


















« Prev Next »
Introduction
Food web is an important ecological concept. Basically, food web represents feeding relationships within a community (Smith and Smith 2009). It also implies the transfer of food energy from its source in plants through herbivores to carnivores (Krebs 2009). Normally, food webs consist of a number of food chains meshed together. Each food chain is a descriptive diagram including a series of arrows, each pointing from one species to another, representing the flow of food energy from one feeding group of organisms to another.
There are two types of food chains: the grazing food chain, beginning with autotrophs, and the detrital food chain, beginning with dead organic matter (Smith & Smith 2009). In a grazing food chain, energy and nutrients move from plants to the herbivores consuming them, and to the carnivores or omnivores preying upon the herbivores. In a detrital food chain, dead organic matter of plants and animals is broken down by decomposers, e.g., bacteria and fungi, and moves to detritivores and then carnivores.
Food web offers an important tool for investigating the ecological interactions that define energy flows and predator-prey relationship (Cain et al. 2008). Figure 1 shows a simplified food web in a desert ecosystem. In this food web, grasshoppers feed on plants; scorpions prey on grasshoppers; kit foxes prey on scorpions. While the food web showed here is a simple one, most feed webs are complex and involve many species with both strong and weak interactions among them (Pimm et al. 1991). For example, the predators of a scorpion in a desert ecosystem might be a golden eagle, an owl, a roadrunner, or a fox.
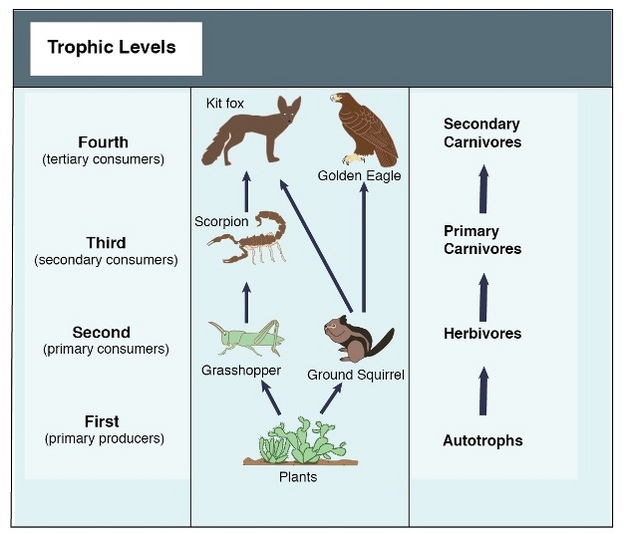
Figure 1: A simple six-member food web for a representative desert grassland.
© 2012 Nature Education Adapted from Cain et al. 2008. All rights reserved. 
The idea to apply the food chains to ecology and to analyze its consequences was first proposed by Charles Elton (Krebs 2009). In 1927, he recognized that the length of these food chains was mostly limited to 4 or 5 links and the food chains were not isolated, but hooked together into food webs (which he called "food cycles"). The feeding interactions represented by the food web may have profound effects on species richness of community, and ecosystem productivity and stability (Ricklefs 2008).
Types of Food Webs
Food webs describe the relationships — links or connections — among species in an ecosystem, but the relationships vary in their importance to energy flow and dynamics of species populations. Some trophic relationships are more important than others in dictating how energy flows through ecosystems. Some connections are more influential on species population change. Based on different ways in which species influence one another, Robert Paine proposed three types of food webs based on the species of a rocky intertidal zone on the coast of Washington (Ricklefs 2008, Figure 2). Connectedness webs (or topological food webs) emphasize feeding relationships among species, portrayed as links in a food web (Paine 1980). Energy flow webs quantify energy flow from one species to another. Thickness of an arrow reflects the strength of the relationship. Functional webs (or interaction food webs) represent the importance of each species in maintaining the integrity of a community and reflect influence on the growth rate of other species' populations. As shown in Figure 2, limpets Acmaea pelta and A. mitra in the community consume considerable food energy (energy flow web), but removal of these consumers has no detectable influence on the abundance of their resources (functional web). The most effective control was exerted by sea urchin Stronglocentrotus and the chiton Katharina (Ricklefs 2008).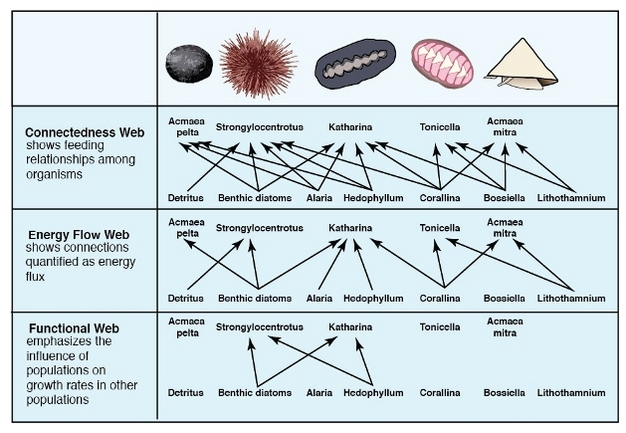
Figure 2: Three types of food web diagrams based on species of a rocky intertidal zone on the coast of Washington.
© 2012 Nature Education All rights reserved.
Applications of Food Webs
Food webs are constructed to describe species interactions (direct relationships).
The fundamental purpose of food webs is to describe feeding relationship among species in a community. Food webs can be constructed to describe the species interactions. All species in the food webs can be distinguished into basal species (autotrophs, such as plants), intermediate species (herbivores and intermediate level carnivores, such as grasshopper and scorpion) or top predators (high level carnivores such as fox) (Figure 1).
These feeding groups are referred as trophic levels. Basal species occupy the lowest trophic level as primary producer. They convert inorganic chemical and use solar energy to generate chemical energy. The second trophic level consists of herbivores. These are first consumers. The remaining trophic levels include carnivores that consume animals at trophic levels below them. The second consumers (trophic level 3) in the desert food web include birds and scorpions, and tertiary consumers making up the fourth trophic level include bird predators and foxes. Grouping all species into different functional groups or tropic levels helps us simplify and understand the relationships among these species.
Food webs can be used to illustrate indirect interactions among species.
Indirect interaction occurs when two species do not interact with each other directly, but influenced by a third species. Species can influence one another in many different ways. One example is the keystone predation are demonstrated by Robert Paine in an experiment conducted in the rocky intertidal zone (Cain et al. 2008; Smith & Smith 2009; Molles 2010). This study showed that predation can influence the competition among species in a food web. The intertidal zone is home to a variety of mussels, barnacles, limpets, and chitons (Paine 1969). All these invertebrate herbivores are preyed upon by the predator starfish Pisaster (Figure 3). Starfish was relatively uncommon in the intertidal zone, and considered less important in the community. When Paine manually removed the starfish from experimental plots while leaving other areas undisturbed as control plots, he found that the number of prey species in the experimental plots dropped from 15 at the beginning of the experiment to 8 (a loss of 7 species) two years after the starfish removal while the total of prey species remained the same in the control plots. He reasoned that in the absence of the predator starfish, several of the mussel and barnacle species (that were superior competitors) excluded the other species and reduced overall diversity in the community (Smith & Smith 2009). Predation by starfish reduced the abundance of mussel and opened up space for other species to colonize and persist. This type of indirect interaction is called keystone predation.
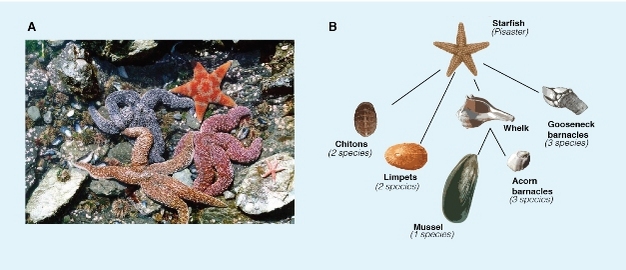
Figure 3
(a) The rocky intertidal zone of the Pacific Northwest coast is inhabited by a variety of species including starfish, barnacles, limpets, chitons, and mussels. (b) A food web of this community shows that the starfish preys on a variety of invertebrate species. Removal of starfish from this community reduced the diversity of prey species due to increased competition.
© 2012 Nature Education All rights reserved.
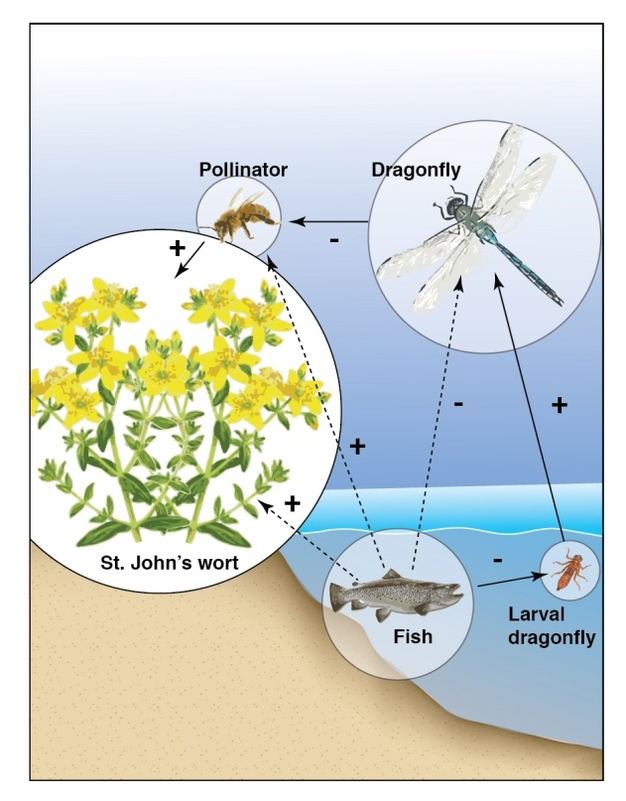
Figure 4: An interaction food web shows that fish have indirect effects on the populations of several species in and around ponds.
The solid arrows represent direct effects, and the dashed arrows indirect effects; the nature of the effect is indicated by + or -. Fish have indirect effects, through a trophic cascade, on several terrestrial species: dragonfly adults (-), pollinators (+), and plants (+)
© 2012 Nature Education All rights reserved.
Food webs can be used to study bottom-up or top-down control of community structure.
Food webs illustrate energy flow from primary producers to primary consumers (herbivores), and from primary consumers to secondary consumers (carnivores). The structure of food webs suggests that productivity and abundance of populations at any given trophic level are controlled by the productivity and abundance of populations in the trophic level below them (Smith & Smith 2009). This phenomenon is call bottom-up control. Correlations in abundance or productivity between consumers and their resources are considered as evidence for bottom-up control. For example, plant population densities control the abundance of herbivore populations which in turn control the densities of the carnivore populations. Thus, the biomass of herbivores usually increases with primary productivity in terrestrial ecosystems.
Top-down control occurs when the population density of a consumer can control that of its resource, for example, predator populations can control the abundance of prey species (Power 1992). Under top-down control, the abundance or biomass of lower trophic levels depends on effects from consumers at higher trophic levels. A trophic cascade is a type of top-down interaction that describes the indirect effects of predators. In a trophic cascade, predators induce effects that cascade down the food chain and affect biomass of organisms at least two links away (Ricklefs 2008). Nelson Hairston, Frederick Smith and Larry Slobodkin first introduced the concept of top-down control with the frequently quoted "the world is green" proposition (Power 1992; Smith & Smith 2009). They proposed that the world is green because carnivores depress herbivores and keep herbivore populations in check. Otherwise, herbivores would consume most of the vegetation. Indeed, a bird exclusion study demonstrated that there were significantly more insects and leaf damage in plots without birds compared to the control (Marquis & Whelan 1994).
Food webs can be used to reveal different patterns of energy transfer in terrestrial and aquatic ecosystems.
Patterns of energy flow through different ecosystems may differ markedly in terrestrial and aquatic ecosystems (Shurin et al. 2006). Food webs (i.e., energy flow webs) can be used to reveal these differences. In a review paper, Shurin et al. (2006) provided evidence for systematic difference in energy flow and biomass partitioning between producers and herbivores, detritus and decomposers, and higher trophic levels in food webs. A dataset synthesized by Cebrian and colleagues on the fate of carbon fixed by primary productivity across different ecosystems was used to show different patterns in food chains between terrestrial and aquatic ecosystems (Figure 5). On average, the turnover rate of phytoplankton is 10 to 1000 times faster than that of grasslands and forests, thus, less carbon is stored in the living autotroph biomass pool, and producer biomass is consumed by aquatic herbivores at 4 times the terrestrial rate (Cebrian 1999, 2004; Shurin et al. 2006). Herbivores in terrestrial ecosystems are less abundant but decomposers are much more abundant than in phytoplankton dominated aquatic ecosystems. In most terrestrial ecosystems with high standing biomass and relatively low harvest of primary production by herbivores, the detrital food chain is dominant (Smith & Smith 2009). In deep-water aquatic ecosystems, with their low standing biomass, rapid turnover of organisms, and high rate of harvest, the grazing food chain may be dominant.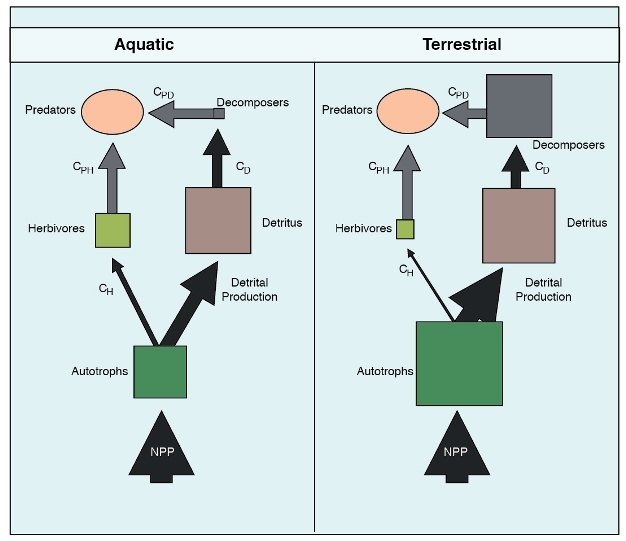
Figure 5: Differences in pathways of carbon flow and pools between aquatic and terrestrial ecosystems.
The thickness of the arrows (flows) and the area of the boxes (pools) correspond to the magnitude. The size of the pools are scaled as log units since the differences cover four orders of magnitude. The C’s indicate consumption terms (i.e. CH is consumption by herbivores). Ovals and arrows in grey indicate unknown quantities.
© 2012 Nature Education All rights reserved.
As a diagram tool, food web has been approved to be effective in illustrating species interactions and testing research hypotheses. It will continue to be very helpful for us to understand the associations of species richness/diversity with food web complexity, ecosystem productivity, and stability.
References and Recommended Reading
Cain, M. L., Bowman, W. D. & Hacker, S. D. Ecology. Sunderland MA: Sinauer Associate Inc. 2008.
Cebrian, J. Patterns in the fate of production in plant communities. American Naturalist 154, 449-468 (1999)
Cebrian, J. Role of first-order consumers in ecosystem carbon flow. Ecology Letters 7, 232-240 (2004)
Elton, C. S. Animal Ecology. Chicago, MI: University of Chicago Press, 1927, Republished 2001.
Knight, T. M., et al. Trophic cascades across ecosystems. Nature 437, 880-883 (2005)
Krebs, C. J. Ecology 6th ed. San Francisco CA: Pearson Benjamin Cummings, 2009.
Marquis, R. J. & Whelan, C. Insectivorous birds increase growth of white oak through consumption of leaf-chewing insects. Ecology 75, 2007-2017 (1994)
Molles, M. C. Jr. Ecology: Concepts and Applications 5th ed. New York, NY: McGraw-Hill Higher Education, 2010.
Paine, R. T. The Pisaster-Tegula interaction: Prey parches, predator food preferences and intertide community structure. Ecology 60, 950-961 (1969)
Paine, R. T. Food web complexity and species diversity. The American Naturalist 100, 65-75 (1966)
Paine, R. T. Food webs: Linkage, interaction strength and community infrastructure. Journal of Animal Ecology 49, 667-685 (1980)
Pimm, S. L., Lawton, J. H. & Cohen, J. E. Food web patterns and their consequences. Nature 350, 669-674 (1991)
Power, M. E. Top-down and bottom-up forces in food webs: do plants have primacy? Ecology 73, 733-746 (1992)
Schoender, T. W. Food webs from the small to the large. Ecology 70, 1559-1589 (1989)
Shurin, J. B., Gruner, D. S. & Hillebrand, H. All wet dried up? Real differences between aquatic and terrestrial food webs. Proc. R. Soc. B 273, 1-9 (2006) doi:10.1098/rspb.2005.3377
Smith, T. M. & Smith, R. L. Elements of Ecology 7th ed. San Francisco CA: Pearson Benjamin Cummings, 2009.










