;)
;)










« Prev Next »

Chromosomes
Cells package their DNA not only to protect it, but also to regulate which genes are accessed and when. Cellular genes are therefore similar to valuable files stored in a file cabinet — but in this case, the cabinet's drawers are constantly opening and closing; various files are continually being located, pulled, and copied; and the original files are always returned to the correct location.
Of course, just as file drawers help conserve space in an office, DNA packaging helps conserve space in cells. Packaging is the reason why the approximately two meters of human DNA can fit into a cell that is only a few micrometers wide. But how, exactly, is DNA compacted to fit within eukaryotic and prokaryotic cells? And what mechanisms do cells use to access this highly compacted genetic material?
What Are Chromosomes?
Cellular DNA is never bare and unaccompanied by other proteins. Rather, it always forms a complex with various protein partners that help package it into such a tiny space. This DNA-protein complex is called chromatin, wherein the mass of protein and nucleic acid is nearly equal. Within cells, chromatin usually folds into characteristic formations called chromosomes. Each chromosome contains a single double-stranded piece of DNA along with the aforementioned packaging proteins.
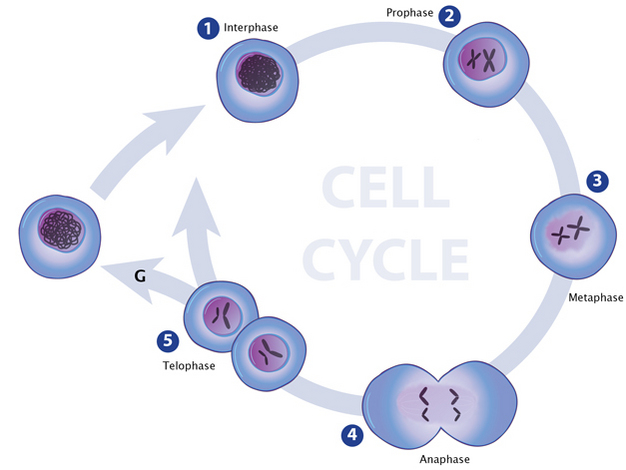
Figure 1: Chromatin condensation changes during the cell cycle.
During interphase (1), chromatin is in its least condensed state and appears loosely distributed throughout the nucleus. Chromatin condensation begins during prophase (2) and chromosomes become visible. Chromosomes remain condensed throughout the various stages of mitosis (2-5).
© 2013 Nature Education All rights reserved. 
Eukaryotes typically possess multiple pairs of linear chromosomes, all of which are contained in the cellular nucleus, and these chromosomes have characteristic and changeable forms. During cell division, for example, they become more tightly packed, and their condensed form can be visualized with a light microscope. This condensed form is approximately 10,000 times shorter than the linear DNA strand would be if it was devoid of proteins and pulled taut. However, when eukaryotic cells are not dividing — a stage called interphase — the chromatin within their chromosomes is less tightly packed. This looser configuration is important because it permits transcription to take place (Figure 1, Figure 2).
In contrast to eukaryotes, the DNA in prokaryotic cells is generally present in a single circular chromosome that is located in the cytoplasm. (Recall that prokaryotic cells do not possess a nucleus.) Prokaryotic chromosomes are less condensed than their eukaryotic counterparts and don't have easily identified features when viewed under a light microscope.
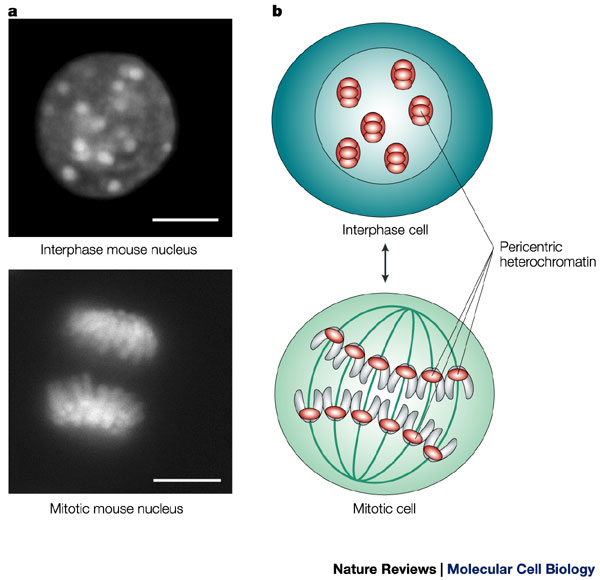
Figure 2: A the appearance of DNA during interphase versus mitosis.
During interphase, the cell's DNA is not condensed and is loosely distributed. A stain for heterochromatin (which indicates the position of chromosomes) shows this broad distribution of chromatin in a mouse cell (upper left). The same stain also shows the organized, aligned structure of the chromosomes during mitosis. Scale bars = 10 microns.
© 2004 Nature Publishing Group Maison, C. & Almouzni, G. HP1 and the dynamics of heterochromatin maintenance. Nature Reviews Molecular Cell Biology 5, 296-305 (2004). All rights reserved.
How Are Eukaryotic Chromosomes Structured?
Eukaryotic chromosomes consist of repeated units of chromatin called nucleosomes, which were discovered by chemically digesting cellular nuclei and stripping away as much of the outer protein packaging from the DNA as possible. The chromatin that resisted digestion had the appearance of "beads on a string" in electron micrographs — with the "beads" being nucleosomes positioned at intervals along the length of the DNA molecule (Figure 3).

Nucleosomes are made up of double-stranded DNA that has complexed with small proteins called histones. The core particle of each nucleosome consists of eight histone molecules, two each of four different histone types: H2A, H2B, H3, and H4. The structure of histones has been strongly conserved across evolution, suggesting that their DNA packaging function is crucially important to all eukaryotic cells (Figure 4).
Histones carry positive charges and bind negatively charged DNA in a specific conformation. In particular, a segment of the DNA double helix wraps around each histone core particle a little less than twice. The exact length of the DNA segment associated with each histone core varies from species to species, but most such segments are approximately 150 base pairs in length. Furthermore, each histone molecule within the core particle has one end that sticks out from the particle. These ends are called N-terminal tails, and they play an important role in higher-order chromatin structure and gene expression.
Figure 4: The nucleosome structure within chromatin.
Each nucleosome contains eight histone proteins (blue), and DNA wraps around these histone structures to achieve a more condensed coiled form.
© 2010 Nature Education All rights reserved.
Why Is Complex Packing Critical for Eukaryotic Chromosomes?

Figure 5: To better fit within the cell, long pieces of double-stranded DNA are tightly packed into structures called chromosomes.
© 2009 Nature Education All rights reserved.
The multiple levels of packing that exist within eukaryotic chromosomes not only permit a large amount of DNA to occupy a very small space, but they also serve several functional roles. For example, the looping of nucleosome-containing fibers brings specific regions of chromatin together, thereby influencing gene expression. In fact, the organized packing of DNA is malleable and appears to be highly regulated in cells.
Chromatin packing also offers an additional mechanism for controlling gene expression. Specifically, cells can control access to their DNA by modifying the structure of their chromatin. Highly compacted chromatin simply isn't accessible to the enzymes involved in DNA transcription, replication, or repair. Thus, regions of chromatin where active transcription is taking place (called euchromatin) are less condensed than regions where transcription is inactive or is being actively inhibited or repressed (called heterochromatin) (Figure 6).

Figure 6: The structure of chromatin in interphase
Heterochromatin is more condensed than euchromatin. Typically, the more condensed chromatin is, the less accessible it is by transcription factors and polymerases.
© 2010 Nature Education All rights reserved.
The dynamic nature of chromatin is regulated by enzymes. For example, chromatin can be loosened by changing the position of the DNA strands within a nucleosome. This loosening occurs because of chromatin remodeling enzymes, which function to slide nucleosomes along the DNA strand so that other enzymes can access the strand. This process is closely regulated and allows specific genes to be accessed in response to metabolic signals within the cell. Another way cells control gene expression is by modifying their histones with small chemical groups, such as methyl and acetyl groups in the N-terminal tails that extend from the core particle. Different enzymes catalyze each kind of N-terminal modification. Scientists occasionally refer to the complex pattern of histone modification in cells as a "histone code." Some of these modifications increase gene expression, whereas others decrease it.
How Are Chromosomes Organized in the Nucleus?
In electron micrographs, eukaryotic interphase chromatin appears much like a plate of spaghetti — in other words, there is no obvious pattern of organization. In recent years, however, investigators have begun using fluorescent probes for each of the different interphase chromosomes. In doing so, they have discovered that these chromosomes have functional and decidedly nonrandom arrangements.
One of the first things these scientists noted was that uncondensed chromosomes occupy characteristic regions of the nucleus, which they termed chromosome territories. The spatial localization of these territories is thought to be important for gene expression. In fact, with the advent of gene-specific probes, researchers are beginning to understand how the arrangement of chromosome territories can bring particular genes closer together. A second major observation related to chromosome territories is that the position of chromosomes relative to one another differs from cell to cell. Such differences reflect variation in gene expression patterns.
Conclusion
The
prokaryotic genome typically exists in the form of a circular chromosome
located in the cytoplasm. In eukaryotes, however, genetic material is
housed in
the nucleus and tightly packaged into linear chromosomes. Chromosomes
are made
up of a DNA-protein complex called chromatin that is organized into
subunits
called nucleosomes. The way in which eukaryotes compact and arrange
their
chromatin not only allows a large amount of DNA to fit in a small space,
but it
also helps regulate gene expression.
eBooks
This page appears in the following eBook












;){kind=link}