


















« Prev Next »

Everyone is familiar with the classic dilemma of the chicken and the egg: which came first? In the field of ecology, a similar conundrum exists between the cause and effect relationship between disturbance and diversity. On the one hand, disturbance can clearly influence community-level diversity (Connell 1978; Petraitis et al. 1989; Sousa 2001). On the other hand, diversity can also mediate disturbance severity, leading to variation in the frequency, intensity, or duration, of actual biomass loss (i.e., realized disturbance) in communities with different initial diversity (Pimm 1984; Tilman 1996; Mulder et al. 2001, Cardinale & Palmer 2002; Allison 2004).
Disturbance Affects Species Diversity
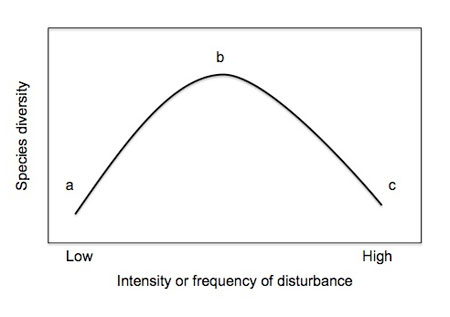
Figure 1: The intermediate disturbance hypothesis (IDH)
(A) Species diversity is low at low disturbance frequency because of competitive exclusion. (B) Species diversity is higher at intermediate disturbance frequency due to a mix of good colonizer and good competitor species. (C) Species diversity is low at high disturbance frequency because only good colonizers or highly tolerant species can persist.
Marine macroalgal communities provide a classic example of the IDH, with numerous studies illustrating a hump-shaped relationship between physical or biological disturbance and algal species diversity (Lubchenco 1978; Sousa 1979; Carpenter 1981; Sousa 2001). For example, Sousa (1979) studied the species diversity of algae growing on boulders in the marine intertidal zone. These boulders get overturned due to the physical force of wave action, and algae growing on the boulders get crushed and/or smothered as a result. Because the frequency of boulder movement is inversely proportional to boulder size (i.e., small boulders overturn frequently, whereas large boulders do not), boulder size can be used as a proxy for disturbance frequency. As predicted by the IDH:
- species diversity is low on small, frequently disturbed boulders, consisting primarily of fast-growing colonizer species;
- large boulders are dominated by competitively superior algae, leading to low diversity;
- diversity is maximized on intermediate boulders, with a mix of colonizers, competitive dominants, and species that lie in between these two extremes.
By experimentally stabilizing small boulders, Sousa (1979) demonstrated that the physical disturbance caused by boulders overturning with different frequency leads to variation in species diversity, rather than some other difference between boulders of different sizes.
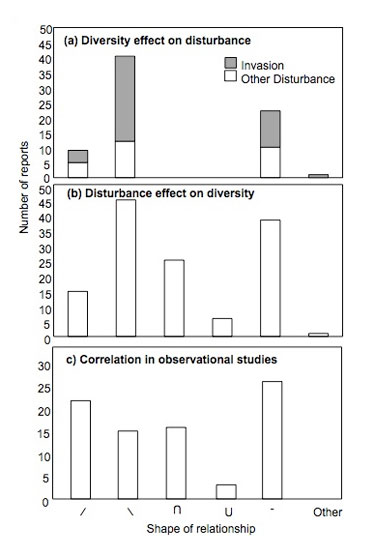
Figure 2: Summary of published relationships between disturbance and diversity
(A) The effect of diversity on realized disturbance in experimental manipulations. (B) The effect of disturbance on diversity in experimental manipulations. (C) The shape of the disturbance (horizontal axis) and diversity (vertical axis) relationship found in correlation studies. / indicates a positive relationship; indicates a negative relationship; the upside-down U indicates a unimodal relationship, peaking at intermediate values; U indicates a unimodal relationship peaking at either extreme; - indicates no significant relationship; ‘Other’ indicates some other form of the relationship (Hughes et al. 2007).
Disturbance significantly affects diversity in a majority of experimental manipulations examining the two factors (~70%; Hughes et al. 2007). However, this relationship can take a variety of shapes other than the unimodal pattern predicted by the IDH (Mackey & Currie 2001, Hughes et al. 2007; Figure 2). For instance, in marine soft sediment communities, competitive exclusion is rare because organisms are able to partition space in three-dimensions rather than just two (Summerson & Peterson 1984); thus the "dip" in sessile species diversity at low disturbance frequency predicted by the IDH does not occur. Similarly, the IDH often does not apply to mobile species because of reduced competitive exclusion, particularly in communities characterized by high habitat complexity (and thus increased niche space) at low disturbance frequency (Bruno et al. 2003, Kimbro & Grosholz 2006). Consistent with these examples, disturbance most commonly decreases species diversity as it increases in frequency or severity in experimental manipulations (Hughes et al. 2007).
Species Diversity Affects Resistance and Resilience to Disturbance
In addition to strong evidence for disturbance effects on diversity, both theory (MacArthur 1955; May 1973; Yachi & Loreau 1999, Loreau et al. 2003) and experiments (Tilman 1996; Mulder et al. 2001; Cardinale & Palmer 2002; Allison 2004; Hughes & Stachowicz 2004) demonstrate that diversity can affect community stability by influencing the response to disturbance and/or environmental fluctuations. For instance, more diverse assemblages are more likely to display a range of functional traits, increasing the probability that one species can compensate for the negative responses of other species to disturbance or environmental change (the insurance hypothesis; e.g., Tilman 1996). A related mechanism is the portfolio effect: if the abundance of different species fluctuates independently, or at least out of phase with one another, then these fluctuations will average each other out, leading to less variation over time in a diverse assemblage (Doak et al. 1998)

Figure 3: Drought resistance in terrestrial grasslands
Resistance to drought, defined as the relative rate of plant community change in biomass from pre-drought to peak drought, is a saturating function of pre-drought plant species richness (Tilman & Downing 1994).
Studies of terrestrial grasslands provide compelling evidence that increased species diversity can reduce variation in community biomass over time, particularly in response to drought (Tilman & Downing 1994; Tilman 1996; Figure 3). Plant species diversity increases resistance to drought in this system, with diverse plots losing less biomass following severe drought (Tilman & Downing 1994). This positive result of diversity occurred because more diverse plots had a higher probability of including a drought resistant species that could compensate for the biomass loss of less resistant species (i.e., the insurance hypothesis; Tilman & Downing 1994). Although there are undoubtedly some environmental fluctuations that are sufficient to overcome the capacity of diversity to respond, a general consensus emerging from studies of biodiversity is that diversity commonly decreases realized disturbance (Figure 2), and reduces variability in community traits (e.g., biomass and productivity) over time (Balvanera et al. 2006; Worm et al. 2006; Hughes et al. 2007; Stachowicz et al. 2007)
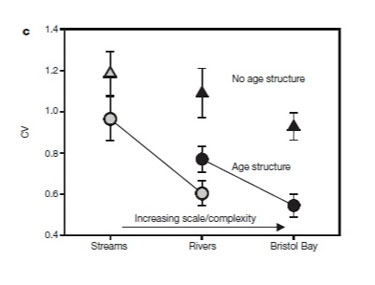
Figure 4: Effects of salmon population diversity on catch variation in Bristol Bay, AK
The coefficient of variation from year to year in total sockeye salmon returns at three spatial scales (streams to rivers to bay-wide) and two levels of life history aggregation (circles represent age structured data and triangles represent variation within dominant age classes). Gray symbols are from the Wood River system, with continuous long-term data on stream populations. Black symbols are for all rivers in Bristol Bay (Schindler et al. 2010).
As with the effects of disturbance on species diversity, the shape of the relationship between diversity and disturbance response can vary (Figure 2), although diversity most commonly reduces realized disturbance (Hughes et al. 2007). Because experimental tests of the effects of diversity typically manipulate common species in the community of interest, they may actually underestimate the impact of diversity: uncommon species can play an important role in the response to disturbance and environmental change (Lyons et al. 2005; Duffy 2009). Future studies need to incorporate variation in relative abundance as well as diversity to tease apart these effects.
Do Disturbance and Diversity Interact?
The role of rare species provides one example of the chicken and egg nature of the diversity-disturbance relationship: disturbance can promote the coexistence of less common species by preventing competitive exclusion (e.g., Sousa 1979), and less common species can strongly influence the response of communities to disturbance (Lyons et al. 2005). Thus, the diversity-disturbance relationship may be more cyclical than unidirectional, with both directions of causation operating simultaneously: disturbance changes diversity, which changes the response of the community to future disturbance, and so on (Hughes et al. 2007).
Several lines of evidence support the suggestion that there are reciprocal effects of disturbance and diversity. First, as detailed above, there is substantial evidence for each of the one-way relationships (disturbance affects diversity and diversity affects realized disturbance). In addition, each of these one-way relationships has been documented in the same system at similar spatial scales (e.g., terrestrial grasslands and marine macroalgal communities; Hughes et al. 2007), suggesting that there is no a priori reason to believe that one direction of causation would necessarily dominate the other. Finally, discrepancies between experimental studies and observations in unmanipulated systems suggest that it is difficult to separate the direction of causation between diversity and disturbance: disturbance most often reduces diversity in experiments, but a positive relationship between the two is commonly found in observational studies (Mackey & Currie 2001; Hughes et al. 2007; Figure 2).
Although the point of dual causation between diversity and disturbance may appear both obvious and esoteric, failing to consider the reciprocal effects of one upon the other could hinder management and conservation decisions. For instance, sockeye salmon population diversity clearly reduces variability in catch (Hilborn et al. 2003; Schindler et al. 2010), sustaining higher fishing pressure than would be possible in the absence of this diversity. Conversely, if fishing pressure, land and hydrological alteration, and climate change continue to reduce population diversity (Gustafson et al. 2007), then catch will likely decline more than expected based on predictions from a single, homogeneous population. Thus considerations of diversity and its stabilizing effects are necessary for effective management of this system.
It turns out that this chicken and egg issue is not limited to the relationship between disturbance and diversity; in fact, there are a variety of ecological processes that are both a cause and a consequence of biodiversity. For example, productivity, or resource availability, can be an important determinant of community diversity (MacArthur 1965; Loreau et al. 2001; Chase & Leibold 2002), and manipulations of species diversity reveal strong effects on community productivity (Balvanera et al. 2006; Cardinale et al. 2006). Resolving the nature and direction of the diversity-productivity relationship has thus been the focus of numerous studies (Loreau et al. 2001; Chase & Leibold 2002; Schmid 2002). A similar example comes from studies of diversity and species invasion: increased diversity in experimental manipulations typically leads to reduced invasion success (Levine et al. 2004; Stachowicz & Byrnes 2006); whereas invasion, by definition, increases community diversity by the addition of the invasive species, it may lead to additional changes in diversity over time due to positive and negative effects of invaders on natives (Bruno et al. 2005; Rodriguez 2006). Though not the focus of this article, these examples support recent suggestions that reciprocal (i.e., chicken and egg) interactions are likely common in ecology (Agrawal et al. 2007).
References and Recommended Reading
Agrawal, A. A., Ackerly, D. D. et al. Filling key gaps in population and community ecology. Frontiers in Ecology and the Environment 5, 145-152 (2007).
Allison, G. The influence of species diversity and stress intensity on community resistance and resilience. Ecological Monographs 74, 117-134 (2004).
Balvanera, P., Pfisterer, A. B. et al. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters 9, 1146-1156 (2006).
Bruno, J. F., Fridley, J. D. et al. In Species Invasions: Insights into Ecology, Evolution, and Biogeography, eds. Sax, D. F., Stachowicz, J. J. et al. (Sunderland: Sinauer Associates, 2005): 13-40.
Bruno, J. F., Stachowicz, J. J. et al. Inclusion of facilitation into ecological theory. Trends In Ecology & Evolution 18, 119-125 (2003).
Cardinale, B. J. & Palmer, M. A. Disturbance moderates biodiversity-ecosystem function relationships: experimental evidence from Caddisflies in stream mesocosms Ecology 83, 1915-1927 (2002).
Cardinale, B. J., Srivastava, D. S. et al. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 443, 989-992 (2006).
Carpenter, R. C. Grazing by Diadema antillarum (Phillips) and its effects on the benthic algal community. Journal of Marine Research 39, 749-765 (1981).
Chase, J. M. & Leibold, M. A. Spatial scale dictates the productivity-biodiversity relationship. Nature 416, 427-430 (2002).
Connell, J. H. Diversity in tropical rain forests and coral reefs. Science 199, 1302-1310 (1978).
Doak, D. F., Bigger, D. et al. The statistical inevitability of stability-diversity relationships in community ecology. American Naturalist 151, 264-276 (1998).
Duffy, J. E. Why biodiversity is important to the functioning of real-world ecosystems. Frontiers in Ecology and the Environment 7, 437-444 (2009).
Grime, J. P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist 111, 1169-1194 (1977).
Gustafson, R. G., Waples, R. S. et al. Pacific salmon extinctions: quantifying lost and remaining diversity. Conservation Biology 21, 1009-1020 (2007).
Hilborn, R., Quinn, T. P. et al. Biocomplexity and fisheries sustainability. Proceedings of the National Academy of Sciences, USA 100, 6564-6568 (2003).
Hughes, A. R., Byrnes, J. E. et al. Reciprocal relationships and potential feedbacks between biodiversity and disturbance. Ecology Letters 10, 849-864 (2007).
Hughes, A. R., Inouye, B. D. et al. Ecological consequences of genetic diversity. Ecology Letters 11, 609-623 (2008).
Hughes, A. R. & Stachowicz, J. J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proceedings of the National Academy of Sciences, USA 101, 8998-9002 (2004).
Huston, M. A general hypothesis of species diversity. American Naturalist 113, 81-101 (1979).
Kimbro, D. L. & Grosholz, E. D. Disturbance influences oyster community richness and evenness, but not diversity. Ecology 87, 2378-2388 (2006).
Levine, J. M., Adler, P. B. et al. A meta-analysis of biotic resistance to exotic plant invasions. Ecology Letters 7, 975-989 (2004).
Loreau, M., Mouquet, N. et al. Biodiversity as spatial insurance in heterogeneous landscapes. Proceedings of the National Academy of Sciences, USA 100, 12765-12770 (2003).
Loreau, M., Naeem, S. et al. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294, 804-808 (2001).
Lubchenco, J. Plant species diversity in a marine intertidal community: importance of herbivore food preference and algal competitive abilities. American Naturalist 112, 23-39 (1978).
Lyons, K. G., Brigham, C. A. et al. Rare species and ecosystem functioning. Conservation Biology 19, 1019-1024 (2005).
MacArthur, R. H. Fluctuations of animal populations and a measure of community stability. Ecology 36, 533-536 (1955).
MacArthur, R. H. Patterns of species diversity. Biological Reviews 40, 510-533 (1965).
Mackey, R. L. & Currie, D. J. The diversity-disturbance relationship: is it generally strong and peaked? Ecology 82, 3479-3492 (2001).
May, R. M. Stability and Complexity in Model Ecosystems. Princeton, NJ: Princeton University Press, 1973.
Mulder, C. P. H., Uliassi, D. D. et al. Physical stress and diversity-productivity relationships: the role of positive interactions. Proceedings of the National Academy of Sciences, USA 98, 6704-6708 (2001).
Petraitis, P. S., Latham, R. E. et al. The maintenance of species diversity by disturbance. The Quarterly Review of Biology 64, 393-418 (1989).
Pimm, S. L. The complexity and stability of ecosystems. Nature 307, 321-326 (1984).
Rodriguez, L. F. Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biological Invasions 8, 927-939 (2006).
Schindler, D. E., Hilborn, R., et al. Population diversity and the portfolio effect in an exploited species. Nature 465, 609-613 (2010).
Schmid, B. The species richness-productivity controversy. Trends In Ecology & Evolution 17, 113-114 (2002).
Sousa, W. P. Distubance in marine intertidal boulder fields: the nonequilibrium maintenance of species diversity. Ecology 60, 1225-1239 (1979).
Sousa, W. P. "Natural disturbance and the dynamics of marine benthic communities," in Marine Community Ecology, eds. M. D. Bertness, S. D. Gaines, et al. (Sunderland, MA: Sinauer Associates, 2001): 85-130.
Stachowicz, J. J., Bruno, J. F., et al. Understanding the effects of marine biodiversity on communities and ecosystems. Annual Review of Ecology, Evolution and Systematics 38, 739-766 (2007).
Stachowicz, J. J. & Byrnes, J. E. Species diversity, invasion success and ecosystem functioning: disentangling the influence of resource competition, facilitation, and extrinsic factors. Marine Ecology Progress Series 311, 251-262 (2006).
Summerson, H. C. & Peterson, C. H. Role of predation in organizing benthic communities of a temperate-zone seagrass bed. Marine Ecology Progress Series 15, 63-77 (1984).
Tilman, D. Biodiversity: population versus ecosystem stability. Ecology 77, 350-363 (1996).
Tilman, D. & Downing, J. A. Biodiversity and stability in grasslands. Nature 367, 363-365 (1994).
Worm, B., Barbier, E. B., et al. Impacts of biodiversity loss on ocean ecosystem services. Science 314, 787-790 (2006).
Yachi, S. & Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proceedings of the National Academy of Sciences, USA 96, 1463-1468 (1999).










