





















« Prev Next »

Neanderthals are a hominin species that existed for at least 200,000 years throughout Europe and western Asia, and disappeared about 27,000 years ago (ya). During this time, they witnessed some of the coldest climatic conditions ever known in these regions. Many of their physical features suggest that they were adapted for the cold, such as their barrel-shaped chests, shorter limbs, and larger brains, all of which suggest a body shape adapted for retaining heat. They excelled at hunting animals and making complex stone tools, and their bones reveal that they were extremely muscular and strong, but led hard lives, suffering frequent injuries. There is no doubt that Neanderthals were an intelligent species, successfully adapted to their environment for over 200 millenia. But how "human" were they? Did they exhibit behaviors that we consider to be uniquely human, such as language, music, art, and religion? And why did they disappear from Europe so soon after modern humans moved in?
Lithic Technology
Middle Paleolithic core technology
The Neanderthals are associated with the archaeological period known as the Middle Paleolithic (MP), which most scholars place between 300,000–35,000 ya (Monnier 2006, Richter 2011). The two main defining characteristics of the stone tools from the Middle Paleolithic are a decrease in the frequency of large bifacial cutting tools (handaxes and cleavers) and the appearance of prepared-core technology, such as Levallois (Figure 1). Microwear and residue studies have suggested that MP tools were used for a wide variety of tasks, including butchery, plant processing, and woodworking (Beyries 1988, Anderson-Gerfaud 1990, Hardy 2004, Hardy et al. 2001, Plisson & Beyries 1998). As the methodologies for these types of analyses improves, we will gain a better understanding of MP tool use. 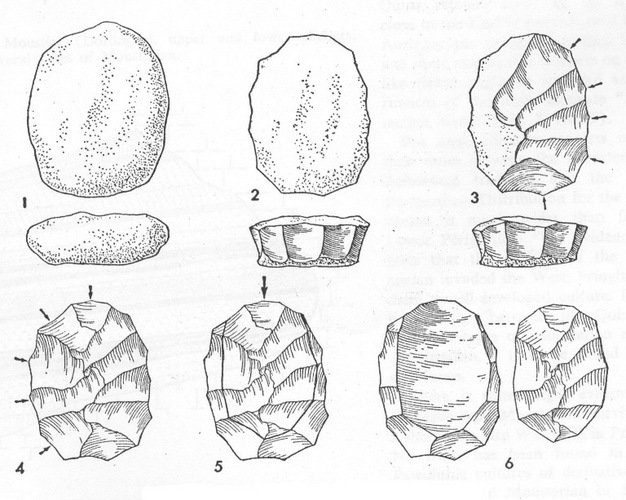
Figure 1: Levallois technology scheme.
Levallois technology scheme (From Bordes 1961b). The stages of manufacture of a Levallois core: 1, selection of an unretouched cobble; 2, removal of flakes around the periphery of the core; 3, removal of flakes from the main surface of the core, using the previous removals as platforms; 4, continuation of flaking in order to create a convex surface; 5, removal of the central flake; 6, the Levallois core and flake.
© 2012 Nature Education All rights reserved. 
Neanderthals' frequent use and mastery of Levallois technology demonstrates that they were expert knappers who had the cognitive abilities necessary to design and execute complex knapping sequences. In addition to Levallois technology (Boëda 1994, 1995), other MP flaking technologies have been identified, such as blade, discoidal, Quina, and bifacial technologies (see Delagnes & Meignen 2006, and references therein).
Retouched flake tools and the great Mousterian debate
Neanderthals used many of the flakes — termed "débitage" in French — that they removed from cores as tools. Some flakes were used without further modification, while others were shaped into various types of retouched tools. Many of the Middle Paleolithic assemblages of cores, flakes, and retouched tools are referred to as Mousterian, the name given to this type of archaeological culture in the late 19th century, from the type site of Le Moustier (France). François Bordes' typology (Bordes 1961a) of Middle Paleolithic flake tools divides them into scrapers (Figure 2), points, denticulates, backed knives, bifaces, and miscellaneous other types. On the basis of the relative frequencies of tool types such as scrapers versus denticulates, as well as the frequency of Levallois flakes and bifaces, Bordes devised a classification of assemblages into different Mousterian facies (Bordes 1953a, 1961b). According to Bordes, these facies were the product of different culture groups (Bordes & de Sonneville-Bordes 1970). Others disagreed, suggesting that the facies represented diachronic patterning (Mellars 1965, 1969), different activities (Binford & Binford 1966), scraper reduction (Dibble 1984, 1995), or intensity of raw material use and climate (Rolland & Dibble 1990). This controversy has been called "The Great Mousterian Debate." Each of these interpretations has furthered our understanding of the meaning of the variability in retouched tool types in the Middle Paleolithic. Unfortunately, a more complete understanding of what these different tool types represent is hampered by a relative lack of information regarding what these tools were used for.
While identifying specifics about tool use has proven difficult, we have begun to learn how Neanderthals used their landscapes by studying patterns of faunal exploitation and raw material transport. Evidence shows that, in western Europe, most raw materials used for stone tools are local and come from sources within 5 km of each site. This suggests that Neanderthals circulated within relatively small territories. In central Europe, however, raw materials traveled further, indicating more wide-ranging mobility patterns (Feblot-Augustins 1993) or a more limited availability of high-quality raw material sources (Mellars 1996). Studies of faunal exploitation and the curation of raw materials from sites throughout Europe have shown that Neanderthals were able to plan subsistence strategies and adapt their tool-making behavior to these subsistence strategies, revealing behavioral flexibility and problem-solving abilities (Kuhn 1995, Burke 2006).
Neanderthals and the Upper Paleolithic
Neanderthals are also associated with an early Upper Paleolithic industry, the Châtelperronian (~35,000 ya), at two sites in France: Arcy-sur-Cure (Leroi-Gourhan 1959, Hublin et al. 1996, Bailey & Hublin 2006) and Saint-Césaire (Léveque et al. 1993). The Châtelperronian is similar to the Aurignacian, and since the rest of the Upper Paleolithic — from the Aurignacian on — is associated with modern humans, the association of Neanderthals with the Châtelperronian has generated a powerful controversy. Some have suggested that Neanderthals were "acculturated" by modern humans bearing the Aurignacian type of Upper Paleolithic, and learned from them to make the blade tools and beads characteristic of the Châtelperronian (Mellars 1996, 2005). Others, however, have countered that the innovations seen in the Châtelperronian occurred before modern humans had arrived, demonstrating that Neanderthals had the ability to make the sophisticated material culture of the Upper Paleolithic (d'Errico 2003, Zilhao 2006). Recently, however, the association of Neanderthal remains-mostly teeth-with Châtelperronian layers at Arcy-sur-Cure has been called into question on the basis of evidence indicating possible mixing of archaeological sediments (Higham et al. 2011, Bar-Yosef & Bordes 2010). The implication is that the Châtelperronian, like the rest of the Upper Paleolithic, may be a product of modern humans, and that the presence of some Neanderthal remains in these layers does not mean that the Neanderthals made the Châtelperronian artifacts.
Hunting technology
Great strides have been made in understanding Neanderthal hunting technology. Neanderthals were consummate hunters of medium and large-sized mammals. There is evidence that they used stone-tipped spears to hunt. For instance, it has been observed that Levallois points often bear impact scars on their tips (Shea 1988). In addition, retouched Mousterian points from several sites in western Europe (La Cotte de St. Brelade, Bouheben, Oscurusciuto, and Abric del Pastor) have been identified as bearing impact scars too (Villa & Soriano 2010). These stone-tipped spears were probably used at close-range for thrusting, or thrown short distances as part of an ambush hunting strategy — neither the stone points nor the features of Neanderthal anatomy support the notion that Neanderthals used long-distance projectiles (Shea & Sisk 2010, Churchill & Schmitt 2002, Rhodes & Churchill 2008).
Subsistence
There are several lines of evidence indicating that Neanderthals were capable hunters whose diets were dominated by meat. First is the faunal evidence from various sites, which indicates that they hunted and butchered red deer (Cervus elaphus), horse (Equus sp.), bison (Bison priscus), wooly rhinoceros (Coelodonta antiquitatis), and many other species of large and medium-sized ungulates (Patou-Mathis 2000). They focused on prime-aged adult prey, as opposed to juveniles and older adults, which are typically the target of cursorial predators (Stiner 2006). This behavior is likely to have begun 250,000 years ago or earlier, as seen at Hayonim Cave, Israel (Stiner 2005). There is also evidence that Neanderthals occasionally specialized in a single prey species, such as at Mauran, France, which is estimated to contain the remains of ~4,000 bison (Farizy et al. 1994). Similarly, the fauna at La Borde, France (Jaubert et al. 1990) and Wallertheim, Germany (Gaudzinski 1995) is also dominated by bovids, and these have been interpreted as kill sites. The site of Salzgitter-Lebenstedt (Germany) seems to have been a specialized reindeer-hunting site, where reindeer were taken in the early autumn during their annual migrations. The cut-marks clearly show meat processing, and the marrow-rich bones were also processed (Gaudzinski & Roebroeks 2000, Gaudzinski 2006). The evidence from all of these sites indicates that Neanderthals were able to hunt large game and often specialized in a species at a particular location and during a particular season.
Nitrogen and carbon isotope studies also support a Neanderthal diet focused on meat. Collagen obtained directly from at least a dozen Neanderthal bones from sites in France, Belgium, and Croatia reveal that Neanderthals were top-level carnivores who obtained most of their dietary protein from large herbivores (Richards & Trinkaus 2009, Bocherens 2009). Nevertheless, there is evidence that Neanderthals incorporated smaller animals and plant foods into their diets too. For instance, Neanderthals collected small game such as tortoises, shellfish, hares, rabbits, and birds in the Mediterranean region, and this trend increased dramatically just prior to or within the MP/UP transition (Stiner 2006). At the site of Vanguard Cave, Gibraltar, a small assemblage of mollusks was found, in addition to a few marine mammal bones (monk seal and dolphin) with cut-marks, indicating that Neanderthals exploited marine resources when they had the opportunity (Stringer et al. 2008).
There is also evidence that Neanderthals occasionally practiced cannibalism — at the site of Moula-Guercy, France, a number of Neanderthal remains were found with butchery marks indicating defleshing and marrow extraction. The similarity between the butchery marks on the Neanderthal bones and those on deer suggests the Neanderthals were consuming their dead for food (Defleur et al. 1999). Finally, there is evidence that Neanderthals consumed plant resources. Starch grains from legumes, date palms, and other plants — some of them exhibiting the transformations that happen during cooking — have been found preserved in dental calculus of Neanderthals from Shanidar, Iraq, and Spy, Belgium (Henry et al. 2011). In addition, a large assemblage of charred seeds and fruits (legumes, acorns, pistachios) were recovered from the Mousterian levels at Kebara Cave, Israel (Lev et al. 2005), and phytoliths recovered from sediments at Amud Cave suggest the consumption of grasses (Madella et al. 2002).
Symbolism
One of the most pressing questions about Neanderthals is whether they had symbolic thought. Symbolic thought is widely agreed to be one of the hallmarks of modern (McBrearty & Brooks 2000) or complex (Langley et al. 2008) human behavior. It is also associated with the cognitive underpinnings of language (Chase 1991, Deacon 1997). Archaeologically, symbolism may be manifested as figurative art, ornamentation, use of pigments, and ritual burial (d'Errico et al. 2003, McBrearty & Brooks 2000, Mellars 2005, Langley et al. 2008). While there is no figurative art associated with Neanderthals, personal ornaments such as pierced or grooved animal teeth and ivory rings have been found at the Châtelperronian sites of Saint-Césaire and Arcy-sur-Cure, France (Lévêque et al. 1993, Hublin et al. 1996). However, as mentioned above, the question remains of whether Neanderthals were the makers of the Châtelperronian (Bar-Yosef & Bordes 2010). If they were not, then there is no other evidence indicating that they made ornaments. Mineral pigments are often used in symbolic contexts among humans today, but their chemical properties make them useful in activities such as hide tanning and for tool-hafting (Hovers et al. 2003). There is evidence that Neanderthals used mineral pigments such as red ochre (hematite) and manganese dioxide at several sites in Europe, including Pech de l'Azé I, France (d'Errico 2003), and Cueva de los Aviones and Cueva Antón, Spain (Zilhao et al. 2009). However, we do not know whether this pigment use was symbolic or not. Neanderthals most likely buried their dead, as evidenced from the presence of articulated Neanderthal skeletons found in dozens of sites (Langley et al. 2008, Riel-Salvatore & Clark 2001) (Figure 3). While some have argued that these skeletons were preserved by natural causes, such as cave roof collapses (Gargett 1989, 1999), rather than deliberately buried, there is general consensus that the completeness of the skeletons validates an interpretation of deliberate burial (Belfer-Cohen & Hovers 1992, d'Errico 2003). The lack of clear grave goods associated with these burials, however, lends doubt to the issue of whether these are symbolic burials, although they do indicate caring for the deceased beyond what is seen in non-human primates (Chase & Dibble 1987). The suggestion that Neanderthals at Shanidar Cave, Iraq, were buried with flowers has never been substantiated. While flower pollen was found in sediments near the burials, it could have been introduced by rodents such as the Persion Jird, which is known to gather flowers and bring them back to its burrow (Sommer 1999). In sum, there is evidence to suggest that Neanderthals had some level of symbolic thought. The use of mineral pigments, the presence of burials, and other indicators of behavioral complexity, such as composite technology, become more frequent after 60,000 ya (Langley et al. 2008), suggesting that behavioral complexity and symbolic thought may have emerged as a component of Neanderthal adaptations towards the end of their existence as a species.

Figure 3: Photograph of the cast of the Neanderthal burial found at Kebara Cave, Israel, in 1983, and dated to 60,000 years before present.
© 2012 Nature Education Courtesy of Ofer Bar-Yosef. All rights reserved.
The Fate of the Neanderthals
The transition from the Middle to the Upper Paleolithic in Europe also marks the transition from a time of Neanderthal occupation to that of modern humans. It is widely accepted that modern humans first arrived in Europe from the southeast, bearing the Aurignacian culture, and replaced the Neanderthals. The latest MP assemblages have been documented in the Iberian peninsula at sites such as Gorham's Cave, Gibraltar at 28,000 ya (Finlayson 2006; but see Zilhao and Pettitt 2006 for a critique of these dates), and it has been suggested that the Neanderthals were "pushed" to this cul-de-sac by the invading modern humans, deteriorating climatic conditions, or both. Some have argued that the colder and highly variable climate of oxygen isotope stage 3 (57–24,000 ya; van Andel & Davies 2003, Finlayson & Carrion 2007) resulted in periods of climatic stress which may ultimately have been responsible for the demise of the Neanderthals (Stringer et al. 2003) through mechanisms such as the fragmentation of Neanderthal-favored habitats and their inability to adapt to new habitats (Finlayson 2004). Others have suggested that the Campanian Ignimbrite volcanic eruption at 40,000 ya initiated a volcanic winter that had a devastating effect on the ecology of eastern and southeastern Europe (Fedele et al. 2008), possibly wiping out populations of Neanderthals from Central Europe to the Caucasus (Golovanova et al. 2010). Demographic modeling (Sorenson 2010) and eco-cultural niche modeling (Banks et al. 2008) have both rejected climate as a driving factor for Neanderthal extinction, and suggested that other factors may have been involved, such as disease, violence, or competition with modern humans. Finally, some have suggested that Neanderthals did not go extinct, but were assimilated into populations of modern humans (Smith et al. 2005). Recent genetic studies have shown that modern European and Asian DNA contains 1–4% Neanderthal genes. This suggests that before Neanderthals became extinct, some, at least, interbred with modern humans (Green et al. 2010).
Glossary
Aurignacian: An early Upper Paleolithic archaeological culture found throughout Europe and western Eurasia, which dates to 30–35,000 years ago. It is characterized by retouched tools made on large blades, and polished split-base bone points. A variety of other worked bone objects and ornaments made of bone or teeth are also present.
Campanian Ignimbrite: A massive volcanic eruption from southern Italy that occurred about 40,000 years ago. Ash from this eruption has been found in deposits throughout the world. The large amounts of ash that resulted in the sky may have severely affected world climate.
Châtelperronian: An early Upper Paleolithic archaeological culture which has been identified at sites in southern France and northern Spain, and which dates to 33–36,000 years ago. It contains both blade and flake technology and a diagnostic retouched stone point, the "Chatelperronian point." It is also characterized by worked bone, including polished bone points, and ornaments such as bone pendants and pierced or grooved teeth.
Cleavers: Large, bifacially flaked core-tools, one end of which is formed by the intersection of a large flake scar on each face. Instead of being pointed, cleavers have a blunt, but sharp tip. They are most common in Africa but do occur in some Lower Paleolithic sites in Europe.
Composite technology: Refers to tools and weapons that consist of several parts, such as stone points hafted to a wooden shaft.
Cursorial: An animal adapted to run.
Diachronic: Refers to changes through time.
Handaxes: Large, pointed, bifacially flaked core-tools. Handaxes and cleavers first appear in the archaeological record in Africa about 1.6 million years ago. By 500,000 years ago, handaxes appear in Europe. Their presence is often used to assign a site to the Lower Paleolithic.
Isotope analysis: The measurement of Carbon (13C, 12C) and Nitrogen (14N, 15N) isotopes in human or animal bone mineral in order to reconstruct the diet of the individual.
Levallois technology: A method for knapping stone that is characteristic of the Middle Paleolithic. A nodule of stone is flaked to produce a core with two faces, one of which is highly convex. The convex face serves as a platform for removing flakes from the other face. These flakes tend to be large and flat, and have a distinctive appearance that qualifies them as "Levallois flakes". Several variants of Levallois technology have been identified by Eric Boëda of the University of Paris X, including one named "preferential" in which the main surface of the core is designed to produce just one large flake before being reshaped for the next removal, and "recurrent", in which a series of Levallois flakes are removed before the core is reshaped. Because Levallois technology entails preparing a core for the removal of just a few flakes of a desired shape and size, it is widely referred to as a prepared core technology.
Microwear analysis: A method for reconstructing the function of stone tools by microscopically studying the wear patterns on their edges, and comparing these to experimentally produced tools.
Middle Paleolithic: An archaeological phase widely recognized throughout Europe and western Asia. It is commonly defined as beginning with the first appearance of prepared-core technology, about 300,000 or 250,000 years ago, and ending with the appearance of the Upper Paleolithic about 25,000 years ago.
Mousterian: An archaeological culture which has been identified at sites throughout Europe and western Asia, and is sometimes used as a synonym for "Middle Paleolithic", with which it is broadly contemporaneous. It is characterized by a variety of retouched stone tools including side-scrapers, points, and denticulates. These tools are usually made on flakes knapped using prepared-core technologies.
Mousterian facies: A set of variants of the Mousterian codified by François Bordes in the 1950s. It includes the Ferrassie Mousterian, characterized by Levallois technology and high frequencies of scrapers; Quina Mousterian, in which Levallois technology is rare but scraper frequencies are high; Denticulate Mousterian, which contains few scrapers but many notched and denticulated tools; and Mousterian of Acheulean Tradition, which contains scrapers as well as small bifaces.
Mousterian points: Retouched flake tools characteristic of the Middle Paleolithic. The retouched edges converge to form a point, which may or may not have been hafted and used as a hunting weapon.
Oxygen isotope stages: Divisions of major cold and warm phases in the Earth's climate, as reconstructed from oxygen isotope cores from the Greenland ice sheets.
Phytoliths: Microscopic silica bodies formed in the cells, and in the interstices between cells, of certain plants. They are often well-preserved in archaeological sites, and their distinctive shapes can sometimes enable a specialist to identify them.
Residue analysis: A method for reconstructing the function of stone tools by microscopically studying residues adhering to the surface of stone tools.
Scraper reduction: A model of scraper manufacture and use developed by Harold Dibble from the University of Pennsylvania. It posits that scraper form reflects intensity of utilization of tools, because increased use leads to repeated resharpening of scrapers, changing their forms.
Symbolism: The act of depicting a concept, object, person, deity, etc. with a visual or auditory representation that does not resemble such concept or object. Symbols do not look like their referents. The meaning of a symbol is shared by a group of people, or a culture. Language is symbolic in nature, because most words do not resemble their referents.
Upper Paleolithic: An archaeological phase widely recognized throughout Europe and western Asia. It begins with the appearance of prismatic blade technology and distinctive Upper Paleolithic stone tools around 35,000 years ago, and in many places it is replaced by the following phase — the Mesolithic — around 10,000 years ago.
References and Recommended Reading
Anderson, P. C. A testimony of prehistoric tasks: Diagnostic residues on stone tool working edges. World Archaeology 12, 181-193 (1980).
Anderson-Gerfaud, P. "Aspects of behaviour in the Middle Paleolithic: Functional analysis of stone tools from south-west France," in The Emergence of Modern Humans, ed. P. A. Mellars (Ithaca, NY: Cornell University Press, 1990) 389-418.
Bailey, S. E. & Hublin, J. Dental remains from the Grotte du Renne at Arcy-sur-Cure (Yonne). Journal of Human Evolution 50, 485-508 (2006). doi:10.1016/j.jhevol.2005.11.008
Banks, W. E. et al. Neanderthal extinction by competitive exclusion. PLoS One 3, e3972 (2008). doi:10.1371/journal.pone.0003972
Bar-Yosef, O. & Bordes, J. Who were the makers of the Châtelperronian culture? Journal of Human Evolution 59, 586-593 (2010). doi:10.1016/j.jhevol.2010.06.009
Bar-Yosef et al. The excavations in Kebara Cave, Mt. Carmel. Current Anthropology 33, 497-550 (1992).
Belfer-Cohen, A. & Hovers, E. In the eye of the beholder - Mousterian and Natufian burials in the Levant. Current Anthropology 33, 463-471 (1992).
Beyries, S. Industries Lithiques: Tracéologie et Technologie. London, UK: British Archaeological Reports, 1988.
Binford, L. R. "Interassemblage variability - The Mousterian and the 'functional' argument," in Explanation of Culture Change: Models in Prehistory (Pittsburgh, PA: University of Pittsburgh Press, 1973) 227-254.
Binford, L. R. & Binford, S. R. (1966). Preliminary analysis of functional variability in the Mousterian of Levallois facies. American Anthropologist 68, 238-295 (1966).
Bocherens, H. (2009). "Neanderthal dietary habits: Review of the isotopic evidence," in The Evolution of Hominin Diets: Integrating Approaches to the Study of Palaeolithic Subsistence, eds. J. J. Hublin, & M. P. Richards (New York, NY: Springer, 2009) 241-250.
Boëda, E. et al. A Levallois point embedded in the vertebra of a wild ass (Equus africanus): Hafting, projectiles and Mousterian hunting weapons. Antiquity 73, 394-402 (1999).
Boëda, E. Le Concept Levallois: Variabilité des Méthodes. Paris, France: CNRS, 1994.
Boëda, E. "Levallois: A volumetric construction, methods, a technique," in The Definition and Interpretation of Levallois Technology, eds. H. L.Dibble & O. Bar-Yosef (Madison, WI: Prehistory Press, 1995) 41-68.
Boeda, E. et al. Middle Palaeolithic bitumen use at Umm el Tlel around 70 000 BP. Antiquity 82, 853-861 (2008).
Bordes, F. H. Essai de classification des industries Moustériennes. Bulletin de la Société Préhistorique Française 50, 457-466 (1953).
Bordes, F. H. Typologie du Paléolithique Ancien et Moyen. Paris, France: CNRS, 1961a
Bordes, F. H. Mousterian cultures in France. Science 134, 803-810 (1961b).
Bordes, F. H. & de Sonneville-Bordes, D. The significance of variability in Palaeolithic assemblages. World Archaeology 2, 61-73 (1970).
Burke, A. Neanderthal settlement patterns in Crimea: A landscape approach. Journal of Anthropological Archaeology 25, 510-523 (2006). doi:10.1016/j.jaa.2006.03.003
Chase, P. G. Symbols and Paleolithic artifacts - Style, standardization, and the imposition of arbitrary form. Journal of Anthropological Archaeology 10, 193-214 (1991).
Chase, P. G. The Emergence of Culture: The Evolution of a Uniquely Human Way of Life. New York, NY: Springer, 2006.
Chase, P. G. & Dibble, H. L. Middle Paleolithic symbolism - A review of current evidence and interpretations. Journal of Anthropological Archaeology 6, 263-296 (1987).
Deacon, T. The Symbolic Species. London, UK: Penguin, 1997.
Defleur, A. et al. Neanderthal cannibalism at Moula-Guercy, Ardeche, France. Science 286, 128-131 (1999).
Delagnes, A. & Meignen, L. "Diversity of lithic production systems during the Middle Paleolithic in France: Are there any chronological trends?" in Transitions Before the Transition: Evolution and Stability in the Middle Paleolithic and Middle Stone Age, eds. E. Hovers & S. L. Kuhn (New York, NY: Springer, 2006) 85-108.
D'Errico, F. The invisible frontier. A multiple species model for the origin of behavioral modernity. Evolutionary Anthropology 12, 188-202 (2003). doi:10.1002/evan.10113
D'Errico, F. et al. Archaeological evidence for the emergence of language, symbolism, and music - An alternative multidisciplinary perspective. Journal of World Prehistory 17, 1-70 (2003).
Dibble, H. L. Interpreting typological variation of Middle Paleolithic scrapers: Function, style, or sequence of reduction? Journal of Field Archaeology 11, 431-436 (1984).
Dibble, H. L. The interpretation of Middle Paleolithic scraper morphology. American Antiquity 52, 109-117 (1987).
Dibble, H. L. Middle Paleolithic scraper reduction: Background, clarification, and review of the evidence to date. Journal of Archaeological Method and Theory 2, 299-368 (1995).
Farizy, C., David, F. & Jauber, J. Hommes et Bisons du Paléolithique Moyen à Mauran (Haute Garonne). Paris, France: CNRS, 1994.
Féblot-Augustins, J. Mobility strategies in the late Middle Paleolithic of Central Europe and Western Europe: Elements of stability and variability. Journal of Anthropological Archaeology 12, 211-265 (1993).
Fedele, F. G., Giaccio, B. & Hajdas, I. Timescales and cultural process at 40,000 BP in the light of the Campanian Ignimbrite eruption, Western Eurasia. Journal of Human Evolution 55, 834-857 (2008). doi:10.1016/j.jhevol.2008.08.012
Finlayson, C. Neanderthals and Modern Humans: An Ecological and Evolutionary Perspective. Cambridge, UK: Cambridge University Press, 2004.
Finlayson, C. & Carrion, J. S. Rapid ecological turnover and its impact on Neanderthal and other human populations. Trends in Ecology & Evolution 22, 213-222 (2007). doi:10.1016/j.tree.2007.02.001
Finlayson, C. et al. Late survival of Neanderthals at the southernmost extreme of Europe. Nature 443, 850-853 (2006). doi:10.1038/nature05195
Gargett, R. H. Grave shortcomings - The evidence for Neandertal burial. Current Anthropology 30, 157-190 (1989).
Gargett, R. H. Middle Palaeolithic burial is not a dead issue: The view from Gafzeh, Saint-Cesaire, Kebara, Amud, and Dederiyeh. Journal of Human Evolution 37, 27-90 (1999).
Gaudzinski, S. Wallertheim revisited: A re-analysis of the fauna from the Middle Paleolithic site of Wallertheim (Rheinhessen, Germany). Journal of Archaeological Science 22, 51-66 (1995).
Gaudzinski, S. Subsistence patterns of Early Pleistocene hominids in the Levant -Taphonomic evidence from the 'Ubeidiya Formation (Israel). Journal of Archaeological Science 31, 65-75 (2004). doi:10.1016/s0305-4403(03)00100-6
Gaudzinski, S. "Monospecific or species-dominated faunal assemblages during the Middle Paleolithic in Europe." in Transitions Before the Transition: Evolution and Stability in the Middle Paleolithic and Middle Stone Age, eds. E. Hovers & S. L. Kuhn (New York, NY: Springer, 2006) 137-147.
Gaudzinski, S. & Roebroeks, W. Adults only. Reindeer hunting at the Middle Paleolithic site Salzgitter Lebenstedt, Northern Germany. Journal of Human Evolution 38, 497-521 (2000).
Golovanova, L. V. et al. Significance of ecological factors in the Middle to Upper Paleolithic transition. Current Anthropology 51, 655-691 (2010). doi:10.1086/656185
Green, R. E., Krause, J. & Briggs, A. W. A draft sequence of the Neandertal genome. Science 328, 710-722 (2010).
Hardy, B. L. Neanderthal behaviour and stone tool function at the Middle Palaeolithic site of La Quina, France. Antiquity 78, 547-565 (2004).
Hardy, B. L. et al. Stone tool function at the Paleolithic sites of Starosele and Buran Kaya III, Crimea: Behavioral implications. Proceedings of the National Academy of Sciences of the United States of America 98, 10972-10977 (2001).
Henry, A. G., Brooks, A. S. & Piperno, D. R. Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium). Proceedings of the National Academy of Sciences of the United States of America 108, 486-491 (2011). doi:10.1073/pnas.1016868108
Higham, T. et al. Chronology of the Grotte du Renne (France) and implications for the context of ornaments and human remains within the Châtelperronian. Proceedings of the National Academy of Sciences of the United States of America 107, 20234-20239 (2010). doi:10.1073/pnas.1007963107
Hovers, E. et al. An early case of color symbolism - Ochre use by modern humans in Gafzeh Cave. Current Anthropology 44, 491-522 (2003).
Hublin, J. J. et al. A late Neanderthal associated with Upper Palaeolithic artefacts Nature 382, 93-93 (1996).
Jaubert, J. Les Chasseurs d'Aurochs de la Borde. Paris, France: Editions de la Maison des Sciences de l'Homme, 1990.
Kuhn, S. L. Mousterian Lithic Techology: An Ecological Perspective. Princeton, NJ: Princeton University Press, 1995.
Langley, M. C., Clarkson, C. & Ulm, S. Behavioural complexity in Eurasian Neanderthal populations: A chronological examination of the archaeological evidence. Cambridge Archaeological Journal 18, 289-307 (2008). doi:10.1017/s0959774308000371
Leroi-Gourhan, A. Étude des restes humains fossiles provenant des Grottes d'Arcy-sur-Cure. Annales de Paléontologie 44, 87-148 (1959).
Lévêque, F., Backer, A. M. & Guilbaud, M. Context of a Late Neandertal: Implications of Multidisciplinary Research for the Transition to Upper Paleolithic Adaptations at Saint-Césaire, Charente-Maritime, France. Madison, WI: Prehistory Press, 1993.
McBrearty, S. & Brooks, A. S. The revolution that wasn't: A new interpretation of the origin of modern human behavior. Journal of Human Evolution 39, 453-563 (2000). doi:10.1006/jhev.2000.0435
Mellars, P. A. Sequence and development of Mousterian traditions in south-western France. Nature 205, 626-627 (1965).
Mellars, P. A. The chronology of Mousterian industries in the Perigord region of south-west France. Proceedings of the Prehistoric Society 35, 134-171 (1969).
Mellars, P. A. The Neanderthal Legacy: An Archaeological Perspective from Western Europe. Princeton, NJ: Princeton University Press, 1996.
Mellars, P. A. The impossible coincidence. A single-species model for the origins of modern human behavior in Europe. Evolutionary Anthropology 14, 12-27 (2005). doi:10.1002/evan.20037
Monnier, G. F. The Lower/Middle Paleolithic periodization in Western Europe - An evaluation. Current Anthropology 47, 709-744 (2006).
Patou-Mathis, M. Neanderthal subsistence behaviours in Europe. International Journal of Osteoarchaeology 10, 379-395 (2000).
Plisson, H. & Beyries, S. Pointes ou outils triangulaires? Données fonctionelles dans le Moustérien Levantin. Paléorient 24, 5-24 (1998).
Rhodes, J. A. & Churchill, S. E. Throwing in the Middle and Upper Paleolithic: Inferences from an analysis of humeral retroversion. Journal of Human Evolution 56, 1-10 (2009). doi:10.1016/j.jhevol.2008.08.022
Richards, M. P. & Trinkaus, E. Isotopic evidence for the diets of European Neanderthals and early modern humans. Proceedings of the National Academy of Sciences of the United States of America 106, 16034-16039 (2009). doi:10.1073/pnas.0903821106
Richter, J. "When did the Middle Paleolithic begin?" in Neanderthal Lifeways, Subsistence and Technology: One Hundred Fifty Years of Neanderthal Study, eds. N. J. Conard & J. Richter (New York, NY: Springer, 2011) 7-14.
Riel-Salvatore, J. & Clark, G. A. Grave markers - Middle and early Upper Paleolithic burials and the use of chronotypology in contemporary Paleolithic research. Current Anthropology 42, 449-479 (2001).
Rolland, N. & Dibble, H. L. A new synthesis of Middle Paleolithic variability. American Antiquity 55, 480-499 (1990).
Schlanger, N. Understanding Levallois: Lithic technology and cognitive archaeology. Cambridge Archaeological Journal 6, 231-254 (1996).
Schmitt, D., Churchill, S. E. & Hylander, W. L. Experimental evidence concerning spear use in Neandertals and early modern humans. Journal of Archaeological Science 30, 103-114 (2003). doi:10.1006/jasc.2001.0814
Shea, J. J. Spear points from the Middle Paleolithic of the Levant. Journal of Field Archaeology 15, 441-456 (1988).
Shea, J. J. & Sisk, M. L. Complex projectile technology and Homo sapiens dispersal into western Eurasia. PaleoAnthropology 2010, 100-122 (2010).
Smith, F. H., Jankovic, I. & Karavanic, I. The assimilation model, modern human origins in Europe, and the extinction of Neandertals. Quaternary International 137, 7-19 (2005). doi:10.1016/j.quaint.2004.11.016
Sommer, J. D. The Shanidar-IV 'flower burial': A re-evaluation of Neanderthal burial ritual. Cambridge Archaeological Journal 9, 127-137 (1999).
Sorensen, B. Demography and the extinction of European Neanderthals. Journal of Anthropological Archaeology 30, 17-29 (2011). doi:10.1016/j.jaa.2010.12.003
Stiner, M. C. Paleolithic Diet and Demography: A 200,000 Year Record from Hayonim Cave (Levant). Cambridge, MA: Peabody Museum Press, Harvard University Press, 2005.
Stiner, M. C. "Middle Paleolithic subsistence ecology in the Mediterranean region," in Transitions Before the Transition: Evolution and Stability in the Middle Paleolithic and Middle Stone Age, eds. E. Hovers & S. L. Kuhn (New York, NY: Springer, 2006).
Stringer, C. B. et al. Neanderthal exploitation of marine mammals in Gibraltar. Proceedings of the National Academy of Sciences of the United States of America 105, 14319-14324 (2008). doi:10.1073/pnas.0805474105
Stringer, C. B. et al. "Climatic stress and the extinction of the Neanderthals," in Neanderthals and Modern Humans in the European Landscape During the Last Glaciation: Archeological Results of the Stage 3 Project, eds. T. H. van Andel & W. Davies (Cambridge, UK: McDonald Institute for Archaeological Research, Cambridge University Press, 2003) 233-240.
Van Andel, T. H. & Davies, W. eds. Neanderthals and Modern Humans in the European Landscape During the Last Glaciation: Archeological Results of the Stage 3 Project. Cambridge, UK: McDonald Institute for Archaeological Research, Cambridge University Press, 2003.
Villa, P. & Soriano, S. Hunting weapons of Neanderthals and early modern humans in South Africa: Similarities and differences. Journal of Anthropological Research 66, 5-38 (2010).
Zilhao, J. Neandertals and moderns mixed, and it matters. Evolutionary Anthropology 15, 183-195 (2006). doi:10.1002/evan.20110
Zilhao, J. & P. Pettitt. On the new dates for Gorham's Cave and the late survival of Iberian Neanderthals. Before Farming, 2006/3, article 3, pp. 95-122. 2006.
Zilhao, J. et al. Symbolic use of marine shells and mineral pigments by Iberian Neandertals. Proceedings of the National Academy of Sciences of the United States of America 107, 1023-1028 (2010). doi:10.1073/pnas.0914088107










