
Abstract
Archaeogenetic studies have described two main genetic turnover events in prehistoric western Eurasia: one associated with the spread of farming and a sedentary lifestyle starting around 7000–6000 bc (refs. 1,2,3) and a second with the expansion of pastoralist groups from the Eurasian steppes starting around 3300 bc (refs. 4,5). The period between these events saw new economies emerging on the basis of key innovations, including metallurgy, wheel and wagon and horse domestication6,7,8,9. However, what happened between the demise of the Copper Age settlements around 4250 bc and the expansion of pastoralists remains poorly understood. To address this question, we analysed genome-wide data from 135 ancient individuals from the contact zone between southeastern Europe and the northwestern Black Sea region spanning this critical time period. While we observe genetic continuity between Neolithic and Copper Age groups from major sites in the same region, from around 4500 bc on, groups from the northwestern Black Sea region carried varying amounts of mixed ancestries derived from Copper Age groups and those from the forest/steppe zones, indicating genetic and cultural contact over a period of around 1,000 years earlier than anticipated. We propose that the transfer of critical innovations between farmers and transitional foragers/herders from different ecogeographic zones during this early contact was integral to the formation, rise and expansion of pastoralist groups around 3300 bc.
Similar content being viewed by others

Main
During the fifth and fourth millennia bc, key technological and social changes took place in southeastern Europe (SEE) which profoundly transformed prehistoric societies. Metal production was among the most important innovations; copper was mined, smelted and used to make axes, jewellery and small tools. The discovery of the necropolis of Varna (4600–4300 bc) on the Black Sea coast led to a reassessment of social inequality in human prehistory, with large quantities of gold and other symbols of power and wealth suggesting unprecedented levels of social stratification10,11,12. The many tell settlements that emerged during the Copper Age (CA, 4900–3800 bc) in SEE, involved in the proto-industrial exploitation of copper13, gold and salt, highlight this advanced social organization and the blossoming of social, political, economic and artisanal activities. Eminent tell sites include Mound Măgura Gorgana near Pietrele on the Lower Danube in Romania14, associated with the Gumelniţa culture and Tell Yunatsite in Bulgaria, associated with the Karanovo culture (Fig. 1 and Extended Data Fig. 1), which were occupied for several centuries15. From around 4600 bc, the similarity and continuous development of material culture and exchange of raw materials in the so-called Gumelniţa–Kodžadermen–Karanovo VI complex across southern Romania (Gumelniţa), northern Bulgaria (Kodžadermen) and Thrace (Karanovo) indicate transregional connectedness and suggest a relatively stable sociopolitical network. Consequently, the roughly simultaneous abandonment of the numerous tell settlements and cemeteries around 4250/4200 bc appears enigmatic (Fig. 1a,c). The underlying circumstances are unclear and might have involved the depletion of resources, the deterioration of soils and possibly also violent conflicts, as evidenced by the destruction horizon at Tell Yunatsite16,17. Historically, this demise was associated with the arrival of new groups from the steppe18 but this proposal has lacked sufficient evidence. However, settlement activity over the following centuries was scarce in the entire western Black Sea region, indicating a ‘dark’ millennium with, for example, Yunatsite not being resettled until about 1,000 years later during the Early Bronze Age (EBA)15.
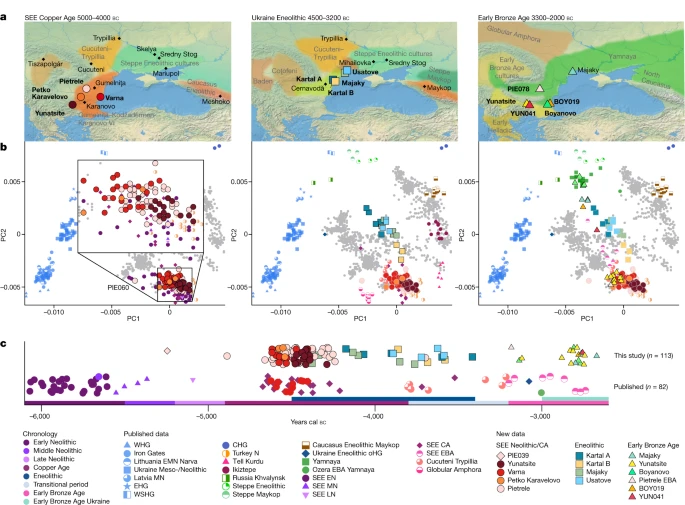
a, Map of sites and relevant archaeological cultures discussed in the text. Maps were made with Natural Earth (https://naturalearthdata.com). b, PCA of newly reported individuals (coloured symbols with black outline) and relevant published groups (coloured symbols, no outline) projected onto the West Eurasian genetic variation of 1,253 individuals from 77 populations. c, Mean radiocarbon dates of relevant published and newly reported individuals from southeastern Europe plotted according to the regional chronology. The suffixes in the group labels present archaeological time periods and geographical regions: N, Neolithic; EN, MN, LN, Early, Middle, Late Neolithic, respectively; CA, Copper Age; EBA, Early Bronze Age; SEE, southeastern Europe; WHG, EHG, WSHG, CHG, oHG, Western, Eastern, West Siberian, Caucasus, outlier Hunter–Gatherers, respectively.
Following the end of the CA, the centre of settlement activity shifted further northeast towards the forest–steppe region, where huge settlements, with thousands of houses, the so-called megasites of the Cucuteni–Trypillia complex (around 4100–3800 bc), emerged19,20. This northwestern Black Sea region represents an interaction zone between late CA farming-associated groups and those of the adjacent steppe region with different ecogeographic conditions. Continued innovations increased human mobility and the exploration of lands hitherto not amenable to agrarian lifestyles, as practised in the regions in SEE and south of the Caucasus for many millennia before. From the contact zones in the northwestern Black Sea region and the Caucasus, a gradual transition from foraging to semi-nomadic pastoralism also followed in the North Pontic region during the sixth and fourth millennium bc, triggered by continued innovations6, transfer of livestock and advances in herd management, food processing, dairying practices8,9,19 and the development of arsenical-copper alloys21. The North Pontic region played a central role in the development of the oldest wheeled vehicles22, while the North Caucasian Maykop culture was critical in the further development of metal alloys, as well as early horse domestication8,9 and a sheep-wool economy combined with extensive dairy production9. The Maykop culture had extremely rich burials with metal weapons indicating ‘high status’ individuals, attesting to social inequality and upheaval during this time23, as such social elites were also found in southern Romania and Bulgaria24. The Cernavodă I (around 4000–3200 bc) and Usatove cultures (3600/3500–3200/3100 bc) in the northwestern Black Sea region played a chief role in the east–west exchange between the Danube and the lower Dniepr25 and these formations, while presumably indigenous, received strong contributions from the Trypillian tradition26.
Similar to the SEE CA tell sites, the megasites and cultural phenomena of the northwestern Pontic region suddenly disappeared and were succeeded around 3300 bc by fully established pastoralists associated with the Yamnaya cultural complex. The expansion of North Pontic pastoralists to the west has been studied in many regions of Europe in recent years27, whereas their emergence and impact on societies in SEE is barely understood. This is relevant insofar as the archaeological record for the subsequent EBA (around 3200–2500 bc), indicates a concomitant rise in settlement activity for the first time since the demise of the CA settlements in the eastern Balkan region28. Burial mounds, associated with the Yamnaya cultural complex, appear frequently and extend along the Danube valley into the Carpathian Basin during the third millennium bc (refs. 4,29). By contrast, the resettlement of sites like Tell Yunatsite involved groups with burial rites not associated with the incoming steppe groups30 (Extended Data Fig. 1c,h).
Archaeogenomic studies have shown that SEE CA individuals carry genetic profiles that resemble those of Neolithic farmers expanding from western Anatolia into Europe31, distinct from both the earlier pre-agropastoralist (Western and Eastern Hunter–Gatherers; WHG/EHG) and later EBA pastoralist groups1,2,4,5, who carried ‘steppe’ ancestry. Individuals from well-known, contemporaneous CA settlements (Pietrele and Yunatsite) and outstanding burial sites (Varna) provide a unique opportunity to study the genetic variation in and between sites at their peak settlement densities. However, the developments following early interactions, which had later given rise to the expansion of pastoralists and their genetic ancestry across Europe, remain unknown. Critically, individuals from the key period of the fifth and fourth millennium bc from the contact zone between SEE, the Trypillian megasites and the steppes have not been analysed genetically. Here, we address this spatial and temporal sampling gap by studying individuals associated with the Cernavodă I and Usatove cultures from the northwestern Black Sea region in today’s Ukraine. Additionally, we analyse EBA individuals from the tell sites Yunatsite and Pietrele, following a possible resettlement of the sites after several centuries of abandonment. We compare these to Yamnaya-associated individuals from eastern Bulgaria, who were buried in mounds typically associated with steppe pastoralists during the third millennium BC and to individuals postdating the Usatove horizon in the northwestern Black Sea area.
In total, we report genome-wide data for 135 (out of 216 attempted) individuals from eight distinct sites (Fig. 1) ranging from around 5400 to 2400 bc: Neolithic (n = 1), CA (n = 95), Eneolithic (n = 18) and EBA (n = 21). All samples were enriched for a panel of 1.24 million single-nucleotide polymorphisms (1,240,000 SNP panel32), ranging from 61,000 to 947,000 SNPs with an average SNP coverage between 0.01× and 3.4×. We used a cut-off of 400,000 SNPs for hapROH and imputation and filtered for >550,000 SNPs for identity-by-descent (IBD) analyses (Supplementary Table A; Methods). We also report 113 new radiocarbon dates (Fig. 1c and Supplementary Table A). To assess the genetic ancestry and variation of the newly typed individuals we first performed principal component analysis (PCA) constructed from 1,253 modern-day West Eurasians from 77 different populations, onto which data from the ancient individuals were projected (Fig. 1b and Supplementary Table B; Methods).
Neolithic and Copper Age ancestries
The earliest-dated individual in our dataset, PIE039 from Pietrele, falls in the expected range of other SEE Neolithic individuals in PCA space, with whom she also shares affinities according to outgroup f3 statistics (Fig. 1b, Fig. 2 and Supplementary Table C). We used f4 statistics of the form f4 (test, PIE039; HGs, Mbuti), where ‘test’ are different Neolithic groups, to identify the genetically most similar Neolithic groups, which were then used as local proxies for quantitative ancestry modelling. We found Hungary_LN_Sopot and Malak Preslavets N to be most symmetrically related to PIE039 with respect to all HG comparisons (|Z| ≤ 1) and thus combined them into local group SEE 1, which could be used as a single source for proximal qpAdm modelling (P = 0.41), confirming shared local ancestry (Extended Data Fig. 2, Fig. 3d, Supplementary Tables D, E, H and Supplementary Information 5).
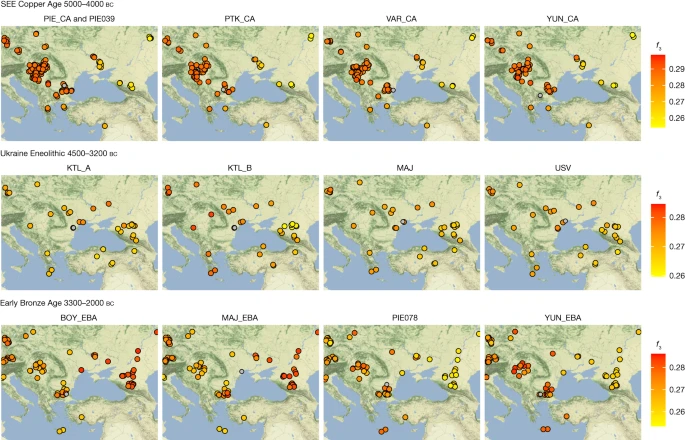
Outgroup f3 statistics of the form f3(test1, test2; Mbuti) plotted onto maps of central/SEE highlighting the shifting genetic affinities over time, separated temporally. Test1 includes groups and individuals newly reported in this study (headers) and their respective location is given by a grey circle. Test2 includes all relevant ancient populations from the respective time period (Supplementary Table C) and their locations are indicated as filled circles with black outlines. Higher f3 statistics (red colours) indicate more shared drift with the respective group in Test1. All f3 statistics, including outlier individuals and published ancient populations further west, can be found in Supplementary Table C. The maps were made in R57 and the tile sets are copyright of Stamen Design, under a Creative Commons Attribution (CC BY 3.0) licence.
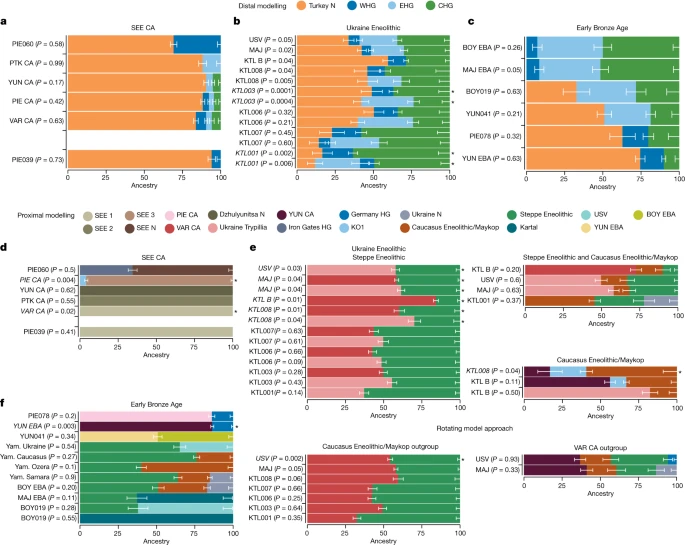
a–c, Distal models with Turkey_N, WHG, EHG and CHG as sources for the three sampled time periods: SEE CA (a); Ukraine Eneolithic (b); and Early Bronze Age (c). d–f, Geographically and temporally proximal models of the three sampled time periods: SEE CA (d); Ukraine Eneolithic (e); and Early Bronze Age (f) (Supplementary Tables G, H, P, Q, V and W). All results shown here were run with the parameter ‘allSNPs: NO’ (Supplementary Information 6). * Indicates non-supported/rejected/failed models when applying a P value cut-off of less than 0.05 (shown in italics). Ancestry proportions are shown with one standard error. Standard errors were computed with the default block jackknife approach.
In PCA space, the chronologically younger SEE CA individuals from the emblematic sites of Yunatsite (YUN), Varna (VAR), Pietrele (PIE) and the multiple burial from Tell Petko Karavelovo (PTK), form a tight cluster that also overlaps with published Neolithic individuals from Anatolia and SEE29 (Fig. 1b). Moreover, outgroup f3 statistics suggest local genetic homogeneity throughout the CA in this region (Fig. 2 and Supplementary Table C). However, all SEE CA groups are slightly shifted towards the EHG/WHG cline in both PC1 and PC2 compared to most published Neolithic individuals. Distal qpAdm modelling (Fig. 3a and Supplementary Table G) confirmed minimal amounts of EHG-, CHG- and WHG-like ancestry, in addition to predominantly Turkey_N-like ancestry. This ancestry composition is already present during the Neolithic29 and confirmed by the test f4 (test, CA; HGs, Mbuti) in which Neolithic groups form a clade with SEE CA with respect to HG groups (Extended Data Fig. 2, Supplementary Tables D and E and Supplementary Information 5). This allows us to identify the best local Neolithic proxy for each SEE CA group and to account for the subtle differences in ancestries. Using the respective, locally preceding, Neolithic groups for proximal qpAdm modelling, we could model all SEE CA groups as a single-source model (Fig. 3d and Supplementary Table H), suggesting genetic continuity at the local scale.
The outlier individual PIE060 is shifted further towards the WHG/EHG cluster in PCA, suggesting an excess of this type of ancestry, which could be confirmed by f4 statistics of the form f4(SEE N, PIE060; HGs, Mbuti) ((|Z| ≥ 3); Supplementary Table F). Ancestry modelling with qpAdm supports a two-way model (Fig. 3d) with SEE N (around 65%) and Iron Gates HG or KO1 (around 35%) as the best proxies. Using DATES33 to determine the time of admixture between SEE N and Iron Gates HG as a local HG ancestry, we obtained an admixture estimate of 16.3 ± 13.4 generations (Z = 1.213), which corresponds to around 81–832 years before the mean 14C date of PIE060, when a generation time of 28 years is assumed34. A flat decay curve (Extended Data Fig. 3a) supports the interpretation of a recent admixture date, which suggests that PIE060 came from a community outside Pietrele with recent contact with HGs. Indeed, individuals with similarly high amounts of HG ancestry have been reported from nearby sites in Malak Preslavets (around 70 km) and Dzhulyunitsa (around 140 km)29.
In line with the autosomal data, the Y-chromosomal and mitochondrial DNA lineages are common in nearly all Neolithic and CA groups studied until now, albeit with several males also carrying typical Mesolithic (C1a and I2a) Y lineages35, including individual PIE060 (Extended Data Fig. 3b and Supplementary Table A). With seven different main lineages among 29 males in Pietrele (I2a1, C1a, G2a, H2, T1a, J2a and R1b-V88), six among 15 males in Varna (I2a1, I2a2, G2a, T1a, E1b1 and R1b-V88) and four among six males at Yunatsite (C1a, G2a, H2, J2a), the Y-chromosomal diversity during the SEE CA was higher than in central/western Europe36,37,38.
When testing for genetic relatedness in each of the SEE CA sites using READ, we detected only three first-degree and two second-degree relationships in total (Supplementary Table I; Methods). To specifically test for links between the contemporaneous SEE CA sites and for more distant genetic relatedness we explored signals of IBD sharing between individuals in and between all sites (Methods). We found no evidence for between-site links up to the fourth to fifth degree and only two pairs of individuals (PIE003-VAR010 and YUN005-VAR030) shared at least two blocks greater than 20 cM indicative of a fifth to seventh degree relationship (Extended Data Fig. 4a and Supplementary Table J). Integrating the normalized sum and number of shared blocks we find higher background relatedness at the intrasite level at Yunatsite and Varna compared to Pietrele, which can be explained by the structure of the sites (a destruction horizon of households and a burial ground with shorter use, respectively, versus tell and settlement burials spanning 350–400 years) (Fig. 1c and Extended Data Fig. 4b). However, analysis of the runs of homozygosity (ROH) per individual using hapROH indicates low levels of parental background relatedness suggesting relatively large effective population sizes, consistent with previous observation across early farming societies (Methods; Extended Data Fig. 5). These findings reflect the settlement density and the wide-spread cultural, rather than close genetic, connectedness of the Gumelniţa–Kodžadermen–Karanovo VI complex, in line with the cross-regional significance of SEE tell sites26.
Early contacts during the Eneolithic
Eneolithic individuals from Ukraine (Ukraine Eneolithic), dated from around 4500–3500 bc, associated with the Cernavodă I and Usatove cultures, form a genetic cline in PCA space (Fig. 1b) between Neolithic/SEE CA individuals and published Eneolithic steppe individuals from the North Caucasus39 and Khvalynsk in western Russia32. This indicates possible admixture between CA farmer-related groups and Eneolithic steppe groups, as in line with cultural interactions described in the archaeological record40,41,42. The observed genetic cline reflects developments over a wide chronological range of around 1,000 years (Fig. 1c and Supplementary Table A). Some of the newly reported 14C dates could be affected by a freshwater reservoir effect43, common in Steppe Eneolithic sites44,45 and could therefore be several centuries younger than their reported dates. However, accounting for this possibility, an offset of around 500 years would still date most of the Ukraine Eneolithic individuals to the fourth millennium bc and thus considerably earlier than the Yamnaya-associated steppe pastoralist expansion.
Individuals from Kartal (around 4150–3400 bc), associated with the Cernavodă I culture, are genetically highly heterogeneous, with five individuals (Kartal A) forming a cline between ‘Steppe Eneolithic’/‘Steppe Maykop’ individuals and Early Neolithic groups, while three other individuals (Kartal B) fall closer to the latter (Supplementary Tables L and M). The five contemporaneous individuals from Majaky (MAJ), are genetically more homogeneous and fall together with the four individuals from the late Eneolithic Usatove type-site (USV/UBK; Supplementary Table A) in the middle of the ‘Kartal cline’. We tested for a correlation between positions of the Ukraine Eneolithic individuals in PC2 and their 14C dates and found none (Spearman’s ρ = 0.113, P = 0.6656). The broadscale shift in genetic affinities between the CA and the Eneolithic, from SEE to the steppe zone, is also clearly visible in outgroup f3 statistics when mapped geographically (Fig. 2 and Supplementary Table C).
To formally characterize the Ukraine Eneolithic individuals, we tested for excess shared ancestry with four Holocene ‘cornerstone’ populations (Turkey_N, WHG, EHG/WSHG and CHG) (Supplementary Information 1.2), using f4-symmetry statistics of the form f4(test, Ukraine Eneolithic; cornerstone, Mbuti) and conditioning on three test populations (Extended Data Fig. 6, Supplementary Tables E, M and N and Supplementary Information 6). First, compared to Turkey_N, Ukraine Eneolithic individuals show excess affinity to all HG groups, as indicated by significantly negative f4 statistics (|Z| ≥ 3) (Extended Data Fig. 6a). Second, conditioning on Steppe Eneolithic (Extended Data Fig. 6b), we observe excess affinity of Ukraine Eneolithic to Turkey_N, a symmetrical relatedness to CHG and WHG, while Steppe Eneolithic groups carry more EHG/WSHG ancestry. On the basis of cultural influences which also link the northern Black Sea through the steppe belt to the North Caucasus region40,41,42, we also test for potential influence of North Caucasian groups. Using Caucasus Eneolithic/Maykop as test (Extended Data Fig. 6c) we find excess affinity of Ukraine Eneolithic to EHG and WHG and Turkey_N, while Caucasus Eneolithic/Maykop share more drift with CHG.
The archaeological record identifies the northwestern Black Sea region as an interaction zone between late CA farming and groups from the steppe region19,20. Such an early interaction has been postulated by Immel et al.46, who have reported Yamnaya-related ancestry in individuals associated with the Cucuteni–Trypillia complex from today’s Moldova. However, on re-analysis of these data we find that this signal can be explained solely by an increase in especially EHG-rich ancestry (Supplementary Information 4 and Supplementary Table O).
To characterize the role of Cernavodă I and Usatove-associated individuals from the postulated interaction zone, who show a clear signal of admixture, we formally tested the contribution of diverse ancestry sources using f4(Steppe Eneolithic/Caucasus Eneolithic/Maykop, Ukraine Eneolithic; test, Mbuti), where test represents SEE and Anatolian CA farmer groups (Supplementary Table P). With respect to Steppe Eneolithic, all Ukraine Eneolithic individuals show excess affinity to all tested CA groups. With respect to Caucasus Eneolithic/Maykop, USV, MAJ, KTL_B, KTL003 and KTL008 show excess affinity to all SEE CA farmer groups, while KTL006 and KTL007 only share drift with Ukraine Trypillia (Supplementary Table P).
Of note, all f4-symmetry tests with Caucasus Eneolithic/Maykop and SEE CA indicate an additional attraction of Ukraine Eneolithic to WHG/EHG (Supplementary Table N), with Iron Gates HG or Ukraine N showing the highest affinity (Supplementary Table Q). This affinity towards WHG/EHG is absent when Steppe Eneolithic is used (Supplementary Table N), implying that scenarios involving potential gene flow from the Caucasus would require an additional source carrying WHG-/EHG-like ancestry as this ancestry is not sufficiently represented by SEE CA or Caucasus Maykop groups.
Using distal qpAdm modelling we find support for a four-way admixture of Turkey_N, EHG, CHG and WHG for KTL001, KTL007, MAJ and USV (Fig. 3b and Supplementary Table R), while individuals KTL003, KTL006 and KTL008 can be modelled alternatively with three sources (Turkey_N+EHG+CHG) and KTL_B individuals only with Turkey_N (around 60%), CHG (around 28%) and WHG (around 12%) ancestry. Following up with proximal qpAdm models to explore potential contribution(s) of temporally and geographically closer groups (Fig. 3e and Supplementary Tables E and S), we find that all Ukraine Eneolithic individuals can be modelled as a two-way model of either VAR_CA or Ukraine Trypillia as farmer-related ancestry source and Steppe Eneolithic as a source of mixed EHG+CHG ancestry.
Since archaeological research suggests a cultural contribution of Steppe Eneolithic and Maykop groups (Supplementary Information 2.2), we specifically tested for alternative scenarios which involved admixture between both groups north of the Caucasus and subsequent spread westwards. Using both associated ancestries and different HGs and SEE CA-related groups as sources in qpAdm modelling (Fig. 3e and Supplementary Table S), we find that KTL001 can indeed be modelled as a three-way mixture of Steppe Eneolithic (around 32%), Caucasus Eneolithic/Maykop (around 46%) and Ukraine N foragers (around 22%), to the exclusion of a SEE CA source. By contrast, MAJ and USV can be modelled as VAR_CA or Ukraine Trypillia (around 50%), Steppe Eneolithic (around 35%) and Caucasus Eneolithic/Maykop (around 15%) as minor third component. KTL_B results in the same model but with a higher VAR_CA component (around 73%) and a minor contribution of Steppe Eneolithic (around 10%) ancestry (Supplementary Table S).
Exploring an alternative scenario which excludes Steppe Eneolithic as a source, we find a well-fit model for KTL008 with YUN_CA (around 17%), Caucasus Eneolithic/Maykop (around 60%) and KO1 (around 23%). Further, KTL_B can be modelled with Ukraine Trypillia (around 82%) and Caucasus Eneolithic/Maykop (around 18%) as second source, which is consistent with the omission of EHG ancestry in the distal qpAdm results for KTL_B (Fig. 3e and Supplementary Table S).
Finally, to test whether we can distinguish between the farmer-related ancestry contributed by SEE CA- or Maykop-associated groups from the Caucasus, or by both, we rotated each source to the outgroups, alternatingly, keeping Steppe Eneolithic as a constant. Here, we find strong support for a genetic contribution from SEE CA rather than Caucasus Eneolithic/Maykop for most KTL individuals (except KTL_B), which can be modelled as Steppe Eneolithic and VAR_CA (Fig. 3e and Supplementary Table S). The same model is supported for MAJ (P = 0.05) but rejected for USV, which indicates that Maykop-associated ancestry is needed for the latter. Indeed, the competing model, with Maykop as an additional source and VAR_CA as an outgroup, results in a well-fit four-way mixture model for USV (P = 0.93) and improved model fit for MAJ (P = 0.33), whereas the models for the remaining KTL individuals are rejected (Supplementary Table S). This provides strong support for an alternative admixture history for USV and MAJ, involving local SEE CA, Steppe Eneolithic, Caucasus Eneolithic/Maykop and a HG-related source, a combination that is distinct from KTL individuals.
The similarities in genetic ancestry presented for MAJ and USV are also observed in the results from the IBD analysis (Extended Data Fig. 4a and Supplementary Table J) in which we find a fourth to sixth degree relationship between MAJ023 and USV006, which reflects the close geographical vicinity of the two sites. The normalized sum and number of shared blocks for Ukraine Eneolithic show a higher background relatedness in USV compared to the other sites (Extended Data Fig. 4b) but also between USV and MAJ and USV and KTL, respectively, which matches the relative chronological overlap of the three sites (Fig. 1c and Supplementary Table A). However, in comparison to the preceding CA and heterogenous KTL individuals, ROH indicate a slightly elevated parental background relatedness for MAJ and USV (Extended Data Fig. 5), suggesting smaller effective population sizes in Usatove-associated groups.
Y-chromosomal evidence from the six Ukraine Eneolithic males reflects lineages from each of the contributing sources (Extended Data Fig. 3b): G2a is probably a Neolithic legacy, while three males carrying I2a1 could be attributed to the local Ukrainian Neolithic or HG groups in general. KTL005 and MAJ009 carry haplotypes R1b/M343(×P297) and R1b1/L754(×M269), respectively, which are ancestral for the pre-M269 branch (P297) and the M269 branch. Importantly, we do not observe R1b-Z2103 or immediate R1b-M269 precursor lineages, which originated in the steppe and are later linked with expansion of steppe-related ancestry.
Genetic ancestries during the Bronze Age
The EBA individuals in this study are characterized by two contrasting clusters of genetic ancestry in PCA space (Fig. 1b) and different genetic affinities in outgroup f3 statistics (Fig. 2 and Supplementary Table C). Individuals from YUN and individual PIE078, who date to the first half of the third millennium BC, resemble the SEE CA groups, whereas BOY_EBA and MAJ_EBA individuals fall within the ‘steppe ancestry’ cluster, commonly associated with the Yamnaya cultural complex. Two outlier individuals, BOY019 and YUN041, fall in the space between. Intriguingly, the males from YUN_EBA/PIE078 carried Y-chromosome lineages I2a, suggestive of a HG legacy, while the males from BOY/MAJ_EBA carried R1b-Z2103 or derived lineages, a characteristic hallmark of Yamnaya-associated ancestry (Extended Data Fig. 3b).
On the basis of these observations we tested for additional attraction towards HG-related groups in YUN_EBA and PIE078 compared to their CA predecessors by using f4(CA, EBA; HGs, Mbuti) and confirmed the excess HG ancestry in EBA individuals from YUN and PIE with significant negative results (|Z| ≤ 3) (Extended Data Fig. 7 and Supplementary Table T). By contrast, for MAJ_EBA, BOY_EBA, BOY019 and YUN041, we tested for additional attraction towards farmer-related groups represented by VAR_CA when compared to Yamnaya-associated groups (test) using f4(test, EBA, VAR_CA, Mbuti) (Supplementary Table E). Here, only the outlier individual YUN041 has a higher affinity to VAR_CA than to other EBA groups (Extended Data Fig. 8 and Supplementary Table U). Distal qpAdm modelling with cornerstone populations confirms the contrasting ancestries of the two main EBA clusters. PIE078 and YUN_EBA can be modelled with Turkey_N, CHG and WHG (Fig. 3c and Supplementary Table X), whereas MAJ_EBA, BOY_EBA, BOY019 and YUN041 require EHG ancestry as an additional source (Fig. 3c).
We then explored the apparent homogeneity of Yamnaya-associated EBA steppe pastoralist groups, by testing for possible contribution(s) from four sources: Ukraine Eneolithic as a proxy for mixed Turkey_N/CHG/EHG ancestry, Ukraine N as an HG-related group, Steppe Eneolithic as pre-Yamnaya genetic substrate and Caucasus Eneolithic/Maykop as a proxy for mixed Turkey_N/CHG-related South Caucasus ancestry, as suggested by ref. 47 and directly supported by our results for the preceding Eneolithic period. First, we formally tested for shared drift between all EBA Yamnaya-associated individuals and Steppe Eneolithic/Caucasus Eneolithic/Maykop with respect to cornerstone populations by using f4(Steppe Eneolithic/Caucasus Eneolithic/Maykop, EBA; cornerstones, Mbuti). With the exception of Yamnaya Caucasus, all EBA individuals show an excess affinity to Turkey_N when compared to Steppe Eneolithic (Extended Data Fig. 9 and Supplementary Table V). Further, when compared to Caucasus Eneolithic/Maykop all EBA individuals share drift with WHG and EHG/WSHG and only YUN041 is also significant for Turkey_N (Extended Data Fig. 9 and Supplementary Table V). Second, we used f4-symmetry statistics of the form f4(steppe1, steppe2; test, Mbuti) where test includes Ukraine N, Ukraine Eneolithic, Caucasus Eneolithic/Maykop and Steppe Eneolithic. Here, with the exception of outlier individual Ukraine_Ozera_EBA_Yamnaya, all f4 statistics are non-significant (|Z| ≤ 3) (Supplementary Table W), which indicates that all Yamnaya-associated individuals including those from Ukraine and Bulgaria are genetically highly similar.
Applying the same rationale and sources to proximal qpAdm modelling to uncover subtle signals (Fig. 3f and Supplementary Table Y), we find that BOY_EBA and Yamnaya Samara can be modelled as a three-way mixture of Steppe Eneolithic, Caucasus Eneolithic/Maykop and Ukraine N. We note that the same three sources contributed to the preceding Ukraine Eneolithic individuals from USV and MAJ (in addition to SEE CA ancestry), which suggests that similar processes had led to the tripartite ancestry formation in the steppe zone during the fourth millennium bc. Indeed, we find that BOY_EBA, MAY_EBA and Yamnaya Samara can also be modelled as a two-way mixture of Steppe Eneolithic and KTL001 (who lacked SEE_CA ancestry). For Ukraine_EBA_Yamnaya, we find support for a three-way model (P = 0.07) with Steppe Eneolithic (around 75%), Caucasus Eneolithic/Maykop (around 14%) and Globular Amphora (around 11%) as a western source but also improved model fit (P = 0.5) for a two-way mixture of Steppe Eneolithic (around 65%) and USV (around 35%) (Supplementary Table Y), which suggests a possible direct contribution of Ukraine Eneolithic groups to steppe pastoralists in the third millennium bc. By contrast, Yamnaya Caucasus individuals from the southern steppe can be modelled as a two-way model of around 76% Steppe Eneolithic and 26% Caucasus Eneolithic/Maykop, confirming the findings of Lazaridis and colleagues47. This two-way mix (40% + 60%, respectively) also provides a well-fit model (P = 0.09) for the Ozera outlier individual, consistent with the position in PCA and corroborating an influence from the Caucasus. Despite the overlap in PCA, these results suggest subtle geographical structure, involving local genetic strata and influences from neighbouring groups in western and southern contact zones, respectively. Individual BOY019 can be modelled successfully with around 63% USV and around 37% Steppe Eneolithic ancestry or around 40% Ukraine Trypillia and around 60% Steppe Eneolithic, suggesting interaction between these two neighbouring groups in the western contact zone or alternatively direct descent from admixed groups (for example, KTL001). Finally, individual YUN041 can be modelled as around 50% local YUN_EBA ancestry and 50% of either BOY_EBA or another Yamnaya-associated source.
Discussion
The genetic homogeneity observed in and across the four CA sites (PIE, YUN, PTK and VAR) of the fifth millennium bc matches the cultural homogeneity of the archaeological records and suggests an extended period of a relative stable sociopolitical network and absence of large-scale cultural and genetic transformations. Shared shorter IBD tracts between sites are consistent with the transregional connectivity visible in the material culture. We can only speculate about the reasons that led to decreasing settlement densities at the end of the CA. Conflict arising from an early expansion of supposedly ‘Indo-European’ groups from the steppe, an idea that was put forward by M. Gimbutas18, is possible but internal competition and strife between CA groups is equally likely. In fact, given the near-identical genetic ancestry profiles of SEE CA groups, we caution that genetic analyses would be blind to internal conflicts, causing the replacement of one CA group by another. Long-lasting droughts and forest fires16 or infectious diseases and ensuing epidemics are other factors that could deplete lands. Indeed, evidence for early forms of Yersinia pestis as old as 5,000 years has been reported48,49,50 and even further back in time for Salmonella enterica51 for individuals associated with transitional foraging and pastoralism. Despite the systematic screening of teeth, we found no evidence for pathogens among the CA individuals of the fifth and fourth millennium bc, apart from two individuals (YUN048 and VAR021), who were positive for the Hepatitis B virus (HBV)52, while individual VAR021 was also positive for Salmonella enterica.
A principal finding from our study indicates early contact and admixture between CA farming groups from SEE and Eneolithic groups from the steppe zone in today’s southern Ukraine, possibly starting in the middle of the fifth millennium bc when settlement densities shifted further north, connecting the lower Danube region with the coastal steppe and Cucuteni–Trypillia groups of the forest–steppe. Archaeological evidence shows that the early CA Gumelniţa groups had already settled deep into the steppe zone by the mid-fifth millennium bc, introducing elements of a farming lifestyle but also carrying cultural influences from local HG groups53. The succeeding Cernavodă I and Usatove archaeological cultures were heavily influenced by local CA cultures and surrounds. During the fourth millennium bc, the northwestern Pontic region experienced intensified contact with Steppe Eneolithic groups, while these in turn also had contact with groups in the North Caucasus, such as Maykop, all of which are mirrored by the genomic data presented here. Moreover, despite the close geographical proximity of the Ukrainian sites studied, we were able to trace different admixture histories. Here, the heterogeneity of the individuals from the site Kartal stands out, which is located on the Danube delta at the northern end of the former distribution of the Chalcolithic Gumelniţa–Kodžadermen–Karanovo VI complex and thus represents the transformative nature and dynamics of the fourth millennium bc in action. By contrast, the more homogenous Majaky and Usatove groups, located north of the Dniester River, show that such assimilation processes had already occurred, suggesting that contact and exchange between transitional foragers and early pastoralist groups from the forest–steppe zone and non-local SEE farmer-associated groups had started already in the late fifth millennium bc. Moreover, variable cultural influences attested by the archaeological record40,41,53 are also traceable genetically. We argue that livestock, innovations and technological advances were exchanged through these zones of interaction, which then led to the establishment of fully developed pastoralism in the steppe by the end of the fourth millennium BC. Gene flow from both contact zones into the steppe could also explain the small amounts of farmer-related ancestry in the emerging Yamnaya pastoralists, which differentiates them from the Steppe Eneolithic substrate and accounts for subtle geographical structure in the vastly expanding territory/range.
The early admixture during the Eneolithic presented in this study appears to be local to the northwestern Black Sea region of the fourth millennium bc and did not affect the hinterland in SEE. In fact, EBA individuals from the fourth and third millennia bc from YUN and PIE do not show traces of steppe-like ancestry but instead a resurgence of HG ancestry observed widely in Europe during the fourth millennium bc (refs. 4,29,54,55). This indicates the presence of remnant HG groups in various non-farmed regions, for example, highlands and uplands or densely forested zones and wetlands and a mosaic of ancestries rather than a genetically uniform CA and EBA Europe.
While only a few tell sites have been resettled by local and/or incoming groups who did not originate in the North Pontic region, we can trace the appearance of migrants from the steppe, clearly attributed to Yamnaya culturally and genetically, in the local time transect at Majaky but also at Boyanovo in the Bulgarian lowlands of the Thracian Plain. The subtle differences in genetic ancestries between these two when compared to different Yamnaya-associated groups account for their geographical locations and different stages of genetic and perhaps, cultural assimilation. Two outlier individuals from EBA YUN and BOY bear witness to occasional admixture between inhabitants of EBA tells and incoming steppe pastoralists. Ultimately, the third millennium bc form of ‘steppe’-ancestry is expected to have reached the Great Hungarian plain, from where it diversified and spread further west. The interaction between local and incoming groups in SEE did not result in archaeologically visible conflicts or a near-complete autosomal genetic turnover as observed in Britain or a replacement of the Y-chromosome lineages in the Iberian Peninsula36,56.
Further integrated archaeogenomic studies are needed to disentangle the dynamics at play around the Black Sea during the formative periods of the admixture clines demonstrated in this study. High-quality genome-wide data from the fifth and fourth millennia bc that allow the direct tracing of IBD blocks shared by contributing groups will hold the key to understanding the population history of West Eurasia.
Methods
Permission statement
Permission to work on the archaeological samples was granted by the respective excavators, archaeologist and curators and museum directors of the sites, who are co-authoring the study.
Radiocarbon dating
Of the 135 individuals reported in this study we obtained new direct 14C dates for 113 individuals. Radiocarbon dating was carried out using accelerated mass spectrometry at the Curt-Engelhorn-Zentrum Archäometrie gGmbH in Mannheim, Germany (Fig. 1c and Supplementary Table A). All samples were calibrated on the basis of the IntCal20 database and using OxCal v.4.4.2. All 14C dates in this study are consistent with the archaeological chronology based on stratigraphy and grave goods. We also included 11 published, direct 14C dates for individuals from Varna58,59,60 (Fig. 1c and Supplementary Table A).
Ancient DNA laboratory procedures
Ancient DNA work was carried out in dedicated clean room facilities of the Max Planck Institute for Evolutionary Anthropology (MPI-EVA), Leipzig and Jena, Germany. We processed 168 petrous bones and 129 teeth in total. Petrous bones were sampled with a minimal invasive method61 and, for the sampling of the teeth, the crown was separated from the root and the inner pulp chamber was drilled out62. DNA was extracted from all samples following a modified protocol refs. 63,64. DNA double-stranded libraries were built using a partial uracil-DNA-glycosylase (UDG-half) treatment65. For samples that did not meet the threshold for further analysis, we attempted to increase the DNA yield by using an automated protocol for producing single-stranded, non-UDG libraries66,67. All libraries were double-indexed with a unique pair of indices68.
First, all indexed libraries were screened by means of shotgun sequencing of 5 million reads on an Illumina HiSeq4000 or NextSeq500 sequencing platform using a single end (1 × 75 base pair (bp) reads) kit, followed by an assessment of human DNA content and DNA damage profiles (initial quality criteria). Libraries above the threshold of 0.1% endogenous DNA were enriched for around 1.2 million SNPs in a targeted in-solution capture (1,240,000 SNP capture)31. Enriched libraries were sequenced on HiSeq4000 and NextSeq500 Illumina platforms using a single-read (SR 75) kit and sequencing 40 million reads for libraries between 0.1% and 2% or 20 million reads for libraries above 2%, resulting in a mean coverage of 0.7× (Supplementary Table A). An additional mitochondrial capture4,69 was performed for individuals for which enough reads could not be obtained as by-catch of the 1,240,000 capture, resulting in an average coverage of 64×. For selected male individuals we also performed an inhouse capture assay for the Y chromosome (YMCA)70 which targets around 10.445 kB on the non-combining region of the Y chromosome and which resulted in a mean coverage of 0.08×. Together, a total of 135 individuals yielded sufficient genomic data for downstream analysis.
Sequence data processing
After demultiplexing, EAGER v.1.92.56 (ref. 71) was used to process raw ancient DNA sequence data. Raw reads were trimmed for Illumina adaptor sequences using AdapterRemoval v.2.3.0 (ref. 72). Subsequently, reads were mapped to the human reference genome hs37d5 using BWA v.0.7.12 (ref. 73) and duplicates were removed using DeDup v.0.12.1 (ref. 71). To analyse characteristic DNA damage in the form of G to A and C to T substitutions, mapDamage v.2.0.9 (ref. 74) was used. The effect of postmortem DNA damage on genotyping was minimized by removing 2 bp from the 3′ and 5′ ends of reads from double-stranded UDG-half-treated libraries (n = 131) using the trimbam function included in bamUtils v.1.0.13 (ref. 75). The resulting filtered bam files were genotyped with pileupCaller v.1.4.0.2 (ref. 76) by randomly calling one allele per position considering the human genome as a pseudohaploid genome (–randomHaploid). Only for quality controls 10 bp were removed from the 3′ and the 5′ ends for non-UDG treated single-stranded libraries, whereas the untrimmed bam files were treated with the–singleStrandMode in pileupCaller for genotyping. Coverage statistics calculations and bam filtering were done using samtools (v.1.3; ref. 77).
Ancient DNA authentication
All libraries, except PTK001, yielded damage patterns characteristic of ancient DNA, which includes short DNA fragment lengths (45–65 bp on average) and postmortem deamination at the end of the molecules (6–17% for partial UDG treatment, 30–38% for non-UDG treatment). We merged Shotgun, 1,240,000 and mitochondrial capture data for each individual, mapped this against the revised Cambridge Reference Sequence for the complete human mitochondrial genome (NC 012920.1) and estimated contamination for both sexes on the mitochondrium using ContamMix78 (Supplementary Table A), ranging from 0.086% to −9.2%. The nuclear contamination for males was estimated using ANGSD79 and ranged from 0.2% to 2%. PTK001 yielded a contamination estimate of around 18% and therefore was excluded from all further analysis. We estimated the genetic sex by calculating the coverage on the X, Y and the autosomal chromosomes, for which the X and Y coverage is normalized by the autosomal coverage and the relative length of each sex chromosome80.
DNA reference datasets
The new genotype data were restricted to two sets of reference panels, the Affymetrix Axiom Genome-wide Human Origins1 array (HO; 593,124 autosomal SNPs)2,81 and the 1,240,000 panel (1.233,013 autosomal SNPs including all of the HO SNPs)31. The number of SNPs covered at least once for each of these reference panels is given in Supplementary Table A.
Genetic relatedness analysis
Genetic relatedness was estimated using READ82, using default parameter settings. Background relatedness was estimated using the median value, across all sites per temporal group (Supplementary Information 7). From pairs of first-degree relatives, the individual with lower number of SNPs on the 1,240,000 target region was excluded from downstream analysis. Three individuals from PIE were identified as identical and were therefore merged for downstream analysis. Two pairs of the newly published samples from YUN CA had to be merged as they were revealed to be the same individuals. One individual from VAR and one from YUN were merged with previously published individuals from each site because they were sampled from the same individual and therefore identical29 (Supplementary Table I).
Assignment of uniparentally inherited haplogroups
Trimmed Shotgun, 1,240,000 and mitochondrial capture reads were aligned to the revised Cambridge Reference Sequence for the complete human mitochondrial genome (NC 012920.1) and a consensus sequence for each individual was retrieved using Geneious v.2019.2.3 (ref. 83). HaploGrep2 (v.2.4.0; ref. 84) was used to assign each consensus sequence to a specific mitochondrial haplogroup (Supplementary Table A). Y-chromosome haplogroups for all male individuals were assigned using the manual assignment method of Y-haplogroup calling as described in ref. 70 (Supplementary Table A). In the case of non-UDG treated sequence, YMCA data were filtered to exclude C to T and G to A transitions on the forward and reverse strands, respectively.
Population genetic analysis
For genome-wide analyses the new data from this study were merged with published ancient and modern data from the Allen Ancient DNA Resource (AADR) v.44.3 (https://reich.hms.harvard.edu/allen-ancient-dna-resource-aadr-downloadable-genotypespresent-day-and-ancient-dna-data). Data on the HO panel (around 600,000 SNPs) were used for PCA using the program ‘smartpca’ v.16000 (EIGENSOFT85). Principal components were computed for 1,253 present-day western Eurasians from 77 different populations (Supplementary Table B) on which ancient individuals were projected, using the options ‘lsqproject: YES’ and ‘shrinkmode: YES’. Individuals with fewer than 30,000 SNPs on the HO-dataset covered were excluded from the PCA. All other analyses were performed on the above merged dataset on the 1,240,000 SNP panel (around 1.24 million SNPs). Outgroup f3 statistics86 were calculated using qp3Pop to obtain the genetic relatedness of a target population to a set of ancient Eurasian populations since the divergence from an African outgroup. The f4 and f3 statistics were calculated using qpDstat and the f4mode: YES function. Standard errors were computed with the default block jackknife approach and 3 s.e. are reported and plotted. The f3 and f4 statistics were calculated using the ADMIXTOOLS81 package.
Genetic admixture modelling
Ancestry modelling and ancestry proportion estimation on the 1,240,000 SNP dataset was performed using qpAdm in ADMIXTOOLS (v.5.1; ref. 4). The following groups were used as a basic set of outgroups for distal modelling: Mbuti.DG, Turkey_Epipaleolithic, Iran_GanjDareh_N, Russia_MA1_HG.SG, Russia_Kostenki14, Italy_North_Villabruna_HG. Depending on the time period, the outgroup set was adjusted according to the specific test. A detailed list of outgroups per test can be found in Supplementary Tables G, H, R, S, X and Y.
Admixture date estimation with DATES for PIE060
The software DATES (v.753)33 was used to estimate the time of the admixture events of ancient populations under the assumption that gene flow occurred as a single event and that the generation time is 28 years34. DATES measures the decay of ancestry covariance to infer the admixture time and estimates the variance of this admixture using a jackknife approach. The following parameters were used for every run: binsize 0.001; maxdis 1; qbin 10; lovalfit 0.45. For PIE060, the two reference populations were chosen on the basis of the best-fitting ancestry model from qpAdm.
Imputation
Samples were imputed using GLIMPSE (v.1.0.1) with the default parameters87,88. Briefly, bam files were trimmed 2 bp to remove ancient DNA damage. We then determined genotype likelihoods from trimmed bam files using bcftools89 with the 1,000G panel (The 1,000 Genomes Project consortium90) as a reference. We used GLIMPSE_impute on genomic chunks of 2,000,000 bp with the buffer size of 200,000 bp to perform imputation. We then ligated the chunks using GLIMPSE_ligate and determined the most likely haplotypes using GLIMPSE_sample. Samples with more than 0.5× coverage on the 1,240,000 positions (around 550,000 SNPs) after imputation were included in IBD analysis. No MAF filtering was performed, since only 1,240,000 positions were retained after imputation.
Runs of homozygosity
The software package HapROH (v.0.64) was used to analyse ROH on pseudohaploid 1,240,000 SNP capture data91. Only samples with more than 400,000 SNPs were included in the analysis to prevent potential false positives (Supplementary Table K).
IBD sharing
IBD sharing analysis was done using ancIBD (v.0.4)92 on individuals with more than 600,000 SNPs and genotype probabilities > 0.99 after imputation with GLIMPSE87,88. We used HapBLOCK to perform the IBD sharing estimation. Imputed samples were merged, then the vcf_to_1240K_hdf command was used to convert the vcf files to the hdf5 format. The hapBLOCK_chroms command was used to perform the IBD sharing analysis for each chromosome at a time using the default parameters. Following that, only shared blocks of more than 220 SNPs per centimorgan and shared blocks of more than 5 cM were kept for data quality purposes and used for plotting (Supplementary Table J).
Metagenomic pathogen screening
Shotgun sequencing data were screened for the presence of pathogen DNA with the screening pipeline HOPS (v.0.2)93. First, adaptor-clipped reads were mapped to a custom-made RefSeq database using MALT v.0.4.0 (ref. 94) in BlastN mode and with semiglobal alignment type and default pipeline settings. The used database included all available complete bacterial and viral genomes as of 2017 in addition to selected eukaryotic pathogen genomes and the human reference sequence GRCh38. The results were filtered with a predefined list of pathogens of interest and possible candidates authenticated on the basis of edit distance distribution, ancient DNA damage pattern and read distribution along the reference genome.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The DNA sequences reported in this paper have been deposited in the European Nucleotide Archive under the accession number PRJEB62503.
References
Lazaridis, I. et al. Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature 513, 409–413 (2014).
Lazaridis, I. et al. Genomic insights into the origin of farming in the ancient Near East. Nature 536, 419–424 (2016).
Hofmanová, Z. et al. Early farmers from across Europe directly descended from Neolithic Aegeans. Proc. Natl Acad. Sci. USA 113, 6886–6891 (2016).
Haak, W. et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522, 207–211 (2015).
Allentoft, M. E. et al. Population genomics of Bronze Age Eurasia. Nature 522, 167–172 (2015).
Sherratt, A. in Pattern of the Past: Studies in the Honour of David Clarke (eds Hodder, I. et al.) 261–306 (Cambridge Univ. Press, 1981).
Hansen, S. in Appropriating Innovations: Entangled Knowledge in Eurasia, 5000–1500 BC (eds Stockhammer, P. & Maran, J.) 136–148 (Oxbow Books, 2017).
Librado, P. et al. The origins and spread of domestic horses from the Western Eurasian steppes. Nature 598, 634–640 (2021).
Scott, A. et al. Emergence and intensification of dairying in the Caucasus and Eurasian steppes. Nat. Ecol. Evol. 6, 813–822 (2022).
Renfrew, C. & Appadurai, A. in The Social Life of Things: Commodities in Cultural Perspective (ed. Appadurai, A.) 141–168 (Cambridge Univ. Press, 1986).
Stockhammer, P. in Rich and Poor—Competing for Resources in Prehistoric Societies (eds Meller, H. et al.) 77–84 (Ludwig-Maximilians-Universität München, 2016).
Hansen, S. in Surplus without the State—Political Forms in Prehistory (eds Meller, H. et al.) 221–246 (Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt—Landesmuseum für Vorgeschichte, 2018).
Demoule, J.-P. in La Révolution Néolithique en France 78–89 (La Découverte, 2007).
Hansen, S. et al. Pietrele am “Lacul Gorgana”: Bericht über die Ausgrabungen in der neolithischen und kupferzeitlichen Siedlung und die geomorphologischen Untersuchungen in den Sommern 2012–2016. Eurasia Antiqua 20, 1–116 (2014).
Boyadzhiev, Y., Boyadzhiev, K., Brandtstätter, L. & Krauß, R. Chronological modelling of the Chalcolithic settlement layers at Tell Yunatsite, Southern Bulgaria. Doc. Praehist. 48, 252–275 (2021).
Todorova, H. Die überregionalen komplexen Gemeinschaften, Kulturblöcke und ökologischen Krisen in der Urgeschichte im Raum der unteren Donau (VII-I Jahrtausend v. Chr.). Studia Praehist. 14, 143–154 (2011).
Boyadzhiev, K. in Southeast Europe and Anatolia in Prehistory—Essays in Honor of Vassil Nokilov on his 65th Anniversary (ed. Krum Bacvarov, R. G.) 261–268 (Verlag Dr. Rudolf Habelt GmbH, 2016).
Gimbutas, M. A. The Civilization of the Goddess: The World of Old Europe (Harper San Francisco, 1991).
Müller, J., Rassmann, K. & Videiko, M. Trypillia Mega-Sites and European Prehistory: 4100–3400 BC (Routledge, 2016).
Gaydarska, B. Early Urbanism in Europe: The Trypillia Megasites of the Ukrainian Forest-Steppe (De Gruyter, 2020).
Hansen, S. Arsenic Bronze: an archaeological introduction into a key innovation. Eurasia Antiqua 23, 139–162 (2021).
Klimscha, F. Transforming technical know-how in time and space. Using the digital atlas of innovations to understand the innovation process of animal traction and the wheel. eTopoi J. Ancient Stud. 6, 16–63 (2017).
Piotrovsky, Y. Y. The Maikop kurgan (Oshad): a modern view. Camera Praehist. 1, 61–75 (2020).
Frînculeasa, A. Earthen burial mounds and the Coţofeni Culture south of the Carpathians. The archaeological research in Ariceștii-Rahtivani—Movila pe Răzoare. Ziridava. Studia Archaeologica 34, 35–90 (2020).
Manzura, I. in Repräsentationen der Macht (ed. Hansen, S.) 73–96 (Kolloquien zur Vor- und Frühgeschichte 25, 2020).
Manzura, B. & Govedarica, I. in Der Schwarzmeerraum vom Äneolithikum bis in die Früheisenzeit [5000-500 v.Chr.]: Band 2: Globale Entwicklung versus Lokalgeschehen (eds Sava, E. et al.) 41–61 (Verlag Marie Leidorf, 2011).
Racimo, F., Sikora, M., Vander Linden, M., Schroeder, H. & Lalueza-Fox, C. Beyond broad strokes: sociocultural insights from the study of ancient genomes. Nat. Rev. Genet. 21, 355–366 (2020).
Alexandrov, S. in Gold & Bronze. Metals, Technologies and Interregional Contacts in the Eastern Balkans during the Bronze Age (eds Alexandrov, S. et al.) 85–96 (National Archaeological Institute and Museum, 2018).
Mathieson, I. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018).
Alexandrov, S. & Kaiser, E. in Der Schwarzmeerraum vom Neolithikum bis in die Früheisenzeit (6000-600 v. Chr.) (eds Nikolov, V. & Schier, W.) 359–370 (Verlag Marie Leidorf, 2016).
Mathieson, I. et al. Genome-wide patterns of selection in 230 ancient Eurasians. Nature 528, 499–503 (2015).
Jones, E. R. et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians. Nat. Commun. 6, 8912 (2015).
Chintalapati, M., Patterson, N. & Moorjani, P. Reconstructing the spatiotemporal patterns of admixture during the European Holocene using a novel genomic dating method. eLife 11, e77625 (2022).
Fenner, J. N. Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies. Am. J. Phys. Anthropol. 128, 415–423 (2005).
Kivisild, T. The study of human Y chromosome variation through ancient DNA. Hum. Genet. 136, 529–546 (2017).
Olalde, I. et al. The genomic history of the Iberian Peninsula over the past 8000 years. Science 363, 1230–1234 (2019).
Brunel, S. et al. Ancient genomes from present-day France unveil 7,000 years of its demographic history. Proc. Natl Acad. Sci. USA 117, 12791–12798 (2020).
Brace, S. et al. Ancient genomes indicate population replacement in Early Neolithic Britain. Nat. Ecol. Evol. 3, 765–771 (2019).
Wang, C.-C. et al. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions. Nat. Commun. 10, 590 (2019).
Rassamakin, Y. in Late Prehistoric Exploitation of the Eurasian Steppe (eds Levine, M. et al.) 59–182 (McDonald Institute Monographs, 1999).
Rassamakin, J. Die nordpontische Steppe in der Kupferzeit: Gräber aus der Mitte des 5. Jts. bis Ende des 4. Jts. v. Chr. (Philipp von Zabern, 2004).
Anthony, D. in The Black Sea Flood Question: Changes in Coastline, Climate, and Human Settlement (eds Yanko-Hombach, V. et al.) 345–370 (Springer, 2007).
Petrenko, V., Gerling, C. & Kaiser, E. Majaki—Ein komplexes Denkmal der Usatovo-Kultur. Eurasia Antiqua 21, 45–74 (2015).
Wood, R. E. et al. Freshwater radiocarbon reservoir effects at the burial ground of Minino, Northwest Russia. Radiocarbon 55, 163–177 (2013).
Anthony, D. W. et al. The Eneolithic cemetery at Khvalynsk on the Volga River. Praehist. Zeitschr. 97, 22–67 (2022).
Immel, A. et al. Gene-flow from steppe individuals into Cucuteni-Trypillia associated populations indicates long-standing contacts and gradual admixture. Sci. Rep. 10, 4253 (2020).
Lazaridis, I. et al. The genetic history of the Southern Arc: a bridge between West Asia and Europe. Science 377, eabm4247 (2022).
Rascovan, N. et al. Emergence and spread of basal lineages of Yersinia pestis during the Neolithic Decline. Cell 176, 295–305 (2019).
Susat, J. et al. A 5,000-year-old hunter-gatherer already plagued by Yersinia pestis. Cell Rep. 35, 109278 (2021).
Andrades Valtueña, A. et al. Stone Age Yersinia pestis genomes shed light on the early evolution, diversity, and ecology of plague. Proc. Natl Acad. Sci. USA 119, e2116722119 (2022).
Key, F. M. et al. Emergence of human-adapted Salmonella enterica is linked to the Neolithization process. Nat. Ecol. Evol. 4, 324–333 (2020).
Kocher, A. et al. Ten millennia of hepatitis B virus evolution. Science 374, 182–188 (2021).
Reingruber, A. & Rassamakin, J. in Der Schwarzmeerraum vom Neolithikum bis in die Früheisenzeit (6000–600 v. Chr.) (eds Nikolov, V. & Schier, W.) 273–310 (Verlag Marie Leidorf, 2016).
Lipson, M. et al. Parallel palaeogenomic transects reveal complex genetic history of early European farmers. Nature 551, 368–372 (2017).
Papac, L. et al. Dynamic changes in genomic and social structures in third millennium bc central Europe. Sci. Adv. 7, eabi6941 (2021).
Olalde, I. et al. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature 555, 190–196 (2018).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing https://www.R-project.org/ (2022).
Higham, T. et al. New perspectives on the Varna cemetery (Bulgaria)–AMS dates and social implications. Antiquity 81, 640–654 (2007).
Krauß, R. et al. Chronology and development of the Chalcolithic necropolis of Varna I. Doc. Praehist. 44, 282–305 (2017).
Higham, T., Slavchev, V., Gaydarska, B. & Chapman, J. AMS dating of the Late Copper Age Varna Cemetery, Bulgaria. Radiocarbon 60, 493–516 (2018).
Orfanou, E., Himmel, M., Aron, F. & Haak, W. Minimally-invasive sampling of pars petrosa (os temporale) for ancient DNA extraction V.2. protocols.io https://doi.org/10.17504/protocols.io.bqd8ms9w (2020).
Neumann, G. U., Andrades Valtuena, A., Fellow Yates, J. A., Stahl, R. & Brandt, G. Tooth sampling from the inner pulp chamber for ancient DNA Extraction V.2. protocols.io https://doi.org/10.17504/protocols.io.bqebmtan (2020).
Dabney, J. et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl Acad. Sci. USA 110, 15758–15763 (2013).
Velsko, I., Skourtanioti, E. & Brandt, G. Ancient DNA extraction from skeletal material. protocols.io https://doi.org/10.17504/protocols.io.baksicwe (2020).
Aron, F., Neumann, G. U. & Brandt, G. Half-UDG treated double-stranded ancient DNA library preparation for Illumina sequencing V.1. protocols.io https://doi.org/10.17504/protocols.io.bmh6k39e (2020).
Gansauge, M.-T. & Meyer, M. Single-stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nat. Protoc. 8, 737–748 (2013).
Gansauge, M.-T., Aximu-Petri, A., Nagel, S. & Meyer, M. Manual and automated preparation of single-stranded DNA libraries for the sequencing of DNA from ancient biological remains and other sources of highly degraded DNA. Nat. Protoc. 15, 2279–2300 (2020).
Stahl, R. et al. Illumina double-stranded DNA dual indexing for ancient DNA V.2. protocols.io https://doi.org/10.17504/protocols.io.bvt8n6rw (2021).
Maricic, T., Whitten, M. & Pääbo, S. Multiplexed DNA sequence capture of mitochondrial genomes using PCR products. PLoS ONE 5, e14004 (2010).
Rohrlach, A. B. et al. Using Y-chromosome capture enrichment to resolve haplogroup H2 shows new evidence for a two-path Neolithic expansion to Western Europe. Sci. Rep. 11, 15005 (2021).
Peltzer, A. et al. EAGER: efficient ancient genome reconstruction. Genome Biol. 17, 60 (2016).
Schubert, M., Lindgreen, S. & Orlando, L. AdapterRemoval v2: rapid adapter trimming, identification, and read merging. BMC Res. Notes 9, 88 (2016).
Li, H. & Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 26, 589–595 (2010).
Jónsson, H., Ginolhac, A., Schubert, M., Johnson, P. L. F. & Orlando, L. mapDamage2.0: fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 29, 1682–1684 (2013).
Jun, G., Wing, M. K., Abecasis, G. R. & Kang, H. M. An efficient and scalable analysis framework for variant extraction and refinement from population-scale DNA sequence data. Genome Res. 25, 918–925 (2015).
Schiffels, S. sequenceTools. GitHub https://github.com/stschiff/sequenceTools (2019).
Li, H. et al. 1000 Genome Project Data Processing Subgroup, The Sequence Alignment/ Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Fu, Q. et al. A revised timescale for human evolution based on ancient mitochondrial genomes. Curr. Biol. 23, 553–559 (2013).
Korneliussen, T. S., Albrechtsen, A. & Nielsen, R. ANGSD: analysis of next generation sequencing data. BMC Bioinformatics 15, 356 (2014).
Mittnik, A., Wang, C.-C., Svoboda, J. & Krause, J. A molecular approach to the sexing of the triple burial at the Upper Paleolithic Site of Dolní Věstonice. PLoS ONE 11, e0163019 (2016).
Patterson, N. et al. Ancient admixture in human history. Genetics 192, 1065–1093 (2012).
Monroy Kuhn, J. M., Jakobsson, M. & Günther, T. Estimating genetic kin relationships in prehistoric populations. PLoS ONE 13, e0195491 (2018).
Kearse, M. et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649 (2012).
Weissensteiner, H. et al. HaploGrep 2: mitochondrial haplogroup classification in the era of high-throughput sequencing. Nucleic Acids Res. 44, W58–63 (2016).
Patterson, N., Price, A. L. & Reich, D. Population structure and eigenanalysis. PLoS Genet. 2, e190 (2006).
Raghavan, M. et al. Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans. Nature 505, 87–91 (2014).
Delaneau, O. GLIMPSE. github.io https://odelaneau.github.io/GLIMPSE/tutorial_b38.html (2020).
Rubinacci, S., Ribeiro, D. M., Hofmeister, R. J. & Delaneau, O. Efficient phasing and imputation of low-coverage sequencing data using large reference panels. Nat. Genet. 53, 120–126 (2021).
Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 27, 2987–2993 (2011).
1000 Genomes Project Consortium. et al. A global reference for human genetic variation. Nature 526, 68–74 (2015).
Ringbauer, H., Novembre, J. & Steinrücken, M. Parental relatedness through time revealed by runs of homozygosity in ancient DNA. Nat. Commun. 12, 5425 (2021).
Ringbauer, H. et al. ancIBD—Screening for identity by descent segments in human ancient DNA. Preprint at bioRxiv https://doi.org/10.1101/2023.03.08.531671 (2023).
Hübler, R. et al. HOPS: automated detection and authentication of pathogen DNA in archaeological remains. Genome Biol. 20, 280 (2019).
Vågene, Å. J. et al. Salmonella enterica genomes from victims of a major sixteenth-century epidemic in Mexico. Nat. Ecol. Evol. 2, 520–528 (2018).
Acknowledgements
We thank E. Kukral, F. Aron, L. Semerau, A. Wissgott, G. Brandt, S. Nagel and E. Essel for their support in the ancient DNA analyses and S. Clayton and K. Prüfer for the processing of the raw sequence data. We thank the teams at (former) MPI-SHH-Archaeogenetics and MPI-EVA-Archaeogenetics for continued support and discussion. This research was funded by the Max Planck Society, the European Research Council under the European Union’s Horizon 2020 research and innovation programme (grant agreement nos 771234-PALEoRIDER, W.H.; 834616-ARCHCAUCASUS, S.H.; and 678901-FoodTransforms, P.W.S.) and the ERA.Net RUS Plus initiative (S&T-277-BIOARCCAUCASUS, S.H.).
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Contributions
W.H., S.H., P.W.S. and J.K. designed the study. R.K., G.G.-R., C.S., M.A.S., G.U.N., A. Chohadzhiev, A.S., K.P., S.R., M.L., I.M., B.G., K. Boyadzhiev, Y.B., K. Beutler and I.B. provided materials and resources. S.P. performed laboratory experiments. S.P., A.B.R., G.G.-R., C.S., G.U.N. and A. Childebayeva analysed the data. W.H., I.M., S.H., P.W.S., Y.B. and K. Boyadzhiev assisted with data interpretation. S.P., S.H., P.W.S., I.M., A.B.R. and W.H. wrote the manuscript with contributions from all co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Mehmet Somel and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Tell settlements and burial sites in Southeastern Europe.
a, Aerial view of Tell Măgura Gorgana near Pietrele, Romania (© Konstantin Scheele, German Archaeological Institute, Eurasia Department). b, Detailed view of the 11m stratigraphy at Pietrele (© Svend Hansen, German Archaeological Institute, Eurasia Department). c, Aerial view of Tell Yunatsite, Bulgaria (© Kamen Boyadzhiev). d, Map of the site Orlovka-Kartal, Ukraine. The base map was sourced from Google Earth https://www.google.com/earth/index.html. e, Characteristic finds from the Eneolithic type-site Usatove. f, Characteristic finds attributed to the Cernavoda I phase. g, Burial in flexed position from grave 10 at Kartal (© Igor Bruyako). h, Infant urn-burial from the Early Bronze Age layer south of tell Yunatsite (© Kamen Boyadzhiev).
Extended Data Fig. 2 F4-statistics for CA groups to determine Neolithic proxies.
F4-statistics show different attraction of the CA to Neolithic groups conditioned on HG groups. Z-scores outside the threshold of (|Z|≥1) are highlighted in orange, f4-values are shown with one standard error. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach.
Extended Data Fig. 3 DATES and Y-chromosomal haplogroup diversity.
a, DATES decay curve for the CA outlier individual PIE060 with SEE N and Iron Gates HGs as source populations. b, Changes of Y-chromosomal haplogroup diversity (colour fills) over time (mean 14C dates cal. BC; x-axis) with respect to changes in autosomal ancestry as reflected in PC2 (y-axis), based on the relative density of female (open circles) and male (colour filled squares) of all newly reported individuals in this study.
Extended Data Fig. 4 Identity-by-descent within and between sites.
a, Results of identity-by-descent (IBD) analysis per pair of individuals. Plotting the sum versus the number of the shared chunks of IBD in window sizes of >12cM resolves degrees of biological relatedness up to the 4-6th degree. Within (left) and between site (right) relationships are highlighted separately. b, Stacked bar plot showing the number (left) and the cumulative distribution of the sum (right) of IBD blocks that are shared between all individuals within and between sites.
Extended Data Fig. 5 Cumulative distribution of the runs of homozygosity tracts of all newly reported individuals.
Runs of homozygosity were estimated with hapROH. Individuals are grouped in relative chronological order from right to left. Expected parental relationship and simulated effective populations sizes are given.
Extended Data Fig. 6 F4 statistics for Ukraine Eneolithic groups to determine attraction to ‘cornerstone’ populations.
F4 statistics show different attractions of Ukraine Eneolithic groups to ‘cornerstone’ ancestry groups conditioned on a, Maykop-associated groups, b, Steppe Eneolithic, and c, SEE CA. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach.
Extended Data Fig. 7 F4 statistics for EBA groups to test for excess HG attraction.
F4 statistics show different attractions of EBA groups to HG groups conditioned on their respective preceding or contemporaneous group. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach.
Extended Data Fig. 8 Testing for Anatolian farmer-related affinity in EBA individuals.
F4 statistics show different attractions of the EBA groups to VAR_CA conditioned on Yamnaya-associated groups. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach.
Extended Data Fig. 9 F4 statistics to explore excess affinity of EBA ‘steppe ancestry’ groups conditioned on preceding Eneolithic groups from the steppe and the Caucasus.
F4 statistics show different attractions of the EBA groups to ‘cornerstone’ populations conditioned on pre-Yamnaya groups, Steppe Eneolithic and Caucasus Eneolithic/Maykop. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. EBA groups are given on the y-axis and test populations are given on the x-axis. Standard errors (SE) were computed with the default block jackknife approach.
Supplementary information
Supplementary Information
This file contains Supplementary Text 1–7, Fig. 1 and additional references.
Supplementary Data
This file contains legends for Supplementary Tables A–Y.
Supplementary Tables
This file contains Supplementary Tables A–Y (see Supplementary Data for legends).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Penske, S., Rohrlach, A.B., Childebayeva, A. et al. Early contact between late farming and pastoralist societies in southeastern Europe. Nature 620, 358–365 (2023). https://doi.org/10.1038/s41586-023-06334-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-023-06334-8
This article is cited by
-
Population genomics of post-glacial western Eurasia
Nature (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
