


















« Prev Next »

Understanding the origin (or origins) of modern humans is one of the central goals of paleoanthropology, but it remains a difficult goal to achieve. Part of this difficulty relates to problems of definitions. What do we mean by ‘modern human'? The term ‘modern human' is often used interchangeably with Homo sapiens. All living human populations belong to Homo sapiens. However, diversity is one of the hallmarks of our species, and living human populations are characterized by a wide range of anatomical and behavioral variation. So, how does a scientist settle on a definition of modernity that will include every member of our species past and present? For example, one approach to testing models of modern human origins is to identify where and when the earliest members of H. sapiens first appeared. However, problems arise when trying to recognize these earliest members of our species. Modern humans around the globe are highly variable, and studies of the skeletal remains have demonstrated that, even in the same geographic area, living populations show substantial morphological differences from populations of Homo sapiens sapiens living >5,000 years ago (Lahr 1996). How different or similar to living humans should we expect someone 200,000 years ago to be? Aside from how different they may have looked, should we expect them to have behaved the same ways as we do? More fundamentally, did these ancient hominins have the ability to think and behave the same as we do? Fortunately, fossil and archaeological evidence provide key sources of data to address some of these questions.
Approaches and Problems
Anatomy
All fossilized elements of the hominin body provide important clues about taxonomy and behavior. However, cranial and dental elements have dominated most discussions about modern human origins. In many cases, interpretations hinge on the attribution of a particular fossil or a suite of fossils to Homo sapiens or another taxon based on features of the skull or teeth. Among fossils of the genus Homo from the last 200,000 years or so, other elements (such as limb bones) rarely show features unique to a single species (e.g., Homo sapiens vs. Homo erectus). There has been considerable success using cranial features to distinguish among species. Some diagnostic cranial features of Homo sapiens include a high rounded brain case, a distinctive chin structure, and a gracile face with a canine fossa (a bony depression below the cheek bone where chewing muscles attach). One method of attributing a particular fossil to a species is by comparison of discrete traits. For example, a feature such as the presence of an inverted-T-shaped chin is present in living humans but absent from all other hominin species (Tattersall & Schwartz 2008). Another method of species attribution is the use of likelihood-based comparisons. Rather than simply determining if a trait is present, this method takes into consideration how frequently a particular trait is found among a population. This approach recognizes that different traits vary in their expression among living populations, and likely did in the past as well. For example, the middle trigonid crest (Figure 1), a particular morphological character of the lower molar teeth, is present in only 11% of Homo sapiens individuals, but this character is present in nearly 100% of Neanderthals (Bailey et al. 2011). 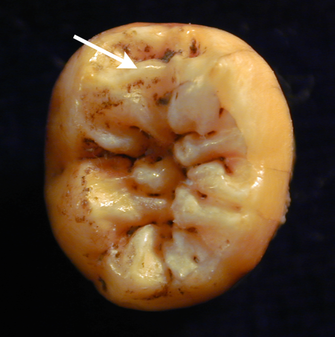
Figure 1: Lower first molar of a Neanderthal.
Lower first molar of a Neanderthal exhibiting a middle trigonid crest (orientation: crest is towards the front of the mouth).
© 2012 Nature Education All rights reserved. 
Paleoanthropologists must base their interpretations on rare items such as human fossils and archaeological sites that happen to be preserved and discovered. For fossils, in particular, paleoanthropologists make the simplifying assumption that the fossils they study — often of a single individual — are representative of the larger population to which they belonged during life. In practical terms, this can make defining — and subsequently recognizing — ancient human species difficult. For example, the type specimen of Homo heidelbergensis, a ~400,000 year old species predating Neandertals, is based on a single mandible. We have little idea what the corresponding cranium would look like. This gap makes it difficult for paleoanthropologists to agree on exactly which fossil individuals belong to this taxon, or the relationship of this species to later hominins (Rightmire 2008). This is particularly critical in the debate about the relationship between Homo sapiens and Neanderthals, a Eurasian population of hominins that some see as a distinct species (Homo neanderthalensis) and others as a regional population of modern humans (Homo sapiens neanderthalensis).
An analysis based on ancient DNA suggests that living populations of Homo sapiens and Neanderthals shared a relatively recent common ancestor around 300,000-500,000 years ago (Endicott et al. 2010). Many features of behavior (e.g., sophisticated tools), anatomy (e.g., large brain size), and the genome (Green et al. 2010) are quite similar between early Homo sapiens and the Neanderthals. Biological or behavioral features that are similar because of recent shared ancestry are termed ‘primitive,' although the recognition of primitive traits is rarely straightforward. A final problem is getting scientists to agree on which features of the skull or teeth are most important to the question at hand. Do primitive features observed in Homo sapiens indicate admixture with Neanderthals (Trinkaus et al. 2003) or are they simply features that were retained from an earlier ancestor (Hublin et al. 2012)? A similar conundrum arises when comparing modern human and Neanderthal genomes (Hodgson et al. 2010).
Archaeology
The archaeological record consists of items of material culture that are important clues to the daily lives of ancient human populations. Archaeological evidence records both changes through time as well as variation among contemporary populations in such things as subsistence methods (e.g., using traps to acquire small game, shellfish acquisition), environments inhabited (including occupation of distant islands such as the continent of Australia), and the use and long-distance movement of items of personal adornment such as shell beads (Marean 2010). 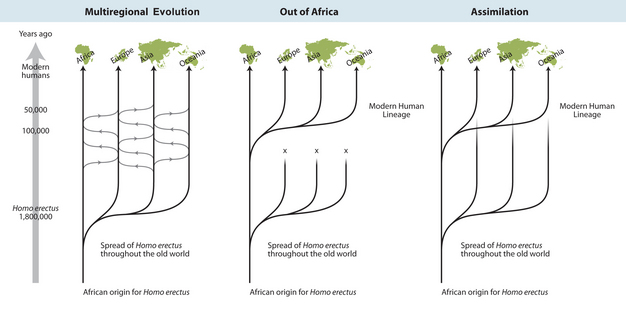
As with morphology, hominin diversity is also an issue for the archaeology of modern human origins. Multiple hominin species or populations may have overlapped in time and space, and rarely can the maker of the stone tools or other archaeological evidence be identified with any certainty. Furthermore, the extremely wide range of behaviors seen among recent and archaeologically recorded populations of hunter-gatherers is so great that defining archaeological criteria for behavioral modernity is difficult to impossible. The ‘trait list approach' employs a checklist of behaviors seen among modern hunter-gatherers for comparison with the archaeological record. Although it is a very logical approach, the system fails because evidence for many of these behaviors is absent from early archaeological sites attributed to Homo sapiens, such as those from Australia, where the earliest evidence of art is some 30,000 younger than the earliest archaeological sites on that continent (Brumm & Moore 2005). Other traits sometimes considered unique to modern humans — such as the use of pigments as colorants — were practiced by hominin populations often excluded from Homo sapiens, such as the Neanderthals (d'Errico &Stringer, 2011). Furthermore, historic populations of hunter-gatherers, such as those from Tasmania, have little in the way of material culture, yet are fully modern Homo sapiens sapiens (Heinrich 2004). Because the "things" found in the archaeological record can be a poor indicator of behavioral modernity, many archaeologists have turned to archaeological evidence for cognitive abilities, suggesting that superior cognitive abilities may be at the root of why our species persists where all other hominin populations have since gone extinct (Wynn & Coolidge 2008). However, even the apparently universal human capacity for symbolic behavior finds uneven archaeological expression, a topic dealt with in more detail below, and in The Transition to Modern Human Behavior (http://www.nature.com/scitable/knowledge/library/the-transition-to-modern-behavior-86614339).
The Competing Models
The ‘Out of Africa' (Replacement), ‘Multiregional Evolution' (Continuity), and ‘Assimilation' models are the three most widely used to interpret the origin of living human populations (Figure 2; Gibbons 2011). The Out of Africa model posits that all living humans (Homo sapiens sapiens) descend from a population that first appeared in Africa and subsequently spread throughout that continent and across the remainder of the globe, replacing existing hominin populations such as the Neanderthals. This would have occurred sometime between perhaps 90,000 and 60,000 years ago (Oppenheimer, 2012). Proponents of the Multiregional Evolution model suggest that hominin populations have maintained extensive social and biological ties across Africa and Eurasia for at least the last 1.8 million years, beginning when members of ancestral species such as Homo erectus dispersed from Africa. In this model, isolation among hominin populations created regional variation in behavior and biology, but occasional contact between groups through the sharing of ideas (cultural transmission) and mates (gene flow) was sufficient to prevent the formation of distinct biological species.
Figure 2: Diagram showing each of the three competing models.
© 2013 Nature Education Modified from Gibbons. All rights reserved.
Assimilation models combine elements of the other two models, and their adherents favor an African origin of Homo sapiens with varying degrees of integration through interbreeding with existing populations as Homo sapiens spread from their place(s) of origin. Evidence needed to support or refute these models include the age and location of the earliest fossils assigned to Homo sapiens, the species attribution and fate of pre-existing hominin populations (such as the Neanderthals), and the extent to which evidence for interbreeding can be found in the fossil record. Analyses of modern and ancient DNA provide increasingly important data (e.g., Green et al. 2010).
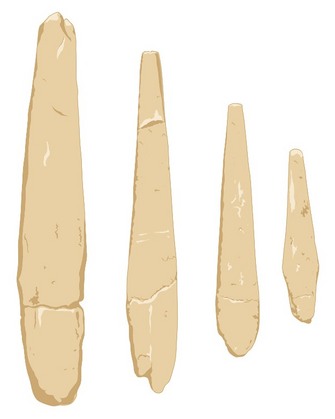
Some paleoanthropologists add an additional complication: Did anatomical modernity and behavioral modernity appear separately, or did they arise simultaneously? According to one view, modern human form preceded the appearance of what we would recognize as modern cognitive abilities. These abilities include the extensive use of symbols, both verbally, as fully developed syntactical language, and visually, as art and personal adornment, as well as the development of complex extended social networks. Richard Klein (2009) suggests that behavioral modernity arose as an advantageous genetic mutation resulting in neural reorganization that spread rapidly among existing populations of anatomically modern humans perhaps 50,000 years ago. The alternative view is that the capacity for modern human behavior is coincident with the origins of Homo sapiens, and that the expression of this capacity in the archaeological record was an incremental process developed through time (McBrearty & Brooks 2000, Shea 2011, Wong 2005). The data needed to test these models includes artifacts and other elements of the archaeological record; that is, the material traces of past human behavior (Tryon et al. 2010). These include objects and tools of bone, stone, antler, and shell, evidence for how they were made and used, and features such as hearths, that show control of fire and modification of the environment. (Figure 3).
Which Model is Best Supported by the Evidence?
At present, the oldest hominin fossils attributed to Homo sapiens occur in Africa, and are found in sediments of the Kibish Formation in southern Ethiopia. Dated to approximately 195,000 years ago (McDougall et al. 2005), the two partial skeletons (Omo I and II) are at least 60,000 years older than the oldest Homo sapiens fossils found outside of Africa, at Skhul and Qafzeh in Israel. The fossils from the Kibish Formation, as well as others, such as crania from Herto, Ethiopia and Jebel Irhoud, Morocco (Smith et al. 2007), dating to approximately 160,000 years ago (Clark et al. 2003, White et al. 2003), are most consistent with an African origin for Homo sapiens. As regards the fate of archaic populations who were already present in the Old World at the time of Homo sapiens' expansions out of Africa, the jury is still out. While it is clear that Neandertals and their kin are no longer around, recent genetic evidence suggests some admixture between them and Homo sapiens at around the time of initial contact (Hodgson et al. 2010). However, while these and other recent genetic analyses are consistent with the Assimilation model, morphological evidence for archaic (e.g., Neandertal) and modern (e.g., H. sapiens) hybrids is controversial (Pearson 2004).
At present, the African archaeological record shows little obvious change at 200,000 years ago with the appearance of Homo sapiens; evidence for symbolic behavior occurs only at sites younger than that age (Wong 2005, Marean 2010). However, the Eurasian archaeological record shows a number of abrupt changes broadly coincident with the appearance of fossils of Homo sapiens on that continent ~40,000 to 30,000 years ago, particularly with the appearance of distinctive Aurignacian archaeological sites. These changes include the types of stone tools used and the ways in which they were made, as well as increased frequency of symbolic behavior and long-distance movement of materials. These differences are consistent with the influx of new populations predicted by the Out of Africa model (Mellars 2005), but have also been interpreted as the rapid spread of new technologies or behaviors among existing populations predicted by the Multiregional Evolution model (Wolpoff et al. 2004). Some regional artifact industries, such as the Châtelperronian, have been considered as the outcome of some form of interaction between populations of Neanderthals and Homo sapiens, consistent with the Assimilation model (Mellars 1996, 2010).
Paleoanthropologists are increasingly recognizing the important role of demography in testing models of human origins. Increases in population size and local population density are probably the root cause for dispersal of some populations of Homo sapiens out of Africa (Mellars 2006). Archaeological evidence demonstrates over-hunting of animal populations resulting from dense populations of early humans as much as 41,000 years ago (Kuhn et al. 2009). Several recent studies have shown that larger and denser human populations are more likely to invent and retain many of those behaviors considered by some to be unique to modern humans (Heinrich 2004, Powell et al. 2009). Importantly, several lines of evidence suggest that Neanderthal populations were small and widely dispersed, suggesting that the absence of abundant archaeological manifestations of evidence of "modern" human behavior among Neanderthals, such as art, may be a result of demography rather than inability (Premo & Kuhn 2010, Mellars & French 2011). Clearly, cognitive definitions of modern humans are complex and difficult to define.
There remains little consensus on the timing and tempo of the evolution of modern humans, and new fossil and archaeological data continually reshape, revise, and complicate current models and hypotheses. For example, early European representatives of Homo sapiens can be distinguished morphologically from preceding and contemporary populations of Neanderthals, and are associated with distinctive Aurignacian artifacts such as spear points made of antler (Figure 3). However, although fossil and genetic data are consistent with an African origin for our species, the Aurignacian has no clear antecedents in the African archaeological record, and probably first appears in the eastern Mediterranean region of Eurasia (Mellars 2005). Thus, some of the earliest, non-Neanderthal, European populations attributed to Homo sapiens combine a recent African biological heritage with behavioral traditions rooted in Eurasia. This emphasizes that biological and behavioral evolution are related in complex ways, and that the suite of morphological and behavioral characteristics of living humans may be the outcome of a far messier reality than any of the current models propose.
References and Recommended Reading
Bailey S. E., Skinner, M. & Hublin, J-J. What lies beneath? An evaluation of lower molar trigonid crest patterns based on both dentine and enamel expression. American Journal of Physical Anthropology 45, 505-518 (2011).
Brumm, A. & Moore, M. W. Symbolic revolutions and the Australian archaeological record. Cambridge Archaeological Journal 15, 157-175 (2005).
Clark, J. D. et al. Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423, 747-752 (2003).
d'Errico, F. & Stringer, C. B. Evolution, revolution or saltation scenario for the emergence of modern cultures? Philosophical Transactions of the Royal Society B: Biological Sciences 366, 1060-1069 (2011).
Endicott, P., Ho, S. Y. & Stringer, C. Using genetic evidence to evaluate four palaeoanthro-pological hypotheses for the timing of Neanderthal and modern human origins. Journal of Human Evolution 59, 87-95 (2010).
Gibbons, A. A new view of the birth of Homo sapiens. Science 331, 392-394 (2011).
Green, R. E. et al. A draft sequence of the Neanderthal genome. Science 328, 710-722 (2010).
Heinrich, J. Demography and cultural evolution: How adaptive cultural processes can produce maladaptive losses: The Tasmanian case. American Antiquity 69, 197-214 (2004).
Hodgson, J. A., Bergey, C. M. & Disotell, T. R. Neanderthal genome: The ins and outs of African genetic diversity. Current Biology 20, R517-R519 (2010).
Hublin, J-J. et al. "Dental evidence from the Aterian human populations of Morocco," in Modern Origins: A North African Perspective, eds. J-J. Hublin, & S. P. McPherron ( Springer, 2012 pp-pp.
Klein, R. G. The Human Career, 3rd ed. Chicago, IL: University of Chicago Press, 2009.
Kuhn, S. L. et al. The early Upper Paleolithic occupations at Üçagizli Cave (Hatay, Turkey). Journal of Human Evolution 56, 87-113 (2009).
Lahr, M. M. The Evolution of Modern Human Cranial Diversity: A Study in Cranial Variation. Cambridge, UK: Cambridge University Press, 1996.
Marean, C. W. When the sea saved humanity. Scientific American 303, 54-61 (2010).
McBrearty, S. & Brooks, A. The revolution that wasn't: A new interpretation of the origin of modern human behavior. Journal of Human Evolution 39, 453-563 (2000).
McDougall, I., Brown, F. H. & Fleagle, J. G. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433, 733-736 (2005).
Mellars, P. The Neanderthal Legacy. Princeton, NJ: Princeton University Press, 1996.
Mellars, P. The impossible coincidence: A single-species model for the origins of modern human behavior in Europe. Evolutionary Anthropology 14, 12-27 (2005).
Mellars, P. Why did modern human populations disperse from Africa ca.60,000 years ago? A new model. Proceedings of the National Academy of Sciences of the United States of America 103, 9381-9386 (2006).
Mellars, P. Neanderthal symbolism and ornament manufacture: The bursting of a bubble? Proceedings of the National Academy of Sciences of the United States of America 107, 20147-20148 (2010).
Mellars, P. & French, J. C. Tenfold population increase in western Europe at the Neandertal-to-modern human transition. Science 333, 623-627 (2011).
Oppenheimer, S. A single southern exit of modern humans from Africa: Before or after Toba? Quaternary International 258, 88-99 (2012). doi:10.1016/j.quaint.2011.07.049
Pearson, O. Has the combination of genetic and fossil evidence solved the riddle of modern human origins? Evolutionary Anthropology 13, 145-159 (2004).
Powell, A., Shennan, S. & Thomas, M. G. Late Pleistocene demography and the appearance of modern human behavior. Science 324, 1298-1301 (2009).
Premo, L. & Kuhn, S. L. Modeling effects of local extinctions on culture change and diversity in the Paleolithic. PLoS ONE 5, e15582 (2010).
Rightmire, G. P. Homo in the middle Pleistocene: Hypodigms, variation, and species recognition. Evolutionary Anthropology 17, 8-21 (2008).
Shea, J. J. Homo sapiens is as Homo sapiens was. Current Anthropology 52, 1-35 (2011).
Smith, T. M. et al. Earliest evidence of modern human life history in North African early Homo sapiens. Proceedings of the National Academy of Sciences USA 104, 6128-6133 (2007).
Trinkaus E. et al. An early modern human from the Pestera cu Oase, Romania. Proceedings of the National Academy of Sciences of the United States of America 100, 11231-11236 (2003).
Tryon, C., Pobiner, B. & Kauffman, R. Archaeology and human evolution. Evolution: Education and Outreach 3, 377-386 (2010).
White, T. D. et al. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423, 742-747 (2003).
Wolpoff, M. H. et al. Why not the Neandertals? World Archaeology 36, 527-546 (2004).
Wong, K. The morning of the modern mind. Scientific American 292, 86-95 (2005).
Wynn, T. & Coolidge, F. A Stone Age meeting of minds. American Scientist 96, 44-51 (2008).










