secretion can be mediated by the cysteine proteases, caspase-1 and caspase-8.35 Thus we tested the role of these two caspases during P. aeruginosa infection. We determined that we could abolish the secretion of IL-1β prior to P. aeruginosa infection with the use of inhibitors to caspase-1 or caspase-8. To our knowledge, these are the first data to link IL-1β secretion post-P. aeruginosa to caspase-8; this pathway has been previously established for M. tuberculosis and fungal infection.34 These results suggested that P. aeruginosa infection can induce the activation of IL-1β release using either caspase-1 and/or caspase-8. If true, this suggests that P. aeruginosa may be able to stimulate non-cannonical inflammasome activation, possibly via cross-reactivity caused by recognition of P. aeruginosa by dectin receptors,45 which are known to be linked to caspase 8 activation.34
Our data suggest that PGE2 can influence a pro-inflammatory environment by exacerbating the levels of IL-1β in macrophages. We detected that PGE2 can mediate a massive increase in IL-1β transcripts. We detected increased IL-1β transcripts in macrophages within an hour poststimulation with PGE2. Increases in IL-1β transcripts were dependent on stimulation of EP2 and EP4 receptors but not by EP3. Although we did not have access to an EP1-selective agonist, we would not anticipate this receptor to regulate cAMP levels.17 Additionally, we were able to detect an increase in IL-1β transcripts following activation of cAMP with forskolin. Moreover, PGE2 stimulation increased transcription of IL-1β in human AMs and human monocyte-derived macrophages. Altogether, we conclude that PGE2 can strongly prime macrophages for IL-1β but not other cytokines and, upon pathogenic stimulation, will lead to exacerbated levels of IL-1β causing IL-1β-mediated injury. Thus we expected to decrease the IL-1β-mediated lung injury post-BMT with the use of COX inhibitors. When we tested the effects of PGE2 inhibition in BMT mice, we were not only able to decrease bacterial load as previously reported7 but also reduce protein leakage and IL-1β in the lung. When our results are taken together, we identified new mechanisms by which P. aeruginosa causes life-threatening effects in HSCT patients. These findings can help in the development of new therapeutic strategies that can improve outcome of HSCT patients with pulmonary complications due to P. aeruginosa and possibly other pathogens. We speculate that COX inhibitors or possibly Anakinra may offer therapeutic benefit at limiting lung injury caused by bacterial infection post-HSCT.
Methods
Mice
Male C57BL/6J mice were purchased from Jackson Laboratories (Bar Harbor, ME) and used at 6–8 weeks of age. Experiments were approved by the University of Michigan Institutional Animal Care and Use Committee.
Reagents
The following are the reagents used: PGE2 (1–1000 nM; Cayman Chemicals, Ann Arbor, MI); forskolin (25 μM; Cayman Chemicals); P. aeruginosa LPS (100 ng ml−1; Sigma-Aldrich, St. Louis, MO); ATP (1 μM; Sigma-Aldrich); caspase-1 inhibitor (10 nM Ac-YVAD-CHO; Enzo Life Sciences, Farmingdale, NY); caspase-8 inhibitor (10 nM Ac-IETD-CHO; BD Biosciences, San Jose, CA); CREB inhibitor (100 μM Naphthol AS-E phosphate; Sigma-Aldrich); EP2 agonist (1 μM Butaprost; Cayman Chemicals), EP3 agonist (10 nM Sulprostone; Cayman Chemicals) EP4 agonist (500 nM ONO-AE1-329; Sigma-Aldrich); PKA agonist (50 μM 6-BNZ-cAMP; Biolog, Hayward, CA); and Epac agonist (50 μM 8-pcpt-2′-OM-cAMP; Biolog).
Cells
AMs were harvested by performing BAL on C57BL/6J mice using 20 ml of supplemented Dulbecco’s Modified Eagle Medium (89% Dulbecco’s Modified Eagle Medium, 10% fetal bovine serum, 1% Pen–Strep, and 5 mM EDTA) as described in Domingo-Gonzalez et al.22 To prepare RNA from interstitial cells, lungs were homogenized in TRIzol (Thermo Fisher Scientific; Waltham, MA) after BAL using a tissue homogenizer (OMNI International; cat# Th115). BMDMs were obtained by flushing bone marrow cells from the femur and tibia of C57BL/6J mice as explained in Trouplin et al.46 Briefly, bone marrow cells were incubated at 37 °C in 5% CO2 for 7 days in BMDM differentiation media (59% Iscove's Modified Dulbecco’s Medium, 30% L-929 cell supernatant, 10% fetal calf serum, and 1% Pen–Strep). Human AMs were collected by ex vivo BAL of human lungs that were not used for transplant. Human monocyte-derived macrophages were harvested by gradient centrifugation with the use of Ficoll-Paque Plus (GE Healthcare, Uppsala, Sweden) from peripheral blood of healthy donors followed by differentiation with human recombinant colony stimulation factor-1 (50 ng ml−1; R&D Systems, Minneapolis, MN) for 7 days.
Bacteria
P. aeruginosa (PA01) and S. aureus (US300) were grown in tryptic soy broth and nutrient broth, respectively, and incubated with gentle agitation overnight at 37 °C. S. pneumoniae (serotype 3, 6303) was grown in Todd Hewitt Broth with 0.5% yeast extract and incubated overnight at 37 °C and 5% CO2. CFU were determined by absorbance relative to known standard curves.
Bone marrow transplantation
Healthy 6–8-week-old C57BL/6J male mice were lethally irradiated with a split dose of 13 Gray (13 Gy) with the use of an X-rad 320 irradiator (Kimtron Inc., Oxford, CT). Irradiated mice were infused with 5 × 106 bone marrow cells from a genetically identical donor (C57BL/6J). Mice were housed for 5 weeks after infusion to achieve full reconstitution of their immune system. The percentage of donor-derived cells was ∼95±1% in the spleen and 82±2% in the lung at this time point, as assessed by transplanting CD45.1+ bone marrow into C57BL/6J CD45.2+ mice as shown in Hubbard et al.20
Model of infection
P. aeruginosa infection was carried out by intratracheal inoculation of 5 × 105 CFU of PA01 in 50 μl of saline solution. The control group received saline solution as placebo. Harvest was performed 24 h after infection; mice were first killed with CO2 asphyxiation and BAL was performed with 1 ml of phosphate-buffered saline containing 5 mM EDTA, followed by lung perfusion and harvest. CFU measurements were obtained by serial dilution and plating on nutrient agar plates. LPS was given by intratracheal inoculation of 50 μg P. aeruginosa–derived LPS in 40 μl of saline solution.
Tetrazolium dye reduction assay of bacterial killing
AMs from C57BL/6 mice or BMDMs were placed into duplicate 96-well plates: one experimental plate and one control plate. Cells from both plates were infected with IgG-opsonized P. aeruginosa (multiplicity of infection 50:1) for 30 min at 37 °C. The cells on the experimental plate were washed and then incubated with or without treatments at 37 °C for 120 min, whereas the cells on the control plate were washed and then lysed with 0.5% saponin in tryptic soy broth (Sigma-Aldrich) and placed at 4 °C. After 2 h, the cells from the experimental plate were lysed with 0.5% saponin in tryptic soy broth. Both plates were then incubated in a shaker at 37 °C for 4 h. Thiazolyl blue Tetrazolium Bromide assay was performed as recommended by the company. Briefly, a total of 5 mg ml−1 Thiazolyl blue Tetrazolium Bromide Sigma-Aldrich was added to each plate and incubated for 30 min. Solubilization solution was added to dissolve formazan salts, and the absorbance was read at 595 nm (A595). Results were expressed as the percentage of survival of ingested bacteria normalized to the percentage of control, where the A595 experiment values were divided by the average of the A595 control values. Survival of ingested bacteria=(A595 experimental plate/A595 control plate) × 100%.
Enzyme-linked immunosorbent assay, protein measurement, immunofluorescence, and immunoblotting
Cytokine measurement was performed with the use of R&D duoset enzyme-linked immunosorbent assay kits for IL-1β, IL-10, and IL-6. Albumin measurements were performed with the Bethyl Laboratory (Montgomery, TX) mouse albumin enzyme-linked immunosorbent assay kit. Protein quantification was achieved with the use of Pierce BCA Assay Kit from Thermo Fisher. Immunofluorescence images were taken with an Olympus 500 confocal microscope (Olympus Corporation, Center Valley, PA).
Histology
Hematoxylin and Eosin stain was performed after perfusion of the lung with phosphate-buffered saline and tissue fixation with 10% formalin. Tissues were left in 10% formalin overnight before replacing fluid with 70% ethanol. Lung samples were processed and stained by McClinchey Histology Services (Stockbridge, MI).
Quantitative real-time PCR
mRNA was isolated using TRIzol according to the manufacturer’s instructions. Relative gene expression measurements were achieved with the use of a Step-one plus real-time PCR system from Applied Biosystems (Foster City, CA). Gene-specific primers and probes were designed with the GenScript Real-time PCR primer design software (Genscript Biotech Corporation, Piscataway, NJ).
Statistical analysis
Graphpad Prism version 6 software (Graphpad Prism Software Inc., La Jolla, CA) was used to analyze experimental results. When groups of two were compared, Student’s t-test was used to determine statistical significance. Groups of ⩾3 were compared using one-way analysis of variance with Bonferroni multiple mean comparisons. Fisher’s exact test was used to compare clearance of bacteria and albumin levels between infected mice.
References
Gratwohl, A. et al. Hematopoietic stem cell transplantation: a global perspective. JAMA 303, 1617–1624 (2010).
Ullah, K. et al. Post-transplant infections: single center experience from the developing world. Int. J. Infect. Dis. 12, 203–214 (2008).
Danby, R. & Rocha, V. Improving engraftment and immune reconstitution in umbilical cord blood transplantation. Front. Immunol. 5, 68 (2014).
Wonnenberg, B. et al. The role of IL-1beta in Pseudomonas aeruginosa in lung infection. Cell Tissue Res. 364, 225–229 (2016).
Cohen, T.S. & Prince, A.S. Activation of inflammasome signaling mediates pathology of acute P. aeruginosa pneumonia. J. Clin. Invest. 123, 1630–1637 (2013).
Domingo-Gonzalez, R., Katz, S., Serezani, C.H., Moore, T.A., Levine, A.M. & Moore, B.B. Prostaglandin E2-induced changes in alveolar macrophage scavenger receptor profiles differentially alter phagocytosis of Pseudomonas aeruginosa and Staphylococcus aureus post-bone marrow transplant. J. Immunol. 190, 5809–5817 (2013).
Sadikot, R.T. et al. Bacterial clearance of Pseudomonas aeruginosa is enhanced by the inhibition of COX-2. Eur. J. Immunol. 37, 1001–1009 (2007).
Aloush, V., Navon-Venezia, S., Seigman-Igra, Y., Cabili, S. & Carmeli, Y. Multidrug-resistant Pseudomonas aeruginosa: risk factors and clinical impact. Antimicrob. Agents Chemother. 50, 43–48 (2006).
Lavoie, E.G., Wangdi, T. & Kazmierczak, B.I. Innate immune responses to Pseudomonas aeruginosa infection. Microbes Infect. 13, 1133–1145 (2011).
Jabir, M.S. et al. Caspase-1 cleavage of the TLR adaptor TRIF inhibits autophagy and beta-interferon production during Pseudomonas aeruginosa infection. Cell Host Microbe 15, 214–227 (2014).
Nakamura, S. et al. Toll-like receptor 4 agonistic antibody promotes host defense against chronic Pseudomonas aeruginosa lung infection in mice. Infect. Immun. 84, 1986–1993 (2016).
Tolle, L. et al. Redundant and cooperative interactions between TLR5 and NLRC4 in protective lung mucosal immunity against Pseudomonas aeruginosa. J. Innate Immun. 7, 177–186 (2015).
Kim, Y.K., Shin, J.S. & Nahm, M.H. NOD-like receptors in infection, immunity, and diseases. Yonsei Med. J. 57, 5–14 (2016).
Imagawa, T. et al. Safety and efficacy of canakinumab in Japanese patients with phenotypes of cryopyrin-associated periodic syndrome as established in the first open-label, phase-3 pivotal study (24-week results). Clin. Exp. Rheumatol. 31, 302–309 (2013).
Dostert, C., Petrilli, V., Van Bruggen, R., Steele, C., Mossman, B.T. & Tschopp, J. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 320, 674–677 (2008).
Dennis, E.A. & Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 15, 511–523 (2015).
O'Callaghan, G. & Houston, A. Prostaglandin E2 and the EP receptors in malignancy: possible therapeutic targets? Br. J. Pharmacol. 172, 5239–5250 (2015).
Cayeux, S.J., Beverley, P.C., Schulz, R. & Dorken, B. Elevated plasma prostaglandin E2 levels found in 14 patients undergoing autologous bone marrow or stem cell transplantation. Bone Marrow Transplant. 12, 603–608 (1993).
Domingo-Gonzalez, R. et al. Inhibition of neutrophil extracellular trap formation after stem cell transplant by prostaglandin E2. Am. J. Respir. Crit. Care Med. 193, 186–197 (2016).
Hubbard, L.L., Ballinger, M.N., Wilke, C.A. & Moore, B.B. Comparison of conditioning regimens for alveolar macrophage reconstitution and innate immune function post bone marrow transplant. Exp. Lung Res. 34, 263–275 (2008).
Hubbard, L.L. et al. A role for IL-1 receptor-associated kinase-M in prostaglandin E2-induced immunosuppression post-bone marrow transplantation. J. Immunol. 184, 6299–6308 (2010).
Domingo-Gonzalez, R. et al. Transforming growth factor-beta induces microRNA-29b to promote murine alveolar macrophage dysfunction after bone marrow transplantation. Am J Physiol Lung Cell Mol Physiol 308, L86–L95 (2015).
Ojielo, C.I. et al. Defective phagocytosis and clearance of Pseudomonas aeruginosa in the lung following bone marrow transplantation. J. Immunol. 171, 4416–4424 (2003).
Yuan, K. et al. Autophagy plays an essential role in the clearance of Pseudomonas aeruginosa by alveolar macrophages. J. Cell Sci. 125 (Pt 2), 507–515 (2012).
Shi, C.S. et al. Activation of autophagy by inflammatory signals limits IL-1beta production by targeting ubiquitinated inflammasomes for destruction. Nat. Immunol. 13, 255–263 (2012).
Wu, D.D. et al. Inhibition of alveolar macrophage pyroptosis reduces lipopolysaccharide-induced acute lung injury in mice. Chin. Med. J. (Engl.) 128, 2638–2645 (2015).
Mortimer, L., Moreau, F., MacDonald, J.A. & Chadee, K. NLRP3 inflammasome inhibition is disrupted in a group of auto-inflammatory disease CAPS mutations. Nat. Immunol. 17, 1176–1186 (2016).
Zoccal, K.F. et al. Opposing roles of LTB4 and PGE2 in regulating the inflammasome-dependent scorpion venom-induced mortality. Nat. Commun. 7, 10760 (2016).
Munoz-Planillo, R., Kuffa, P., Martinez-Colon, G., Smith, B.L., Rajendiran, T.M. & Nunez, G. K(+) efflux is the common trigger of NLRP3 inflammasome activation by bacterial toxins and particulate matter. Immunity 38, 1142–1153 (2013).
Yang, D., He, Y., Munoz-Planillo, R., Liu, Q. & Nunez, G. Caspase-11 requires the pannexin-1 channel and the purinergic P2X7 pore to mediate pyroptosis and endotoxic shock. Immunity 43, 923–932 (2015).
Domingo-Gonzalez, R. & Moore, B.B. Innate immunity post-hematopoietic stem cell transplantation: focus on epigenetics. Adv. Neuroimmune Biol. 5, 189–197 (2014).
Ballinger, M.N., McMillan, T.R. & Moore, B.B. Eicosanoid regulation of pulmonary innate immunity post-hematopoietic stem cell transplantation. Arch. Immunol. Ther. Exp. (Warsz.) 55, 1–12 (2007).
Cheng, X., Ji, Z., Tsalkova, T. & Mei, F. Epac and PKA: a tale of two intracellular cAMP receptors. Acta Biochim. Biophys. Sin. (Shanghai) 40, 651–662 (2008).
Gringhuis, S.I. et al. Dectin-1 is an extracellular pathogen sensor for the induction and processing of IL-1beta via a noncanonical caspase-8 inflammasome. Nat. Immunol. 13, 246–254 (2012).
Afonina, I.S., Muller, C., Martin, S.J. & Beyaert, R. Proteolytic processing of interleukin-1 family cytokines: variations on a common theme. Immunity 42, 991–1004 (2015).
Al Moussawi, K. & Kazmierczak, B.I. Distinct contributions of interleukin-1alpha (IL-1alpha) and IL-1beta to innate immune recognition of Pseudomonas aeruginosa in the lung. Infect. Immun. 82, 4204–4211 (2014).
Lossos, I.S. et al. Bacterial pneumonia in recipients of bone marrow transplantation. A five-year prospective study. Transplantation 60, 672–678 (1995).
Whittle, A.T., Davis, M., Shovlin, C.L., Ganly, P.S., Haslett, C. & Greening, A.P. Alveolar macrophage activity and the pulmonary complications of haematopoietic stem cell transplantation. Thorax 56, 941–946 (2001).
Ballinger, M.N. et al. Critical role of prostaglandin E2 overproduction in impaired pulmonary host response following bone marrow transplantation. J. Immunol. 177, 5499–5508 (2006).
Sokolowska, M. et al. Prostaglandin E2 inhibits NLRP3 inflammasome activation through EP4 receptor and intracellular cyclic AMP in human macrophages. J. Immunol. 194, 5472–5487 (2015).
Zhang, Y., Li, X., Carpinteiro, A., Goettel, J.A., Soddemann, M. & Gulbins, E. Kinase suppressor of Ras-1 protects against pulmonary Pseudomonas aeruginosa infections. Nat. Med. 17, 341–346 (2011).
Hubbard, L.L., Wilke, C.A., White, E.S. & Moore, B.B. PTEN limits alveolar macrophage function against Pseudomonas aeruginosa after bone marrow transplantation. Am. J. Respir. Cell Mol. Biol. 45, 1050–1058 (2011).
Bosmann, M. et al. CD11c+ alveolar macrophages are a source of IL-23 during lipopolysaccharide-induced acute lung injury. Shock 39, 447–452 (2013).
Schumann, R.R. et al. Lipopolysaccharide activates caspase-1 (interleukin-1-converting enzyme) in cultured monocytic and endothelial cells. Blood 91, 577–584 (1998).
Mennink-Kersten, M.A., Ruegebrink, D. & Verweij, P.E. Pseudomonas aeruginosa as a cause of 1,3-beta-D-glucan assay reactivity. Clin. Infect. Dis. 46, 1930–1931 (2008).
Trouplin, V., Boucherit, N., Gorvel, L., Conti, F., Mottola, G. & Ghigo, E. Bone marrow-derived macrophage production. J. Vis. Exp. e50966 (2013).
Acknowledgements
This work was supported by NIH grants: AI117229, HL127805, and HL119682 (to B.B.M.). G.J.M.-C. was supported by T32AI007413 and a University of Michigan Rackham Merit fellowship. We thank Dr Nick Lukacs (University of Michigan, Ann Arbor, MI) for provision of LC3b−/− mice.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
Author Contributions
G.J.M.-C. and B.B.M. designed the experiments and wrote the manuscript. Experiments were performed by G.J.M.-C., Q.M.T., A.B.P. and C.A.W. All authors approved the final version of the manuscript.
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Supplementary information
Rights and permissions
About this article

Cite this article
Martínez-Colón, G., Taylor, Q., Wilke, C. et al. Elevated prostaglandin E2 post–bone marrow transplant mediates interleukin-1β-related lung injury. Mucosal Immunol 11, 319–332 (2018). https://doi.org/10.1038/mi.2017.51
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2017.51
This article is cited by
-
Macrophages evoke autophagy of hepatic stellate cells to promote liver fibrosis in NAFLD mice via the PGE2/EP4 pathway
Cellular and Molecular Life Sciences (2022)
