



















« Prev Next »
The Physical Environment
Life on Earth is possible primarily because light, water, and a comfortable temperature allow it to flourish. These features are part of our physical environment and part of our everyday experience. Temperature, humidity, and light vary across the Earth's surface, and from the surface skyward (see also: http://www.nature.com/scitable/knowledge/library/introduction-to-the-basic-drivers-of-climate-13368032). These changes in microclimatological conditions provide challenges and opportunities for species adapting to these changes. A very important component of the physical environment that makes life on Earth possible is consistent temperature, maintained through the natural greenhouse effect, which keeps the Earth at 20ºC on average. The natural greenhouse effect is the absorption of a part of the sun's rays by naturally abundant greenhouse gases in the atmosphere such as carbon dioxide (CO2), water vapor (H2O) and methane (CH4). However, too much of a good thing can be bad, which is why adding more CO2 to the atmosphere than is necessary to maintain the natural greenhouse effect, has become a problem.
Variations in the physical environment, such as temperature, light, and precipitation, depend upon the time of year, time of day, topography, and latitude. These variations are commonly called climate, which can be expressed on a large areal scale (e.g., the tropical climate) or on a very local scale (e.g., the climate of a forest), which we call microclimate. The distribution of light, and therefore the temperature distribution, on Earth depends upon both the tilt of the Earth's axis and the rotation of the Earth around the sun. Another feature of having a rotating Earth, and having different temperatures on Earth, is the creation of wind. Because the Earth is a sphere and because of the location of its axes, the greatest amount of radiation is received at the equator, which is then redistributed towards the poles. Wind re-distributes heat and mass, such as water vapor, carbon dioxide, and other gases, throughout the globe. This redistribution of mass and heat is the primary determinant of vegetation profiles (see biomes). Precipitation is distributed unevenly throughout the globe, and on a local scale. As biological processes depend on the physical environment, of particular interest are ecosystem scale variations in climate (i.e., microclimate).
The Physical Environment in Forests
Precipitation.
All plant growth depends on the availability of water, which in forest ecosystems is provided primarily by precipitation, but can also be provided by access to groundwater or, in riparian forest communities, to stream water. Forest canopies intercept some precipitation, and thus only 50-80% of rainfall reaches the forest floor, of which some of that evaporates before entering the soil. Of the precipitation that reaches the soil, approximately 50-80% will be transpired back into the atmosphere by the vegetation. Thus, forests serve as large water pumps between the soil and the atmosphere.
Soil.
Trees are rooted in soil, which provides nutrients and water that support tree growth. The availability of nutrients and water are dependent on soil texture (sand, silt, loam, clay content), pH, cation exchange capacity, and water holding capacity, that are determined not only by soil texture but also by organic matter content, and porosity. These properties are largely a function of the geology of a specific site, which is determined by the bedrock quality, weathered bedrock material, and organic matter incorporated into the weathered bedrock material. Depending on the soil properties, more or less nutrients or water become available to the trees. Nutrients are cycled between the biosphere and the pedosphere (soil) through tree uptake, litterfall, and decomposition. Some nutrients are re-absorbed, for example nitrogen, before the leaves are dropped. However, most nutrients are recycled into the soil via decomposition through microbes and fungi. More importantly, trees are the primary pathway by which water moves between the soil and the atmosphere. However, trees can also redistribute water within the soil. When trees have access to groundwater and the topsoil is dry, then hydraulic lift can occur. Hydraulic lift is the biologically mediated process of water redistribution through primarily plant roots (Caldwell et al. 1998, Dawson 1993), but can also be facilitated through, for example, mycorrhizal mycelia from lower moist soil layers to upper dryer soil layers. Recently, it has been also shown that roots transport water laterally in the soil to dryer soil regions (Nadezhdina et al. 2010).
Light.
Forests alter microclimatological conditions due to their height, leaf area, and density, through the absorption of radiation and cooling through transpiration. For example, in a deciduous forest canopy, light reaches the floor in the winter time, but less so in the summer time when leaves are blocking the passage of light to the ground (see Figure 1-left hand side Feb 4th and right hand side July 24th 2006 in the New Jersey Pine Barrens oak-pine forest, from Schäfer et al. 2010). This has consequences for photosynthesis, transpiration, and understory processes. Photosynthesis requires light and carbon dioxide (CO2) which enters through stomata in the leaves and needles of forest tree species. Stomata are pores that can open and close to regulate the uptake of CO2 and the loss of water (transpiration). This is a basic trade-off that all plants need to make, and highlights the critical role of stomata in controlling transpiration and photosynthesis. As light penetrates the canopy, it is attenuated (i.e., decreased). Thus leaves lower in the canopy receive less light and have to develop in such a way to maintain a positive carbon balance via photosynthesis. This is accomplished by orienting the leaves horizontally, and making them larger and thinner, which allows them to capture more light per amount of leaf material-in essence displaying shade leaf properties. Concomitant with reduced light, carboxylation efficiency, or the efficiency of carbon uptake in leaves, is also reduced (Schäfer et al. 2010). Leaf shape and placement, though, can play an important role. In coniferous canopies, due to the shoot structure and narrow leaf shape, more light penetrates, thus enhancing photosynthesis lower in the canopy and in the understory species. The effect of light bouncing in-between needles on a shoot, is referred to as the penumbral effect (Anderson & Miller 1974).
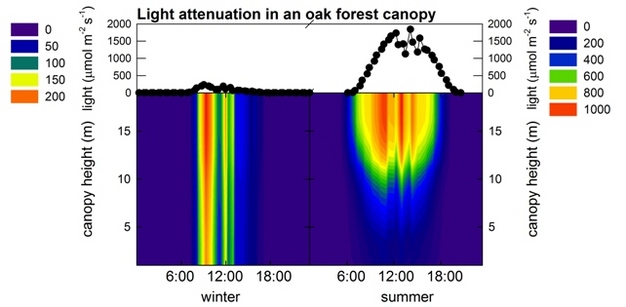
Figure 1
Top graph: light intensity (in μmol photons m-2 s-1) above the canopy. Bottom graph: light attenuation in an oak forest in early February (left hand side) and end of July (right hand side). Please note different scales for color bars. All units are in μmol photons m-2 s-1.
Wind and temperature.
Wind redistributes heat and mass, such as water vapor, carbon dioxide, and other gases in the atmosphere. Generally, in forest canopies, just like light, wind is attenuated due to the resistance of the stems, branches and leaves (see Figure 2 adapted from Clark et al. 2010). More specifically, wind speed and leaf size, shape and distribution can influence the exchange of CO2 and water at the leaf level (small scale) and at the canopy level (large scale) by affecting boundary layer conductance of leaves or canopies (Figure 3). At the leaf level, the boundary layer is a very thin layer of stagnant air surrounding the surface of a leaf. The thinner it is the less resistance it offers to the movement of water and CO2 molecules. One way in which plants can reduce their leaf boundary layer is having narrow leaves, such as pine needles, or leaves with large indentations, such as pin oak leaves. Wind increases leaf boundary layer conductance by reducing the thickness (and therefore the resistance) of the boundary layer as wind speed increases.
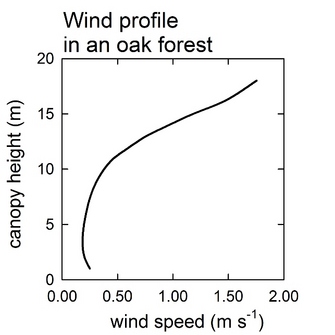
Figure 2
Wind profile through an oak forest in the New Jersey Pine Barrens throughout a year.
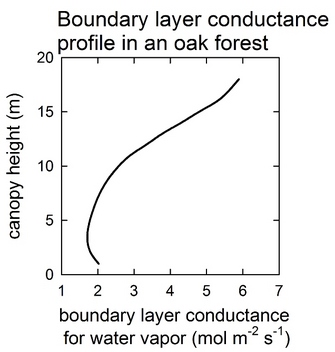
Figure 3
Boundary layer conductance profile through an oak forest canopy in the New Jersey Pine Barrens.
The boundary layer of the canopy is determined by the shape, density, and aerial spread of the crowns of trees in a forest. Together these qualities of the canopy are the roughness. Widely spread conifers with pointy tops have a higher roughness than oaks with round tops and denser packed leaves. The rougher the canopy, the greater the wind turbulence it creates, and the more wind (and therefore CO2 and water vapor) moves between the leaves of the trees. Thus, conifers tend to have thinner leaf and canopy boundary layers, and oaks tend to have thicker leaf and canopy boundary layers. When boundary layers are thin, leaves and the air surrounding them are more coupled (i.e., the wind has a larger impact on leaf physiology), and when the boundary layers are thicker, the leaves and the surrounding air are decoupled (less direct impact on leaf physiology).
The impact of the boundary layer on leaf physiology is linked to the vapor pressure gradient, which is the difference in water vapor concentration between the inside of the leaf and the air surrounding the leaf. A thicker boundary layer reduces the vapor pressure gradient by increasing the resistance to water vapor movement. Less water vapor movement away from the leaf means less evaporative cooling, and the leaves get warmer. If leaves get too warm, the process of photosynthesis breaks down, and the tree suffers. Thus, oak leaves at the bottom of the canopy can be wider (which increases their boundary layer thickness) and horizontally oriented (which increases their light absorption) even though there is less wind, because there is less light at the bottom of the canopy and therefore less heating of the leaves. At the top, the leaves need to be narrower and more vertically oriented to reduce the boundary layer and heat absorption. In conifers, because of the way the leaves and crowns are constructed, all the leaves have thin boundary layers. The thinner the boundary layer, the more important stomatal control of water loss becomes, because water vapor is more likely to move out of the leaf (greater vapor pressure gradient).
Jarvis & McNaughton (1986) have quantified the influence of the atmosphere on gas exchange in forest canopies with a decoupling factor Ω. It describes the linkage of the physical environment with the biological processes in plant canopies and is defined as
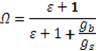
The decoupling coefficient Ω is unitless, and falls between 0 and 1. The rougher the canopy, the greater boundary conductance (gb) becomes, the closer Ω is to 0, the more coupled the canopy is to the atmosphere, and stomatal control becomes more important. In contrast, the less rough the canopy, the smaller the boundary layer conductance becomes, Ω is closer to 1, the canopy is decoupled from the atmosphere, and stomata conductance has less impact on transpiration.
Summary
The physical environment of forests is determined by edaphic (soil) factors and micro-climate (precipitation, light, temperature, and wind). Gas-exchange within forest canopies depends upon the microclimatic condition within the forest, as well as, edaphic factors and water availability. Current changes in the global physical environment will undoubtedly alter the physical environment of forests. Changes in the physical environment will affect gas-exchange and thereby the capacity of forests to remove carbon dioxide from the atmosphere. The interactions between various physical factors and forest gas-exchange will feedback to microclimatic conditions as lower transpiration rates decrease humidity or higher transpiration rates increase humidity. Thus, forest ecosystems are a part of the physical environmental feedback loop and therefore our understanding of these interactions is crucial in order to predict forest growth, species survival, mass and heat exchange in the forest, and the ensuing microclimatic conditions.
Glossary
Biomes - Large scale units of similar vegetation and climate.
Boundary layer - Resistive layer of air on the surface of leaves that varies with wind speed and leaf size, and controls transpiration.
Carboxylation efficiency - Efficiency of the major photosynthesis enzyme RUBISCO to assimilate CO2Cation exchange capacity - Capacity of soil particles to provide cations, such as potassium (K), sodium (Na), magnesium (Mg), calcium (Ca), and other important plant micro- and macro-nutrients.
Climate - Long term average of temperature, humidity, air pressure, wind speed and direction.
Decomposition - The mechanical, chemical and biological breakdown of organic material over time.
Hydraulic lift - The biological process of redistributing water through plant roots into different soil volumes/layers.
Microclimate - Environmental conditions on a small scale, such as around a leaf or a plant.
Pedosphere - The entirety of soil, outermost layer of the Earth's crust (pedos means soil in Greek).
Penumbral effect - The shadow-edge effect whereby light that penetrates through openings creates an increased light level at the edge (e.g., through canopy gaps).
pH - Hydroxyl (OH-) ion concentration in a pure water solution displayed as the negative logarithm with base ten, i.e., 10-7 = pH 7, which is neutral.
Porosity - The amount of space in a given soil volume that is filled either with air or water.
Riparian - Edge of rivers, land-water interface.
Shade leaf - A leaf displaying anatomical, morphological, and physiological conditions due to low light conditions the leaves developed under. Shade leaves are larger and thinner than sun leaves (leaves developing under full sunlight).
Soil texture - Grain fractions in soil such as sand (0.05 - 2mm), silt (0.002-0.05 mm), and clay (<0.002 mm).
Stomata - Pore opening on surface of plant leaves commonly at the bottom of the leaf but also in some plant species at the top or on both sides of the leaf. Gateway of gas-exchange between the plant and the atmosphere.
Vapor pressure gradient - A measure of water vapor concentration difference between inside of the leaf and the air.
Water holding capacity - The ability of the soil to hold water against gravity.
Weather - Day to day variations in temperature, humidity, air pressure, wind speed and direction.
Weathering - The breakdown of rocks into smaller fractions such as sand, silt, and clay. Weathering can occur through physical/mechanical (ice, movement), chemical (water, acids) and biological (through root exudates) means.
Ω - Decoupling coefficient between atmosphere and biosphere (leaves) that ranges between 0 and 1. The closer to 1, the more decoupled leaves are from the atmosphere, as is the case for large leaves under the full sun and low wind conditions. In this case radiation drives transpiration, not the vapor pressure gradient.
References and Recommended Reading
Anderson, M. C. & Miller, E. E. Forest cover as a solar camera: Penumbral effects in plant canopies. Journal of Applied Ecology 11, 691-697 (1974).
Caldwell, M. M., Dawson, T. E. & Richards, J. H. Hydraulic lift: Consequences of water efflux from the roots of plants. Oecologia 113, 151-161 (1998).
Campbell, G. & Norman, J. M. An Introduction to Environmental Biophysics. New York, NY: Springer-Verlag, 1998.
Clark, K. L., Skowronski, N. & Hom, J. Invasive insects impact forest carbon dynamics. Global Change Biology 16, 88-101 (2010).
Dawson, T. E. Hydraulic lift and water use by plants: Implications for water balance, performance and plant-plant interactions. Oecologia 95, 565-574 (1993).
Jarvis, P. G. Coupling of carbon and water interactions in forest stands. Tree Physiology 2, 347-368 (1986).
Jarvis, P. G. & Mcnaughton, K. G. Stomatal control of transpiration: Scaling up from leaf to region. Advances in Ecological Research 15, 1-49 (1986).
Nadezhdina, N. et al. Trees never rest: The multiple facets of hydraulic redistribution. Ecohydrology 3, 431-444 (2010).
Schäfer, K. V. R. et al. Impact of insect defoliation on forest carbon balance as assessed with a canopy assimilation model. Global Change Biology 16, 546-560 (2010).










