


















« Prev Next »

The topic of what to do with the Middle Pleistocene hominin fossils that cannot readily be placed in the modern Homo sapiens hypodigm, both morphologically and behaviorally, but yet cannot be easily allocated to H. erectus sensu lato has been long debated by paleoanthropologists (e.g., Binford, 1985; Tattersall 1986; Wolpoff et al. 1994; Rightmire 1998, 2008; Brauer 2008; Tattersall & Schwartz 2008; Bae 2010; Stringer 2012). Traditionally, these fossils have been referred to as archaic Homo sapiens, though other names have been used such as Mid-Pleistocene Homo, and Homo heidelbergensis, as well as others with a more regional focus: H. rhodesiensis (Africa), H. soloensis (Indonesia), H. daliensis (North China), H. mabaensis (South China). This brief review will cover the general characteristics of archaic H. sapiens morphology, paleoecology, and behavior.
Archaic Homo sapiens (morphology)
Most paleoanthropologists agree that Homo erectus and H. sapiens are distinct species (but see Wolpoff et al. 1994). The primary morphological characters of H. erectus are a long and low cranium, a pronounced supraorbital torus, a pronounced postorbital constriction, an angled occipital torus, a cranium that is widest at the base, the absence of a chin, and a cranial capacity of about 1,000 cc. The primary morphological traits of H. sapiens that distinguish it from H. erectus are a more rounded and high cranium, a reduced supraorbital torus, the lack of a postorbital constriction, a less angled occipital torus, a cranium that is widest at the top of the parietals rather than at the base, a chin, and a cranial capacity of about 1,350 cc. Homo erectus postcranial fossils are not frequently reported, but studies do suggest that in terms of overall size, the H. erectus postcrania fall within the range of modern H. sapiens, but generally are more robust. The primary distinctions between the two species appear to be morphological variation found in the crania.
A wide range of Middle Pleistocene hominin fossils from different regions of the Old World that do not fit comfortably within either H. erectus s.l. or modern H. sapiens have been allocated to archaic H. sapiens. The African fossils include Bodo, Broken Hill, Ndutu, and Elandsfontein, while the western Eurasian fossils include Mauer, Arago, Petralona, Sima de los Huesos, and possibly Zuttiyeh from Israel. The primary Chinese archaic H. sapiens fossils are from Dali, Jinniushan, Chaoxian, and Maba, while the Solo fossils from Indonesia can also be included. The ages of these hominin fossils range from about 640,000 years ago (Bodo) to as recently as about120,000 years ago (Maba) (Tattersall 1986; Pope 1992; Wu & Poirier 1995; Etler 1996; Rightmire 1998; Stringer 2002; Conroy 2005; Rightmire 2008; Cartmill & Smith 2009; Klein 2009; Bae 2010). The locations of these hominin fossil localities can be found in Figure 1 and representative samples of the hominin fossils are presented in Figures 2 and 3.
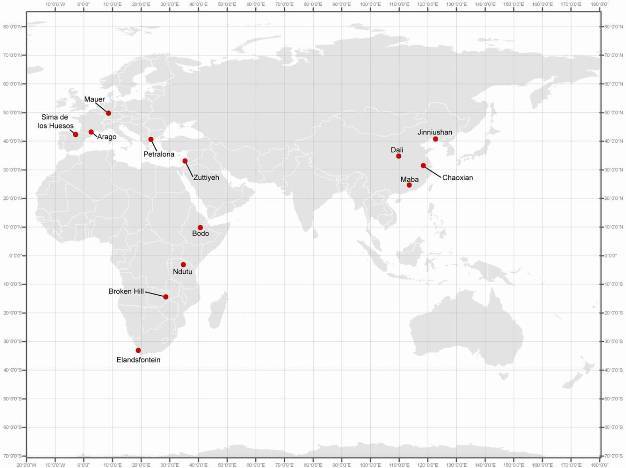
Figure 1
Locations of important archaic Homo sapiens/H. heidelbergensis localities across the Old World.
© 2013 Nature Education All rights reserved. 
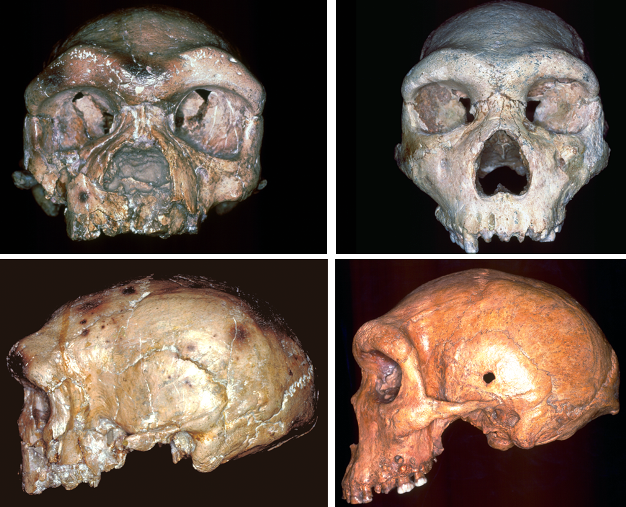
Figure 2
Frontal (top) and lateral (bottom) views of typical Homo heidelbergensis crania from Europe and Africa (left: Petralona; right: Kabwe).
© 2013 Nature Education Images reproduced with the kind permission of Milford Wolpoff. All rights reserved.
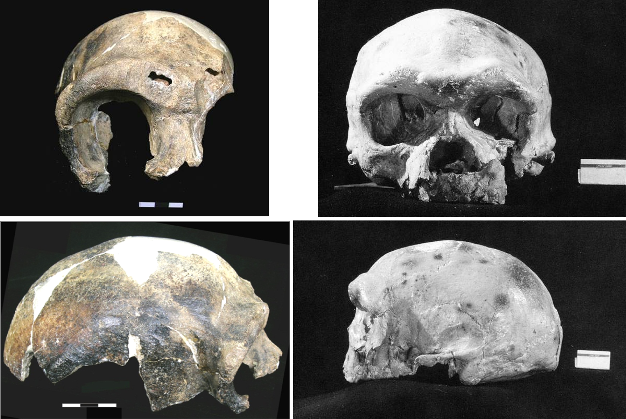
Figure 3
Frontal (top) and lateral (bottom) views of typical archaic Homo sapiens crania from China (left: Maba; right: Dali).
© 2013 Nature Education Maba images reproduced with the kind permission of Wu Liu and Dali images reproduced with the kind permission of Xinzhi Wu. All rights reserved.
In general, archaic Homo sapiens were ‘‘admitted to membership in our species because of their almost modern-sized brains, but set off as ‘archaic' because of their primitive looking cranial morphology" (Cartmill & Smith 2009). The primary morphological characteristics of archaic H. sapiens (Rightmire 2004, 2008) are: 1) average cranial capacity (~1,200 cc) and a proportional increase in encephalization that places them between modern H. sapiens (~1,350 cc) and H. erectus s.l. (~1,000 cc); 2) a reduced postorbital constriction, to account for the increase in cranial capacity; 3) the degree of overall cranial robustness somewhere between H. erectus s.l. and H. sapiens; 4) and compared to H. erectus, a more rounded and less angled occipital region. Relatively few studies have been conducted on postcranial remains. One of the more recent studies was an analysis of the postcranial remains from Jinniushan, China (Rosenberg et al. 2006). The Rosenberg et al. (2006) study reconstructed the Jinniushan hominin as a stocky female, lending support for the Allen and Bergmann's Rules for ecogeographic adaptation to cold environments.
Over the past several decades, there has been a push by a number of paleoanthropologists (e.g., Tattersall 1986; Groves & Lahr 1994; Rightmire 1998, 2004, 2008; Stringer 2002; Tattersall & Schwartz 2008) to refer to the Middle Pleistocene archaic Homo sapiens as H. heidelbergensis, following the original designation of the Mauer mandible which is the holotype of the taxon (Schoetensack 1908). The Mauer mandible is best known for being very robust, has a pronounced anteroposterior mandibular ramus length, and lacks a mental eminence (i.e., chin) (Mounier et al. 2009). Although the holotype of H. heidelbergensis is a mandible, similarities between Mauer and other penecontemporaneous hominin mandibles (e.g., Arago) appear to support the original designation as a new species. In turn, associated crania (e.g., Arago, Petralona) are then used to represent the H. heidelbergensis hypodigm. Overall similarities in general cranial bauplan between many of the European (e.g., Arago, Petralona) and African (e.g., Broken Hill) fossils have supported the lumping of these fossils into H. heidelbergensis (e.g., Rightmire 2008; Tattersall and Schwartz 2008).
Nonetheless, in a characteristic insightful observation made more than one half century ago, Mayr (1950) noted that "[s]ome anthropologists, in fact, imply that they use specific and generic names merely as labels for specimens without giving them any biological meaning." Thus, it might be argued that at least in certain cases, paleoanthropologists may be using specific and generic names, without any strong evolutionary biological underpinning. In other words, the different specific and generic names are used simply to distinguish different groups or regional populations. In turn, this has led some scientists to suggest that archaic H. sapiens should still be considered a viable option, particularly because not all Middle Pleistocene hominin fossils from Europe, Africa, and particularly eastern Asia can be easily allocated to the H. heidelbergensis hypodigm (Pope 1992; Wu & Brauer 1993; Rightmire 2004, 2008; Brauer 2008; Cartmill & Smith 2009; Bae 2010). For instance, Rightmire (2004) observed that the Chinese Dali clivus is likely shallow while the African Broken Hill "clivus is exceptionally deep and forward-sloping." It should be noted that there is a great deal of morphological variation within and between the European and African H. heidelbergensis fossils as well (Wu & Brauer 1993; Rightmire 2004). As some paleoanthropologists have noted recently (e.g., Brauer 2008; Cartmill & Smith 2009; Bae 2010), replacing one vague name (archaic H. sapiens) with another vague name (H. heidelbergensis) contributes little to deepening our understanding of Middle Pleistocene hominin phylogenetic relationships, which is required for our understanding of modern human origins.
Archaic Homo sapiens (paleoecology and behavior)
Besides Early Pleistocene occupations in higher latitudes by Homo erectus in Georgia (and possibly the Nihewan Basin, northern China), for the most part Middle Pleistocene archaic Homo sapiens were the most wide and northerly ranging hominin group prior to the arrival of modern H. sapiens on the scene (Cartmill & Smith 2009; Klein 2009). They occupied most regions of the Old World from southern Africa (e.g., Saldanha, South Africa) to western Europe (e.g., Mauer, Germany) to northeastern Asia (Jinniushan, China). Archaic H. sapiens occupied cold environments (e.g., Lehringen, Jinniushan) in the higher latitudes. In all likelihood, however, occupation of these higher latitudes was probably restricted to interglacial stages. Current data indicate that archaic H. sapiens were not able to move into higher altitudes until very late in human prehistory (~40,000-30,000 years BP), when barriers from very harsh environments were effectively overcome. This was a period when only modern H. sapiens with fully modern behavior roamed the landscape (Norton and Jin 2009; but see recent discussion on Homo floresiensis).
Evidence of adaptation to a colder climate include Middle Pleistocene open-air sites that display evidence of structures (e.g., Bilzingsleben) and more evidence of cave occupation (e.g., Le Lazaret, Jinniushan). The latter postdate such famous H. erectus cave sites as Zhoukoudian Locality 1 (Klein 2009). In order to colonize Eurasia, early hominins probably had to have controlled fire. However, evidence for fire only appears sporadically in the Early Pleistocene record, with the earliest generally accepted evidence appearing at the Middle Pleistocene site of Gesher Benot Ya'aqov in Israel (Goren Inbar et al. 2004; but see the recent evidence from the Early Pleistocene Wonderwerk cave in South Africa as discussed inBerna et al. 2012). The paucity of evidence of fire at sites older than the Late Pleistocene is probably related to the likelihood that occupations by H. erectus and archaic H. sapiens were short term compared to Neanderthals and modern H. sapiens.
The stone toolkits of archaic Homo sapiens did not differ much from earlier H. erectus. In the western Old World (Africa, Middle East, Europe) and South Asia, archaic H. sapiens relied on Lower Paleolithic core and flake stone tools with bifacially worked lithics commonly known as handaxes. In eastern Asia, the stone tools most commonly associated with archaic H. sapiens are the Oldowan core and flake tools of the Lower Paleolithic. It was originally noted that handaxes were absent in eastern Asia (Movius 1944). This pattern in the archaeological data came to be known as the Movius Line. The line was drawn between South Asia and eastern Asia with handaxes to the west of the line and traditional Oldowan core and flake tools to the east of the line (Norton and Lycett 2009). Although handaxes have been found in eastern Asia (Norton et al. 2006; Norton and Bae 2009; Lycett and Bae 2010; Lycett and Norton 2010), the general archaeological patterning still indicates a relative paucity of handaxes east of the Movius Line.
Early researchers (e.g., Binford 1984, 1985) suggested archaic Homo sapiens (and Neanderthals) were not efficient hunters. However, based on current archaeological research there is growing evidence to the contrary. For instance, three wooden spears were found at Schoningen, an open-air site from Germany that dates to about 400,000 years ago (Thieme 1997). The Schoningen spears are each two meters in length and are thought to have been used in a thrusting motion in close contact with animals rather than as throwing spears (Klein 2009). Taphonomic analysis of the plethora of horse bones from the archaic H. sapiens open-air site Xujiayao in northern China indicates these hominins were effectively procuring and processing the horses (Norton and Gao 2008; see also similar evidence from La Cotte de St. Brelade off the coast of England referenced byScott 1980). This growing efficiency in the regular procurement of animals is thought to have influenced the trend in the increase in encephalization quotient evident in the hominin fossil record from H. erectus to archaic H. sapiens to modern H. sapiens (Rightmire 2004; Bae 2010). However, there may have been periods of dietary stress where food sources not normally utilized were accessed by archaic H. sapiens. For instance, the Bodo (Ethiopia) and Arago (France) hominin fossils display evidence of cut marks, suggestive of cannibalistic behavior (de Lumley 1975; White 1985). It is difficult to determine whether these cut marks are the result of food processing or for cultural reasons, perhaps early symbolism.
Concluding Remarks
Different schools of thought lead to different ways of viewing the hominin fossil record. There appears to be growing support in the field for lumping the African and European archaic Homo sapiens fossils into H. heidelbergensis (Rightmire 1996, 1998, 2004; 2008; Stringer 2002; but see Brauer 2008). However, arguments have been made to keep the Asian, particularly eastern Asian, hominin fossils in a separate group (e.g., pre-modern H. sapiens, archaic H. sapiens) (Pope 1992; Bae 2010). Behaviorally, growing evidence suggests that archaic H. sapiens were successful at procuring animals. The widespread presence of Middle Pleistocene handaxes in the western Old World and the rarity of similar sites in eastern Asia still need to be explained, although there appears to be increasing support for a few hypotheses. Questions over the validity and meaning of archaic H. sapiens (and H. heidelbergensis), both taxonomically and behaviorally, will surely continue to be intensively debated in the coming years (e.g., Binford, 1985; Tattersall 1986; Groves & Lahr 1994; Rightmire 1998, 2008; Brauer 2008; Tattersall & Schwartz 2008; Bae 2010; Stringer 2012). Fortunately, with the ever-increasing Middle Pleistocene hominin fossil and associated archaeological and paleoecological datasets, debates like these will hopefully have some resolution as we move forward.
Acknowledgements
I thank Holly Dunsworth for the kind invitation to contribute this paper. Many thanks to Wu Liu, Milford Wolpoff, and Xinzhi Wu for permission to use the images of the hominin fossils and to Josephine Yucha for producing Figure 1. Generous funding to support this research was provided by the Academy of Korean Studies - Korean Studies Promotion Service (AKS-2010-DZZ-3102). I appreciate the thoughtful comments from Holly Dunsworth and two anonymous reviewers. I take full responsibility for any errors that may be present.
References and Recommended Reading
Bae, C. J. The late Middle Pleistocene hominin fossil record of Eastern Asia: synthesis and review. Yearbook of Physical Anthropology 53, 75-93 (2010).
Berna, F. et al. Microstratigraphic evidence of in situ fire in the Acheulean strata of Wonderwerk Cave, Northern Cape province, South Africa. Proceedings of the National Academy of Sciences 109, E1215-E1220 (2012).
Binford, L. R. Faunal Remains from Klasies River Mouth. New York: Academic Press (1984).
Binford, L. R. Human ancestors: changing views of their behavior. Journal of Anthropological Archaeology 4, 292-327 (1985).
Brauer, G. The origin of modern anatomy: by speciation or intraspecific evolution? Evolutionary Anthropology 17, 22-37 (2008).
Cartmill, M. & Smith, F. H. The Human Lineage. Hoboken NJ: Wiley-Blackwell (2009).
Conroy, G. C. Reconstructing Human Origins, 2nd edition. New York: W.W. Norton (2005).
de Lumley, H. Cultural evolution in France in its paleoecological setting during the Middle Pleistocene. In: After the Australopithecines, ed. K. W. Butzer & G. L. Isaac, pp. 745-808. The Hague: Mouton (1975).
Etler, D. A. The fossil evidence for human evolution in Asia. Annual Review of Anthropology 25, 275-301 (1996).
Goren-Inbar, N. et al. Evidence of hominin control of fire at Gesher Benot Ya'aqov, Israel. Science 304, 725-727 (2004).
Groves, C. P. & Lahr, M. M. A bush not a ladder: speciation and replacement in human evolution. Perspectives in Human Biology 4, 1-11 (1994).
Klein, R. G. The Human Career: Human Biological and Cultural Origins, 3rd edition. Chicago, IL: University of Chicago Press (2009).
Lycett, S. J. & Bae, C. J. The Movius Line and Old World Palaeolithic patterns: the state of the debate. World Archaeology 42, 521-544 (2010).
Lycett, S. J. & Norton, C. J. A demographic model for Palaeolithic technological evolution: the case of East Asia and the Movius Line. Quaternary International 211, 55-65 (2010).
Mayr, E. Taxonomic categories in fossil hominids. Cold Spring Harbor Symposia on Quantitative Biology 15, 109-118 (1950).
Mounier, A., Marchal, F. & Condemi, S. Is Homo heidelbergensis a distinct species? New insight on the Mauer mandible. Journal of Human Evolution 56, 219-246 (2009).
Movius, H. L. Early Man and Pleistocene Stratigraphy in Southern and Eastern Asia. Harvard University, Peabody Museum papers 19(3). Cambridge: Peabody Museum (1944).
Norton, C. J. & Gao, X. Hominin-carnivore interactions during the Chinese Early Paleolithic: Taphonomic perspectives from Xujiayao. Journal of Human Evolution 55, 164-178 (2008).
Norton, C. J. & Jin, J. The evolution of modern humans in East Asia: behavioral perspectives. Evolutionary Anthropology 18, 247-260 (2009).
Norton, C. J. & Lycett, S. J. The Movius Line. McGraw-Hill Yearbook of Science and Technology 2010, 248-250 (2009).
Norton, C. J., Bae, K. D. Erratum to "The Movius Line sensu lato (Norton et al., 2006) further assessed and defined" J. H. Evol. 55 (2008) 1148-1150. Journal of Human Evolution 57, 331-334 (2009).
Norton, C.J., Bae, K. D., Harris, J. W. K., Lee, H. Y. Middle Pleistocene handaxes from the Korean Peninsula. Journal of Human Evolution 51, 527-536 (2006).
Pope, G. G. Craniofacial evidence for the origin of modern humans in China. Yearbook of Physical Anthropology 35, 243-298 (1992).
Rightmire, G. P. The human cranium from Bodo, Ethiopia: evidence for speciation in the Middle Pleistocene? Journal of Human Evolution 31, 21-39 (1996).
Rightmire, G. P. Human evolution in the Middle Pleistocene: the role of Homo heidelbergensis. Evolutionary Anthropology 6, 218-227 (1998).
Rightmire, G. P. Brain size and encephalization in early to mid-Pleistocene Homo. American Journal of Physical Anthropology 124, 109-123 (2004).
Rightmire, G. P. Homo in the Middle Pleistocene: hypodigms, variation, and species recognition. Evolutionary Anthropology 17, 8-21 (2008).
Rosenberg, K. R. et al. Body size, body proportions, and encephalization in a Middle Pleistocene archaic human from northern China. Proceedings of the National Academy of Sciences USA 103, 3552-3556 (2006).
Schoetensack, O. Der Unterkiefer des Homo heidelbergensis aus den Sanden von Mauer bei Heidelberg. Leipzig: Ein Beitrag zur Paleontologie des Menschen (1908).
Scott, K. Two hunting episodes of middle Palaeolithic age at La Cotte de Saint‐Brelade, Jersey (Channel Islands). World Archaeology 12, 137-152 (1980).
Stringer, C. Modern human origins: progress and prospects. Philosophical Transactions of the Royal Society of London B 357, 563-579 (2002).
Stringer, C. The status of Homo heidelbergensis (Schoetensack 1908). Evolutionary Anthropology 21, 101-107 (2012).
Tattersall, I. Species recognition in human paleontology. Journal of Human Evolution 15, 165-175 (1986).
Tattersall, I. & Schwartz, J. H. The morphological distinctiveness of Homo sapiens and its recognition in the fossil record: clarifying the problem. Evolutionary Anthropology 17, 49-54 (2008).
Thieme, H. Lower Paleolithic hunting spears from Germany. Nature 385, 807-810 (1997).
White, T. D. Acheulian man in Ethiopia's Middle Awash Valley: The implications of cut marks on the Bodo cranium. Amsterdam: Nederlands Museum voor Anthropologie en Praehistorie (1985).
Wolpoff, M. et al. The case for sinking Homo erectus: 100 years of Pithecanthropus is enough! Courier Forschung-Institut Senckenberg 171, 341-361 (1994).
Wu, X. & Brauer, G. Morphological comparison of archaic Homo sapiens crania from China and Africa. Zeitschrift fur Morphologie und Anthropologie 79, 241-259 (1993).
Wu, X. Z. & Poirier, F. E. Human Evolution in China. Oxford: Oxford University Press (1995).










