
Abstract
Bone destruction in chronic gout is correlated with deposits of monosodium urate (MSU) crystals. Bone with MSU tophi were histopathologically shown to have altered remodeling and cellular distribution. We investigated the impact of neutrophils in bone remodeling associated with MSU and demonstrated that neutrophils, through elastase localized at their surface, induced retraction of confluent osteoblasts (OBs) previously layered on calcified matrix. This OB retraction allowed osteoclasts to resorb cell-free areas of the matrix. This neutrophil effect was concentration dependent and time dependent and required direct contact with OBs. Exposure of OBs to MSU greatly promoted neutrophil adherence to OBs. Neutrophil membrane at the contact zone with OBs showed concentrated fluorescence of dye PKH-67, indicating a cellular contact. Neutrophil–OB interaction increased the survival of neutrophils, reduced their release of lactoferrin in presence of MSU and did not change OB-mediated mineralization. The adhesion of neutrophils to OBs was heterotypic through neutrophil CD29/CD49d and OB-fibronectin peptide CS1. Leukotriene B4 (LTB4) and platelet-activating factor (PAF) were also involved in neutrophil adherence to OBs, as shown by the blocking effect of selective LTB4 and PAF receptor antagonists, and a cytosolic phospholipase A2α (cPLA2α) inhibitor. Blockade of CD49d/CS1 and inhibition of the cPLA2α had subadditive effects, reducing by 60% the adherence of neutrophils to OBs. Taken together, these data showed that neutrophil adhesion to MSU-activated OBs was mediated by the β1 integrin CD29/CD49d-fibronectin peptide CS1 receptors and cPLA2α-derived metabolites and impacts on OB and osteoclast functions. These interactions could be involved in the local bone remodeling process of gout.
Similar content being viewed by others
Main
Tissue changes of chronic gouty arthritis have been ascribed to deposition of monosodium urate (MSU) microcrystals and inflammation. The hallmark of chronic gout are intraarticular and periarticular tophi. Urate deposits in bone and intracortical bone erosions during gout have been radiologically shown.1 These bone erosions have been correlated with intraosseous tophi by using computed tomography.2 Mechanisms of the altered bone remodeling process responsible for these lesions remain largely unknown. Histopathologically, MSU gives two types of lesions of uratic foci (1) some from adjacent joints are surrounded by an inflammatory tissue and (2) others are found without inflammatory tissue and are ‘located in the immediate subchondral bone and even those present deeper in the spongiosa undoubtedly form there, resulting from the direct deposition of urates in the bone marrow’, according to Jaffe.3 These pathological features suggest direct contact between MSU and bone cells.
Fibroblasts, that originate from stromal cells, were present in the inflammatory tissue surrounding MSU crystal deposition in tophi.3 Fibroblasts were shown to be activated by MSU.4 MSU also directly activated stromal cells which, in turn, promote osteoclastogenesis.5 Moreover, osteoblasts (OBs), that originate from stromal cells, undergo functional deregulation by MSU with reduced parameters of bone formation and increased generation of inflammatory mediators like PGE2, interleukin (IL)-6, and IL-8.6 Indeed, stromal cells and OBs are the major source of IL-6 and IL-8.7 IL-8 is a chemokine that attracts and activates neutrophils, the major cell type involved in gouty arthritis.8, 9 Neutrophils and their precursors are abundantly present in bone marrow. Interestingly, bone marrow neutrophilia induced by IL-6 was shown to be associated with important histological modifications of bone like low number of OBs and reduction of bone turnover.10, 11 In gouty tophi with chronic deposits of MSU in bone structures where bone cells can be directly exposed to MSU, mediators like IL-6 and IL-8 can be locally generated and may affect neutrophils. It is also noteworthy that normal and inflammatory neutrophils can adhere to bone cells.12 Since stromal cells are greatly activated by MSU, it seems possible that neutrophils locally interact with bone cells like OBs during chronic gout. Of relevance to this hypothesis, neutrophils express β2 integrins (CD11a–d/CD18) involved in their adhesion and migration, and the β1 integrin CD29/CD49d (also known as β1/α4) in pathophysiological conditions.13, 14, 15 OBs also express integrins with potential counter-receptors for neutrophil integrins.16 The nature of the integrins implicated in the adhesion of neutrophils to human OBs is presently unknown. Neutrophil adhesion to cells also implicates inflammatory bioactive lipids like platelet-activating factor (PAF) and leukotriene B4 (LTB4).17, 18 In this study, we investigated the impact of neutrophils in bone remodeling driven by MSU.
We first demonstrated that OB morphology was altered by neutrophils, which strongly adhered to MSU-activated OBs. In the presence of MSU-activated OBs, neutrophils showed increased survival and reduction of lactoferrin (LF) release. These direct OB–neutrophil interactions required the adhesion molecules CD49d/fibronectin peptide CS1, as well as LTB4 and PAF. We then carried out histological analyses of gouty tophi in bone that confirmed the presence of MSU deposition and neutrophils in bone structures with an altered morphology of adjacent OBs.
MATERIALS AND METHODS
Reagents
Cycloheximide, SB203580, PD98069, alizarin red S (ARS) and the cell linker PKH-67 were purchased from Sigma Chemical (St Louis, MO, USA). Calcein-AM, propidium iodide (PI), and CellTracker Orange CMTMR were from Invitrogen Canada (Burlington, ON, Canada). Receptor-associated protein (RAP), β-glycerophosphate, human neutrophil elastase, and neutrophil elastase inhibitor II (MSACK) were obtained from Calbiochem (San Diego, CA, USA). CS1 peptide was provided by American Peptide Company (Sunnyvale, CA, USA). Pyrrophenone, BN52021, CP105696 were respectively from Shionogi (Osaka, Japan), Beaufour (Paris, France), and Pfizer (Groton, USA). PAF and LTB4 were purchased from Cayman Chemical (Ann Arbor, MI, USA). Antibodies (Ab) were from different sources: blocking rat anti-CD49d IgG2b Ab (clone R1–2; Cedarlane, Hornby, ON, Canada), blocking anti-MadCAM-1 IgG1 mouse monoclonal Ab (mAb) (clone CA102.2C; Bender MedSystems, Vienna, Austria), anti-VCAM-1 IgG1 mAb (clone 1.G11B1; Chemicon, Billerica, MA, USA), anti-CD54-phycoerythrin (PE) IgG1 mAb (clone HA58; eBioscience, San Diego, CA, USA), anti-CD49d-FITC IgG1, anti-CD29-FITC IgG1, anti-CD49e-PE and anti-CD29-FITC IgG1 mAbs (R&D Systems, Minneapolis, MN, USA), and anti-elastase IgG1 mAb (clone NP57; Dako, Denmark).
Cell Preparation
Volunteers signed a consent form in accordance with the Declaration of Helsinki, and the institutional review board of the Université Laval approved the study. Normal human OBs were grown from trabecular bone explants until confluence before they were recovered and plated at starting densities of 0.5–1 × 106 cells/well in αMEM+10% FBS with antibiotics.6 All incubations were performed at the first passage and 80–90% cell confluence in six-well plates (37°C, humidified atmosphere, 5% CO2). Confluent OBs were preactivated with 1 mg/ml sterile pyrogen-free triclinic MSU (provided by Dr R De Médicis, University of Sherbrooke; mean size: 10 × 1.25 μm, as determined by scanning electron microscopy) for 1 h at 37°C, and washed twice to eliminate non-adherent MSU. Human neutrophils were obtained from peripheral blood of healthy volunteers, as previously described.19 Neutrophils were preincubated at 37°C 30 min with 1 μg/ml calcein-AM. OBs coincubated with labeled neutrophils (short-term neutrophil adherence with an optimal neutrophil/OB ratio of 20/1) were vigorously washed two times to eliminate non-adherent cells. Since a 1-h contact of MSU with OBs was associated with partly engulfed crystals that could affect neutrophils,6 amounts of MSU present in OBs and at their surface after washes were measured and averaged 0.52±0.03 mg/ml (n=6). This latter concentration (half of MSU initially added to OBs) was used to stimulate neutrophils where appropriate. OB retraction (and its reversal) was assessed by using confluent OBs coincubated with neutrophils (neutrophil/OB: 6/1) that were preincubated with elastase inhibitor (100 μM) or with vehicle for 30 min, or by using confluent OBs incubated with purified neutrophil elastase rather than neutrophils. After 48 h of coincubation and two vigorous washes, cells were analyzed by microscopy. OBs or neutrophils were preincubated 30 min with 5 μg/ml blocking anti-CD49d Ab or control rat IgG2b, 10 μg/ml of anti-VCAM-1 or anti-MadCAM-1 mAbs or control mouse IgG1, 150 μg/ml of CS1 peptide, or 5 min with vehicle or 300 nM pyrrophenone (cPLA2α inhibitor),20 1 μM BN52021 (PAF receptor antagonist), 1 μM CP105696 (LTB4 receptor antagonist), 500 nM NS-398 (COX-2 inhibitor), 1 μM indomethacin (COX-1/2 inhibitor), 5 μM SB203580 (p38 inhibitor) or 8 μM PD98069 (ERK inhibitor). All inhibitors and antagonists were used at concentrations optimal for their specific effects in incubation medium containing 10% FBS, and no toxicity (PI exclusion test) on cells was observed as assessed by cytofluorometry.
Microscopy
Five sections of surface areas of each well were studied by microscopy (magnification × 100) and photographed to count adherent fluorescent neutrophils and to calculate percentages of total adherent neutrophils from the whole population of neutrophils added to OBs. Count analysis was performed with the ImagePro software (Media Cybernetics, Bethesda, MD, USA). Neutrophil effects on OB morphology were observed with an Olympus research inverted microscope CKX41. Count analysis and determination of cell-free area were performed with the ImagePro software. Confocal microscopy assessed the contact zones of neutrophils adherent to OBs. Confluent OBs stained with 2 μM CMTMR (30 min, 37°C) were fixed with 4% paraformaldehyde. Neutrophils, previously stained with 4 μM PKH-67 (2 min, room temperature), were added to confluent MSU-preactivated OBs (ratio neutrophils/OB, 20/1), incubated 1 h at 37°C and vigorously washed. Analyses were performed with an Olympus Fluoview 300 microscope using Argon-ion (488 nm) and Helium-neon (543 nm) lasers. Forty-three slices of 0.5 μm were scanned (magnification × 600, plan Apo, NA 1.4 with a kalman filter 3 in order to reconstitute z plans).
Bone sections with gouty tophi were from patients registered in the Department of Pathology that obtained the required informed consents. Bones that contained gouty tophi were without infection. They were decalcified for 18 h using Decalcifier II, as recommended by the manufacturer (Surgipath Medical Industries, Richmond, IL, USA), embedded and fixed in paraffin and processed into consecutive 4–12 μm thick sections using a microtome RM 2135 (Leica, Mississauga, ON, Canada) before staining with hematoxylin and eosin (H&E) or a modified tetrachrome method.21 MSU was confirmed by polarizing light microscopy. Images were previously captured with Nikon Optiphot microscope equipped with a Dage MTI DC-330 camera.
Mineralization
OBs were seeded at 0.2 × 106 cells/well in six-well plates and maintained in αMEM +10% FBS+10 mM β-glycerophosphate (37°C, humidified atmosphere, 5% CO2). Medium was replaced every 3–4 days during 20 days culture. Neutrophils (6 × 106 cells equivalent to neutrophil/OB ratio of 6/1, corresponding to 30% of total neutrophils added initially) were added or not on washed MSU-preactivated OBs at day 8 and replaced every 2 days. To remove non-adherent neutrophils, cells were washed three times with PBS. Cells were then fixed 20 min with buffered formalin and 40 mM ARS at pH 4.0–4.2 were added for 20 min. After four washes with H2O, ARS was extracted by incubating cells with acetic acid. Cells and media were then heated at 85°C, pH 4.2 was restored with NaOH, ARS detection was obtained at 405 nm absorbance.22
Resorption
Osteoclast-like cells (OCs) were generated from human peripheral blood mononuclear cells, as described.12 Briefly, peripheral blood mononuclear cells obtained by centrifugation over Ficoll-Paque were resuspended (5 × 106 cells/ml) in MEM supplemented with 10% FBS and depleted of lymphocytes by adherence (5 × 106 cells/ml, 37°C, 90 min). Adherent cells that contained OC precursors were incubated with MEM supplemented with M-CSF (25 ng/ml), RANKL (40 ng/ml) and 1,25 (OH)2 vit. D3 (10 nM). After 7 days of incubation, adherent cells were treated with accutase and were referred to as human differentiated OCs. OBs (30 000 cells/well in 96-well plates) were cultured on calcified matrix BioCoat™ Osteologic™ Discs (BD biosciences) until confluency in αMEM+10% FBS. Confluent OBs were stimulated with 1 mg MSU/106 cells for 1 h, as above. Neutrophils (150 000/well) were added to MSU-preactivated OBs and replaced once after 2 days. At day 7, neutrophils were removed and 100 000 differentiated OCs were added to MSU-preactivated OBs. Positive controls of resorption were obtained with OCs directly added on Osteologic Discs. Each experiment (from controls without neutrophils to experiments with added neutrophils) was always carried out with the same source of osteoclast precursors to lead to a similar number of differentiated OCs. Incubations with differentiated OCs were pursued for 13 days before evaluation of resorption pit areas by microscopy, as previously described.12
Lactoferrin
Neutrophils (107/ml) were incubated for 2 h (37°C) with 0.5 mg/ml MSU in HBSS alone or with OBs (neutrophil/OB: 10/1) preincubated with or without 200 nM RAP for 3 h (37°C). Neutrophils were also incubated with 0.5 mg/ml MSU above cell culture inserts (0.4 μm pore size, BD Falcon, Franklin Lakes, NJ, USA) disposed over OBs preactivated with or without 1 mg/ml MSU. LF in supernatants was measured by ELISA, as described.23
Cytofluorometry
To evaluate surface markers, cells were incubated (4°C) 20 min with 10% normal goat or human serum (blocking step) and for another 20 min with PE-, FITC-conjugated mAb, or anti-human neutrophil elastase mAb (2 μg/ml) followed by a secondary FITC-conjugated Ab. The cellular viability was analyzed by flow cytometry. OBs (106) were preactivated or not with 1 mg/ml MSU for 1 h and after two vigorous washes were incubated with or without neutrophils (ratio neutrophils/OB, 10/1) for 48 h (37°C) and then removed using accutase. Necrotic and late apoptotic cells were identified by PI incorporation. Neutrophils were incubated for 48 h (37°C) with or without confluent OBs in the presence or absence of 0.5 mg/ml MSU, then removed and analyzed by a flowcytometer EPICS-XL (Beckman Coulter, Miami, FL, USA). Four thousand events were counted and neutrophils and OBs were analyzed on the basis of forward and side-light scatters. These cells were easily individualized by their forward-light scatter, which was established at ≤60 000 and ≥100 000 arbitrary units for neutrophils and OBs, respectively.
Statistics
Results are mean values±s.e.m. Analyses were performed with Instat 3.0 (GraphPad Software, San Diego, CA, USA). Comparisons between two groups were analyzed by paired or unpaired t-tests. Comparisons between more than two groups were studied by a one-way ANOVA with repeated measures and Bonferroni post hoc test. Significance was set at P<0.05.
RESULTS
Modifications of OB Morphology Induced by Neutrophils
Since MSU-stimulated OBs produce IL-8,6 neutrophils present in bone space are likely attracted towards activated OBs, suggesting possible cell-to-cell interactions. Knowing that MSU can affect cellular viability,24 the effect of MSU on neutrophil and OB viability was evaluated before studying interactions between neutrophils and OBs. MSU did not alter neutrophil survival, and neutrophils with OBs in basal conditions or with MSU-preactivated OBs showed increased viability (Figure 1a). MSU (up to 1 mg/ml) or neutrophils added to OBs did not significantly modify OB viability (Figure 1b).

Reciprocal effects of neutrophils on OBs. (a) Neutrophils alone (Control) were stimulated or not with MSU, or coincubated with confluent OBs (preactivated or not with MSU) at 37°C for 48 h. (b) Controls were OBs alone or incubated with neutrophils without MSU at 37°C for 48 h. OBs were preactivated by MSU (+MSU) and then incubated at 37°C for 48 h. Propidium iodide incorporation by necrotic or late apoptotic neutrophils (a) or OBs (b) was evaluated by cytofluorometry. Results are expressed in percentages of viable cells (mean values±s.e.m.; a, n=7; b, n=6). (c) Confluent resting OBs (1, 2, 3; magnified views 1a, 2a, 3a) or OBs preactivated (4, 5, 6) with MSU, were coincubated with (images 2, 3, 5, 6) or without (images 1 and 4) neutrophils for 48 h at 37°C. Control (1, 2, 3) and control (1, 1a, 4) were cells without MSU or without neutrophils and insert, respectively. To avoid cellular contact, 0.4 μm cell culture insert were used between both cell types (images 3, 3a and 6). OB morphology was analyzed by an Olympus research inverted microscope CKX41 (magnification × 200). (d, e) Effects of neutrophil concentration on OBs and kinetic studies of the neutrophil effect. Confluent OBs were incubated with neutrophils at neutrophils/OB ratio 0/1 (control), 2/1, 6/1, 10/1 for 48 h. (e) Kinetic studies of the neutrophil effect. Confluent OBs were incubated with neutrophils for 18, 24, 48, and 72 h. Neutrophils were removed and cell-free area was determined, as described in Materials and methods. (f) Reversal of the neutrophil effect on OBs. Confluent OBs were incubated with neutrophils for 48 h (white histogram). Neutrophils were then removed by washing and OB culture was pursued for 48 h (black histogram). (g) Cytofluorometry analysis of elastase expression by neutrophils incubated alone (dotted line) or with OBs (plain line) for 24 h (representative of five donors). (h) Confluent OBs were incubated with purified neutrophil elastase at 0.5, 1, and 3 μg/ml for 6 h. (i) Neutrophils were preincubated or not with 100 μM elastase inhibitor for 30 min and then incubated with confluent OBs, for 48 h. Cell-free areas were determined, as described in Materials and methods. Experiments in (d–f) (n=6) were performed with two different OB donors each with three different neutrophil donors. Experiments in (h) were performed with three different OB donors and experiments in (i) with two different OB donors each with six different neutrophil donors. Statistical analyses: (a (#P<0.02), f) paired two-tailed t-test; (d, e, h, i) repeated measures ANOVA with Bonferroni's multiple comparison test.
Neutrophils were found to markedly alter OB morphology after 48 h with or without MSU preactivation of OBs (Figure 1c). OBs changed from usual cuboidal shape to unusual stellate morphology with polygonal or spindle shape. Confluent OBs typically arranged in palisade-like monolayers were induced by neutrophils to greatly retract with an increase in areas of underlying cell-free substratum (Figure 1c1, 1c1a, 1c2, and 1c2a). Preactivation of OBs by MSU did not modify the induction of OB retraction by neutrophils (Figure 1c4 and 1c5). This neutrophil effect on OB morphology was dependent on cellular contact, since it was abrogated by adding a microporous membrane between neutrophils and OBs (Figure 1c3, 1c3a, and 1c6). It was also dependent on concentration of neutrophils added to confluent OBs with a threshold of 2/1 (neutrophil/OB ratio) and a significant effect at a 6/1 ratio (Figure 1d). This latter ratio corresponds to the percentage of neutrophils (30%) that remained adherent to MSU-preactivated OBs (see below), and was used for long-term incubations (>24 h). Kinetic studies with a 6/1 ratio showed that this effect was detected within 18 h and was significant after 48 h (Figure 1e). Neutrophils removed after 48 h of interactions with OBs led to a reversal of the neutrophil effect and continuing the culture for 48 h without neutrophils allowed to cover by OBs 70±9% of cell-free areas (Figure 1f). The stellate morphology of OBs that was acquired during contact with neutrophils disappeared and OBs retrieved their palisade-like arrangement (not shown). Interestingly, neutrophils have been shown to induce endothelial detachment and retraction through membrane elastase.25 After a 24-h incubation of neutrophils with OBs, analysis by cytofluorometry of elastase on neutrophil surface (Figure 1g) showed an increased number of elastase-expressing neutrophils from 3.0±0.2% to 9.7±1.2% (P=0.0018, n=5, unpaired t-test). Incubation of OBs with purified neutrophil elastase was associated with OB retraction concomittent to a loss of their palisade-like arrangement and an increase of cell-free underlying matrix (Figure 1h). The present effect of neutrophils on OB arrangement and morphology was significantly attenuated by preincubating neutrophils with an elastase inhibitor (Figure 1i).
Bone Mineralization and Resorption in the Presence of Neutrophils
Could alteration of OB morphology by neutrophils, as demonstrated above, be associated with modifications of OB functions and bone remodeling? Long-term neutrophil effect on OBs cultured in appropriate conditions of mineralization was investigated by staining OBs with alizarin to evaluate calcium-rich deposits.22 Confluent OBs were typically organized in nodular zones with homogeneous ARS staining (Figure 2a1). Neutrophils present in OB cultures led to OB retraction with an increased concentration of ARS in zones of aggregated OB (Figure 2a2). ARS concentrations were, however, similar in both conditions (Figure 2b). These data indicate that neutrophils, while inducing diffuse OB retraction, did not alter quantitatively matrix mineralization by retracted OBs. However, this mineralization was irregular with ARS concentrated under retracted OBs only. Additionally, these morphologic modifications could be associated with increased bone resorption. Note that OCs are greatly implicated in erosive gout.5 Differentiated OCs added to MSU-preactivated OBs arranged in confluent monolayers had a very low resorption capacity of the underlying calcified matrix. Neutrophils added to MSU-preactivated OB cultures in the presence of OCs led to appearance of large resorption areas of the cell-free matrix (Figure 2c–e). In these conditions, remaining neutrophils did not influence resorption activity of OCs (resorption area: 4156±1053 vs 3976±257 arbitrary units; number of pits: 69±9 vs 69±4; OCs alone vs OCs+remaining neutrophils; n=3).

Bone remodeling in the presence of neutrophils. (a) Mineralization by MSU-preactivated OBs with vehicle (image 1) or with neutrophils (image 2) added every 2 days from days 8 to 20 of culture (representative of three different experiments). (b) ARS analyses were performed at day 20. Values are expressed in μM (mean values±s.e.m.; three different donors of OBs, two different donors of neutrophils). (c–e) Resorption of a calcified matrix by human differentiated OCs. OBs were cultured on Osteologic™ Discs, were activated by MSU, and incubated with or without neutrophils added every 2 days for 7 days. After elimination of non-adherent neutrophils, differentiated OCs were added for 13 days of culture. Differentiated OCs cultured alone on Osteologic™ Discs for 13 days were positive controls of bone resorption. Illustration of resorption lacunae by active OCs (c). Results are expressed in arbitrary units of total area of resorption (d) and in number of resorption pits (e). Values are mean values±s.e.m. (two donors of OBs, two donors of OCs, and four donors of neutrophils). Statistical analyses: *P<0.05 (unpaired two-tailed t-test).
Adherence of Neutrophils to OBs
Since OB retraction induced by neutrophils required direct contact, we next determined the adherence capacity of neutrophils to MSU-preactivated OBs. Preincubation of OBs with 1 mg/ml MSU (optimal condition for neutrophil adherence: neutrophil/OB: 20/1) stimulated neutrophil adherence on OBs by 45–50-fold (Figure 3a). After washes, 28.7±5.4% (n=12) of the neutrophils added to OBs remained adherent. Neutrophils firmly adhered to OBs with an intimate contact between the two cells (Figure 3b). The neutrophil membrane in contact with OBs displayed invaginations and expansions with local concentration of the fluorescent dye PKH-67. Alive MSU-preactivated OBs or OBs previously fixed with PFA similarly stimulated neutrophil adherence (data not shown).

Adherence of neutrophils to MSU-preactivated OBs. (a) Confluent OBs were unstimulated (Control) or stimulated with MSU and washed before addition of calcein-AM-prelabeled neutrophils (neutrophil/OB: 20/1) for a subsequent incubation of 1 h at 37°C. Fluorescent cells were counted by epifluorescence microscopy (magnification × 100). Results are expressed in cell number of adherent neutrophils (mean values±s.e.m., n=15). (b) Confluent OBs, previously stained with CMTMR, were stimulated with 1 mg/ml MSU for 1 h at 37°C, washed and fixed with 4% paraformaldehyde. Neutrophils, prelabeled with Green Fluorescent Linker PKH-67, were incubated with OBs for 1 h at 37°C. Cells were washed and visualized by confocal microscopy (magnification × 1500, plan Apo, NA 1.4 with a kalman filter 3 to reconstitute z plans; 43 slices of 0.5 μm were scanned). 1: z plan of y axis; 2: stacking view; 3: z plan of x axis. (c–f) Inhibition of neutrophil degranulation of LF by MSU-preactivated OBs. Neutrophils were incubated with or without MSU (control) and with or without confluent OBs preactivated or not by MSU (c). Neutrophils were incubated with MSU in control medium or in conditioned medium (CM) obtained from OBs preactivated by MSU during 2 h (d). Cell culture inserts were disposed between neutrophils and OBs, the other experimental conditions were similar to c (e). OBs were preincubated with vehicle or RAP, the other experimental conditions were similar to c (f). LF was measured by ELISA and results expressed in ng/ml. Values are mean values±s.e.m. (n=15 in c, n=5 in d, n=6 in e, f corresponding to at least two donors of OBs and three donors of neutrophils). Statistical analyses: unpaired two-tailed t-test, ***P<0.001 (a); one-way ANOVA followed by Bonferroni post hoc test (c), or paired two-tailed t-test (d–f): *P<0.05, **P<0.01. In brackets: percentages of inhibition (mean values±s.e.m.).
Inhibition of Neutrophil Release of LF by OBs
Neutrophil adhesion increases protein release from secondary granules which contain LF, a glycoprotein increasing anabolic functions of OBs while inhibiting osteoclastogenesis.26, 27 MSU addition to neutrophils stimulated LF release (Figure 3c). OBs did not release detectable amounts of LF. Neutrophils added to MSU-preactivated OBs did not show increased LF release (Figure 3c), which indicates an OB inhibitory effect on neutrophil degranulation. Conditioned medium from MSU-preactivated OBs reduced by 30% only LF release (Figure 3d). A similar 30% reduction was obtained when neutrophils were not in direct contact with OBs (Figure 3e). Since LF is rapidly endocytosed by OBs, similar experiments were performed with RAP that inhibits LF endocytosis.28 RAP did not alter the inhibition of LF release (Figure 3f). OBs previously fixed with 4% PFA failed to inhibit neutrophil degranulation in the same conditions (data not shown). Together, these data indicate that live OBs inhibited neutrophil degranulation mainly through direct cell–cell contact, and to a lesser extent by soluble factors.
Integrin Involvement in Neutrophil Adhesion to OBs
A corollary of the above results is that cell-to-cell interactions are important to neutrophil effects on OBs. Adherent neutrophils express the β1 integrin CD29/CD49d, and various inflammatory conditions increase neutrophil expression of the subunit CD49d.14, 29, 30 Neutrophils also express the β1 integrin subunit CD49e.31 CD29 and ICAM-1 were expressed by OBs, while CD49d and CD49e were not (Table 1). None of the integrin subunits studied was upregulated by MSU on OBs (Table 1). Stimulation of neutrophils by MSU increased by threefold the number of cells that expressed CD49d with no effect of cycloheximide (Table 1).
Preincubation of neutrophils with blocking anti-CD49d Ab inhibited their adhesion to MSU-preactivated OBs by 34% (Table 2), suggesting that the β1 integrin CD29/CD49d is one of the adhesion pathways implicated. Although the integrins VCAM-1 and MadCAM-1 are possible counter-receptors of the β1 integrin CD29/CD49d,32 blocking anti-VCAM-1 and anti-MadCAM-1 Ab did not significantly modify the adhesion of neutrophils to OBs (Table 2).
The β1 integrin CD29/CD49d is also known to adhere to the peptide CS1 of fibronectin, which is an extracellular matrix component provided by OBs.33, 34 Neutrophil preincubation with CS1 peptide significantly reduced the adhesion by 30% (P=0.0019), a result similar to that obtained with the blocking anti-CD49d Ab. No additive nor subadditive inhibitory effect was recorded by the combined use of the CS1 peptide and anti-CD49d Ab (Table 2). These results indicate that CS1 is the only OB counter-receptor for the β1 integrin CD29/CD49d present on neutrophils. Thus, the heterotypic association of the β1 integrin subunit CD49d to the fibronectin peptide CS1 accounts for one third of neutrophil adhesion to OBs.
Implication of Bioactive Lipids in Neutrophil Adhesion to OBs
The bioactive lipids LTB4 and PAF increase neutrophil adhesion to cells by modulating adhesion molecules.18, 35, 36 These lipids are rapidly produced by activated neutrophils from arachidonic acid and lyso-PAF, which are simultaneously released from 1-O-alkyl-2-arachidonoyl-glycerophosphocholine through the action of the cytosolic phospholipase A2α (cPLA2α). Arachidonic acid is converted in LTB4 through the 5-lipoxygenase pathway, and lyso-PAF is converted into PAF by a specific acetyl-transferase. Neutrophil adhesion to MSU-preactivated OBs was inhibited by 39% when neutrophils were preincubated with the cPLA2α inhibitor pyrrophenone, which blocks both the biosynthesis of LTB4 and PAF. The PAF and LTB4 receptor antagonists BN52021 and CP105696 reduced neutrophil adhesion by 42 and 38%, respectively. In combination, BN52021+CP105696 inhibited neutrophil adhesion to OBs by 69% (Table 2). The effect of pyrrophenone was reversed by the addition of 10−10 M LTB4 or PAF (Table 2). In further support of a role of LTB4 and PAF in neutrophil–OB adhesion, 10−7 M exogenous LTB4 or PAF increased by threefold neutrophil adhesion to MSU-preactivated OBs. Interestingly, LTB4 and PAF had no effect on neutrophil adhesion to non-preactivated OBs (data not shown).
NS-398, a COX-2 inhibitor, decreased neutrophil adhesion to OBs by 32%, while indomethacin had no effect. Surprisingly, no reversal of the NS-398 effect was obtained when the end products (or their stable analogs) PGE2, PGD2, Iloprost, or IBOP were subsequently added (Table 2). These data do not support the involvement of COX-derived products in neutrophil adhesion to OBs, and suggest a non-COX-2-dependent inhibitory effect of NS-398.
The extracellular signal-regulated kinase and p38 mitogen-activated protein kinases both contribute to the activation of cPLA2α.37 Preincubation of neutrophils with SB203580 (p38 inhibitor) or PD98069 (ERK inhibitor) decreased neutrophil adhesion to OBs by 24 and 34%, respectively (Table 2). Preincubation of neutrophils with SB203580 or PD98069 and pyrrophenone showed no additive inhibitory effects on cell adhesion in support of a common pathway between these MAPK and cPLA2α.
The effects of pyrrophenone (cPLA2α inhibitor) and blocking anti-CD49d Ab were subadditive in decreasing the adherence of neutrophils to OBs (Table 2). Neutrophil adherence was inhibited by 34% with anti-CD49d Ab, and by 39% with pyrrophenone alone, while the inhibition raised to 56% when neutrophils were pretreated with the two agents. The inhibition by the two products added together was significantly higher than that of the products added alone. Thus, even if LTB4 or PAF could affect the expression of adhesion molecules, these results indicate that the cPLA2α and CD29/CD49d pathways can independently regulate the adhesion of neutrophils to OBs.
Altered OBs in Tophaceous Gout
Considering that the histopathology of chronic gout shows urate deposits in subchondral bone and deeper in trabecular bone and bone marrow,3 the presence of neutrophils close to bone cells is very likely. Paraffin-embedded section of advanced tophaceous deposits showed white chalky and nodular materials corresponding to deposits of MSU in various tissues including bone structures (Figure 4a). Intramedullary MSU deposits were visualized alongside the endosteal bone where MSU have been dissolved by formalin. This preparation has left pale amorphous crystal-like hyaline areas surrounded by dense fibrous tissue. Polarized light showed a few remaining crystals (Figure 4b). Highly concentrated MSU deposits were also individualized directly in bone matrix (Figure 4c). Several tophaceous nodules were rimmed by inflammatory and necrotic tissues with fibroblasts, giant multinucleated cells, and a mononuclear histiocytic reaction. Certain tophaceous deposits were not surrounded by granulomatous tissue. Modified tetrachrome, which is designed for paraffin-embedded bone after formalin fixation,21 allowed to visualize irregular mineralized bone. Osteoid (deep blue) was clearly distinguished from normal mature bone (red). Pale bluish nodular deposits of MSU were irregularly surrounded by mature lamellar bone (Figure 4d). Moreover, few and irregular osteoid tissues were observed with very rare and dispersed OBs, some of which were directly located on osteoid that showed crystals (Figure 4d, inset). Interestingly, MSU tophi were also present deeper in the trabecular bone with minor inflammation only (Figure 5a), that is reminiscent of Jaffe's findings.3 Polymorphonuclear cells were individualized close to OBs and in the tissues surrounding tophi deeper in the trabecular bone (Figure 5). It is also interesting that bone matrix close to tophi was irregularly calcified as shown by the presence of new incompletely calcified matrix (Figures 4d and 6a). Osteoid tissues were increased with altered morphology of OBs, which were small and dispersed on the osteoid (Figures 4d inset and 6a), rather than large, side-by-side and organized in their typical palisade-like arrangement (Figure 6b). Many isolated OBs were also included into new osteoid tissues (Figures 4d and 6a). OCs were also easily individualized in resorption lacunae close to urate deposits (Figure 6c). The histomorphometric analysis of cortical and trabecular bone (data not shown) revealed a decrease of bone formation (reduction of calcified matrix and OB number), an increase of osteoid tissue, and an important increase of bone resorption (augmentation of eroded bone surfaces and of number of active OCs).

Histopathology of bone with tophaceous deposits. (a) White chalky materials of tophaceous deposits in paraffin-embedded toe, after removal of paraffin. (b) H&E staining under polarized light showing amorphous materials in medullary cavity and alongside the endosteal cortical bone (magnification: × 20). (Inset) Focus on tophus visualized with red filter. (c) Crystalline amorphous materials directly in osteoid and calcified bone matrix (magnification: × 100). (d) Modified tetrachrome staining: bluish crystalline deposits with irregular inflammatory tissues surrounded by dark red mature lamellar bone, and with different degrees of bone mineral maturity showing irregular dark blue osteoid on the surface and inside bone (magnification: × 40). (Inset) Focus on crystals in osteoid; arrows show OBs non-contiguous and located on blue-stained osteoid tissue.

Neutrophils in bone with tophaceous deposits. (a) Giemsa staining of trabecular bone with urate deposit in a zone showing polymorphonuclear leukocytes (magnification: × 20). (b) Enlargement of a zone close to the inferior part of the urate deposit seen in (a). (c) Osteoblasts (OB) are visualized (magnification: × 400) beside a polymorphonuclear cell (Neutro), dispersed on irregular blue osteoid. (Inset) Focus on two OBs below a neutrophil. Note that OBs and the polymorphonuclear cell are not exactly in the plane of focus.

OBs in bone with tophaceous deposits. (a) Modified tetrachrome staining showing multiple bluish urate tophi with major inflammatory tissues, irregular dark blue osteoid and dark red mature bone (image 1; magnification: × 20). Enlargement of osteoid tissue close to a urate tophus seen in image 1: urate tophus, inflammatory tissues, osteoid, and lamellar bone are visualized; white arrows show dispersed OBs retracted without palisade arrangement (image 2; magnification: × 40). Image 3: enlargement of a2 (magnification: × 200). (b) Histology of normal bone. A 7-μm cross-section of transiliac bone biopsy stained by the Masson-Goldner technique shows the calcified bone matrix in green, the osteoid border in red/orange, fibrous and connective tissue in light green, and cells in orange. Note that OBs are regular and large cells, all adjacent side-by-side with a palisade-like arrangement (image 1; magnification: × 40). Image 2: enlargement of b1. (c) H&E staining showing an active osteoclast (OC) in a resorption lacunae where few OBs can be individualized. Note that irregular dark blue osteoid is present on the surface and inside bone adjacent to urate deposits.
Overall, these data suggest that tophaceous deposits in bone greatly affect local remodeling with an imbalance of endosteal bone turnover characterized by: (1) MSU present in bone tissues, in particular in new bone matrix, (2) reduced number of OBs without typical palisade-like arrangement, and (3) increased OC activity corresponding to a great uncoupling of bone cell activity. Polymorphonuclear cells are present close to OBs and bone tophi lesions. It emerges from these histopathological data (1) that OBs are greatly affected by MSU in chronic gout and (2) that local polymorphonuclear cells could affect bone cell functions.
DISCUSSION
This study demonstrates that OB functions are altered in tophaceous gout and that neutrophils can affect OB functions leading to decreased bone formation and increased bone resorption. Histopathology of gouty bones with chronic tophaceous deposits has shown the presence of crystals in bone tissues, MSU deposits in the bone matrix with altered morphology and organization of OBs, and the presence of polymorphonuclear cells close to bone cells. In this regard, the reversibility and short duration of neutrophil adhesion to OBs that we showed in vitro may account for the difficulty of finding such direct contact in tophaceous tissues ex vivo. However, the present data indicated that progressive MSU deposits have prolonged effects on OBs. While reducing bone formation by OBs, MSU stimulate these OBs to generate inflammatory mediators like IL-6 and IL-8,6 factors known to attract and activate neutrophils. Thus, these cells that are present locally in bone microvasculature and marrow space in non-pathological conditions, can easily migrate in different inflammatory bone tissues. Interestingly, the neutrophil concentration that significantly affected OB morphology was close to that of neutrophils remaining adherent (30%) to MSU-preactivated OBs. Then, neutrophils in direct contact with OBs, activated or not by MSU, cause OB retraction leading to a cell-free bone matrix vulnerable to osteoclastic bone resorption. Bone regulatory factors like parathyroid hormone and TGF-β can induce OB elongation and retraction with subsequent increased resorption of the cell-free matrix accessible to active OCs.38, 39 It has also been reported that IL-6 can affect OB arrangement and morphology, and that elastase promotes endothelial cell retraction.25, 40 Importantly, elastase mobilized by activated neutrophils remains bound to their membrane, and is catalytically active.41, 42 We show herein that neutrophils in direct contact with OBs induced their retraction, in analogy to monocytic pre-OCs, which have been shown to activate OB retraction through direct interaction with these cells,43 and that membrane elastase of neutrophils is responsible for OB retraction. Moreover, even if neutrophil adhesion to OBs was not associated with major changes of OB functions, MSU-activated OBs stimulated neutrophil adhesion to these cells, and in turn adherent neutrophils induced morphological changes of OBs and indirectly favored increased bone resorption by activated OCs. It is noteworthy that OCs are greatly increased in bone with tophaceous deposits and that osteoclastogenesis is strongly activated by MSU crystals.5
Reciprocal effects of neutrophil–OB interactions secondary to MSU activation of OBs (changes of OB morphology and neutrophil release of LF) required direct contacts between the two cell types, suggesting a functional importance of the pathways involved in cellular adherence. As visualized by confocal microscopy, adhesion of neutrophils to OBs showed a concentration of the membrane lipid dye at the anchorage zone of neutrophils to OBs. This dye accumulation is reminiscent of that reported for neutrophils adherent to endothelial cells.44 It is also useful to stress that invaginations of neutrophil membrane at the contact area with OBs might correspond to a ruffled border facilitating exchanges, as reported in adherent neutrophils.45 On the other hand, even if the duration of neutrophil adherence to OBs is relatively short (similar numbers of neutrophils adhered to OBs after 2 and 4 h post-addition of neutrophils to MSU-activated OBs; after 18 h, these neutrophils were viable but non-adherent; data not shown), it is noteworthy that (1) neutrophil–OB interactions rapidly alter cellular functions, (2) OBs in the presence of MSU (crystals that persist in bone tissues) chronically produce factors chemotactic for neutrophils, and (3) each OB which is much larger than neutrophils can interact with several neutrophils.
The mechanisms of neutrophil adhesion to OB implicate, at least, the β1 integrin CD29/CD49d present on neutrophils and the fibronectin peptide CS1 as the OB counter-receptor. The inhibition of neutrophil adherence to OB by the anti-CD49d antibody suggested that CD49d was involved in the adhesion. This is in agreement with the increased expression of CD49d by neutrophils, as previously reported,29 and its lack of expression by OBs. In the experimental conditions used, 30% of the neutrophils initially added adhere to MSU-preactivated OBs. Interestingly, the blocking anti-CD49d antibody inhibited the binding of about 40% of those 30% adherent neutrophils, indicating that only 12–15% of the neutrophils can adhere to OBs in a CD49d-dependent manner. Notably, this value corresponds to the percentage of neutrophils expressing CD49d after MSUM stimulation. Moreover, the function-blocking anti-CD49d antibody (clone R1–2), which was raised against a subunit (B1 epitope) involved in heterotypic binding,46 and our findings of the fibronectin peptide CS1 as a counter-receptor of CD49d, confirmed that neutrophil adhesion to OBs is heterotypic. In agreement, the blocking of CD49d or of CS1 similarly inhibited the adherence of neutrophils, and other counter-receptors of CD49d such as VCAM-1 and MAdCAM-1 were not implicated. Moreover, even if the β2 integrins (CD11a–d/CD18) are involved in the adhesion of neutrophils to the vascular endothelium and their transendothelial migration during inflammation,13 these integrins were previously shown to be non-involved in neutrophil adhesion to MSU-preactivated OBs.47
Adhesion of neutrophils to various cell types supports key regulatory processes of inflammation. For example, interactions between neutrophils and inflammatory fibroblasts increase neutrophil survival.48 The present results add the OBs to the cell types that enhance neutrophil viability. Interestingly, OBs have been shown to promote the survival of hematopoietic progenitor cells through cell–cell interactions that depend in part on the β1 integrin CD29/CD49d.49 On the other hand, large amounts of LF are present in neutrophils, and partially engulfed MSU by OBs could activate neutrophils to release LF known to increase bone formation.26 However, OBs inhibit neutrophil release of LF in response to MSU. Thus, the prolonged survival of neutrophils in the presence of MSU and OBs might enhance deleterious effects of neutrophils on bones, particularly since MSU stimulate neutrophil production of reactive oxygen species that can alter mineralization.50, 51
Washings of MSU-preactivated OBs remove non-phagocytized microcrystals; however, 1 h after exposure of OBs to MSU, phagocytized crystals remain incompletely engulfed and therefore may activate neutrophils.6 That neutrophils were still able to adhere to MSU-preactivated OBs fixed by paraformaldehyde implicate that activation of neutrophils by MSU-activated OBs can occur through at least two distinct mechanisms (1) stimulation by incompletely engulfed crystals and (2) stimulation by the OBs through adhesion molecules (CD29/CD49d-CS1). Interestingly, neutrophil interaction with MSU rapidly causes the release of inflammatory bioactive lipids.52 The 39% inhibition of adhesion by pyrrophenone (a cPLA2α inhibitor) and its reversal by 10−10 M LTB4 or PAF confirmed the role of these lipids in neutrophil adherence to OBs. The importance of both lipids was also underscored by the dose-dependent stimulatory effect of both mediators (up to 335±67% and 206±23% at 10−8 M LTB4 or 10−7 M PAF, respectively) on neutrophil adherence to OBs, an effect that required OB preactivation by MSU (data not shown), and by the inhibitory effects of LTB4 and PAF antagonists. Both lipids have been shown to increase neutrophil adhesion to other mesenchymal cells.17, 18 Importantly, PAF and LTB4 directly act on neutrophil adhesion to MSU-preactivated OBs since PAF and LTB4 antagonists showed additive effects. Hence, PAF has been reported to stimulate elastase mobilization at the membrane of neutrophils, in particular at the active front of migrating neutrophils.53 This stimulation might be associated with the neutrophil-induced retraction of OBs, as discussed above. On the other hand, LTB4 has been shown to activate neutrophil adhesion by increasing neutrophil surface expression of prestocked β2 integrin.54 However, neutrophil adhesion to MSUM-preactivated OB was not inhibited by blocking the β2 integrin pathway.47 Taken together, these data and the observation of the partially additive effects of pyrrophenone and anti-CD49d Ab demonstrate that the effect of LTB4 in neutrophil adhesion to MSU-preactivated OBs is partly independent of the β1 and β2 integrin pathways investigated. On the other hand, the absence of additive effects of p38 and ERK inhibitors with pyrrophenone on neutrophil adhesion to OBs is compatible with the implication of p38 kinase and ERK in the regulation of cPLA2α, as previously reported.37 Furthermore, COX-derived products were not involved in neutrophil adhesion to OBs since indomethacin, which inhibits both COXs, did not modify this adhesion. The absence of reversal effect of PGs, IBOP, or Iloprost on NS-398-induced inhibition suggests a non-COX-2-dependent inhibitory effect of NS-398. Accordingly, NS-398 possesses inhibitory activities independent of COX-2 and PGE2.55, 56
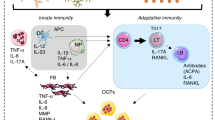
In conclusion, MSU promote the adhesion of neutrophils to OBs via CD49d/CS1, LTB4- and PAF-dependent mechanisms. These findings highlight novel aspects of the deregulation of bone physiology in lesions of chronic gout where bone cells are in direct contact with MSU. By releasing IL-8 and IL-6, and by enhancing neutrophil survival, OBs can be implicated in the local inflammatory process of gout. Accordingly, in vivo experiments in rodents have shown the capacity of neutrophils to inhibit OBs and activate OCs, whereas LF had the opposite effects.26, 57, 58 Inflammatory conditions also stimulate neutrophil expression of regulatory factors involved in bone remodeling like RANK, RANKL, and OPG.12, 59 Therefore, neutrophil adhesion to OBs could impact several regulatory mechanisms of cellular functions implicated in the abnormal bone remodeling observed in chronic gout as summarized in Figure 7. It emerges from the present findings that neutrophils and OBs might be important players in bone lesions of chronic gout, and may represent targets for therapies and prevention of bone damage.

Schematic representation of pathological sequences associated with MSU deposits in bone. (1) Initiation: progressive MSU deposits in bone tissues where osteoblasts (OB) have the capacity to ingest crystals leading to reduction of bone formation and to production of inflammatory mediators.6 Cytokines like IL-6 and IL-8 produced by MSU-activated OBs are known to affect neutrophils that are greatly present in bone spaces.11, 60 (2) MSU-activated OBs share the function of greatly inducing adhesion of locally present neutrophils which in turn induce OB retraction. (3) Cell-free matrix related to OB retraction becomes vulnerable to functionally active OCs locally present. Bioactive lipids like PGE2 generated by MSU-activated OBs activates OC functions that are also largely stimulated by MSU-activated stromal cells.5 The overal effects of neutrophil–OB interactions in the presence of MSU are increased neutrophil survival, promotion of important bone resorption with no possible compensation by bone anabolic effects of lactoferrin (LF), which is reduced by neutrophil–OB interactions. Direct interactions of neutrophil with OBs are possible through the α4 subunit CD49d of β1 integrins and the fibronectin peptide CS1. These interactions are also affected by PAF and LTB4, bioactive lipids that are known to impact on cellular adhesion through PAF receptors and release of granules.17, 18 OB retraction induced by neutrophils require direct contact of OBs with neutrophils leading to the effect of neutrophil elastase located at the membrane.
References
Monu JU, Pope Jr TL . Gout: a clinical and radiologic review. Radiol Clin North Am 2004;42:169–184.
Dalbeth N, Clark B, Gregory K, et al. Mechanisms of bone erosion in gout: a quantitative analysis using plain radiography and computed tomography. Ann Rheum Dis 2009;68:1290–1295.
Jaffe HL . Gout. In: Jaffe HL (ed). Metabolic, Degenerative and Inflammatory Diseases of Bones and Joints. Lea & Febiger, Publishers. Philadelphia, PA, 1972, pp 479–491.
Hasselbacher P . Stimulation of synovial fibroblasts by calcium oxalate and monosodium urate monohydrate. A mechanism of connective tissue degradation in oxalosis and gout. J Lab Clin Med 1982;100:977–985.
Dalbeth N, Smith T, Nicolson B, et al. Enhanced osteoclastogenesis in patients with tophaceous gout: urate crystals promote osteoclast development through interactions with stromal cells. Arthritis Rheum 2008;58:1854–1865.
Bouchard L, de Medicis R, Lussier A, et al. Inflammatory microcrystals alter the functional phenotype of human osteoblast-like cells in vitro: synergism with IL-1 to overexpress cyclooxygenase-2. J Immunol 2002;168:5310–5317.
Siddiqi A, Burrin JM, Wood DF, et al. Tri-iodothyronine regulates the production of interleukin-6 and interleukin-8 in human bone marrow stromal and osteoblast-like cells. J Endocrinol 1998;157:453–461.
Mitsuyama K, Toyonaga A, Sasaki E, et al. IL-8 as an important chemoattractant for neutrophils in ulcerative colitis and Crohn's disease. Clin Exp Immunol 1994;96:432–436.
Schumacher HR, Phelps P . Sequential changes in human polymorphonuclear leukocytes after urate crystal phagocytosis. An electron microscopic study. Arthritis Rheum 1971;14:513–526.
Girasole G, Jilka RL, Passeri G, et al. 17 beta-estradiol inhibits interleukin-6 production by bone marrow-derived stromal cells and osteoblasts in vitro: a potential mechanism for the antiosteoporotic effect of estrogens. J Clin Invest 1992;89:883–891.
Kitamura H, Kawata H, Takahashi F, et al. Bone marrow neutrophilia and suppressed bone turnover in human interleukin-6 transgenic mice. A cellular relationship among hematopoietic cells, osteoblasts, and osteoclasts mediated by stromal cells in bone marrow. Am J Pathol 1995;147:1682–1692.
Chakravarti A, Raquil MA, Tessier P, et al. Surface RANKL of toll-like receptor 4-stimulated human neutrophils activates osteoclastic bone resorption. Blood 2009;114:1633–1644.
Gao JX, Issekutz AC . Mac-1 (CD11b/CD18) is the predominant beta 2 (CD18) integrin mediating human neutrophil migration through synovial and dermal fibroblast barriers. Immunology 1996;88:463–470.
Kubes P, Niu XF, Smith CW, et al. A novel beta 1-dependent adhesion pathway on neutrophils: a mechanism invoked by dihydrocytochalasin B or endothelial transmigration. FASEB J 1995;9:1103–1111.
Reinhardt PH, Elliott JF, Kubes P . Neutrophils can adhere via alpha4beta1-integrin under flow conditions. Blood 1997;89:3837–3846.
Gronthos S, Stewart K, Graves SE, et al. Integrin expression and function on human osteoblast-like cells. J Bone Miner Res 1997;12:1189–1197.
Damtew B, Spagnuolo PJ . Platelet activating factor amplifies human neutrophil adherence to bovine endothelial cells: evidence for a lipoxygenase dependent mechanism. Inflammation 1992;16:425–436.
Hoover RL, Karnovsky MJ, Austen KF, et al. Leukotriene B4 action on endothelium mediates augmented neutrophil/endothelial adhesion. Proc Natl Acad Sci USA 1984;81:2191–2193.
Roberge CJ, de Medicis R, Dayer JM, et al. Crystal-induced neutrophil activation. V. Differential production of biologically active IL-1 and IL-1 receptor antagonist. J Immunol 1994;152:5485–5494.
Flamand N, Picard S, Lemieux L, et al. Effects of pyrrophenone, an inhibitor of group IVA phospholipase A2, on eicosanoid and PAF biosynthesis in human neutrophils. Br J Pharmacol 2006;149:385–392.
Ralis ZA, Watkins G . Modified tetrachrome method for osteoid and defectively mineralized bone in paraffin sections. Biotech Histochem 1992;67:339–345.
Gregory CA, Gunn WG, Peister A, et al. An Alizarin red-based assay of mineralization by adherent cells in culture: comparison with cetylpyridinium chloride extraction. Anal Biochem 2004;329:77–84.
Rusu D, Drouin R, Pouliot Y, et al. A bovine whey protein extract can enhance innate immunity by priming normal human blood neutrophils. J Nutr 2009;139:386–393.
Akahoshi T, Nagaoka T, Namai R, et al. Prevention of neutrophil apoptosis by monosodium urate crystals. Rheumatol Int 1997;16:231–235.
Yoshida N, Cepinskas G, Granger DN, et al. Aspirin-induced, neutrophil-mediated injury to vascular endothelium. Inflammation 1995;19:297–312.
Cornish J, Callon KE, Naot D, et al. Lactoferrin is a potent regulator of bone cell activity and increases bone formation in vivo. Endocrinology 2004;145:4366–4374.
Xu X, Hakansson L . Degranulation of primary and secondary granules in adherent human neutrophils. Scand J Immunol 2002;55:178–188.
Naot D, Grey A, Reid IR, et al. Lactoferrin—a novel bone growth factor. Clin Med Res 2005;3:93–101.
Ibbotson GC, Doig C, Kaur J, et al. Functional alpha4-integrin: a newly identified pathway of neutrophil recruitment in critically ill septic patients. Nat Med 2001;7:465–470.
Reinhardt PH, Naccache PH, Poubelle PE, et al. Monosodium urate crystals promote neutrophil adhesion via a CD18-independent and selectin-independent mechanism. Am J Physiol 1996;270:C31–C39.
van den Berg JM, Mul FP, Schippers E, et al. Beta1 integrin activation on human neutrophils promotes beta2 integrin-mediated adhesion to fibronectin. Eur J Immunol 2001;31:276–284.
Davenpeck KL, Sterbinsky SA, Bochner BS . Rat neutrophils express alpha4 and beta1 integrins and bind to vascular cell adhesion molecule-1 (VCAM-1) and mucosal addressin cell adhesion molecule-1 (MAdCAM-1). Blood 1998;91:2341–2346.
Puleo DA, Bizios R . Mechanisms of fibronectin-mediated attachment of osteoblasts to substrates in vitro. Bone Miner 1992;18:215–226.
van Dinther-Janssen AC, Pals ST, Scheper RJ, et al. Role of the CS1 adhesion motif of fibronectin in T cell adhesion to synovial membrane and peripheral lymph node endothelium. Ann Rheum Dis 1993;52:672–676.
Arnould T, Michiels C, Remacle J . Increased PMN adherence on endothelial cells after hypoxia: involvement of PAF, CD18/CD11b, and ICAM-1. Am J Physiol 1993;264:C1102–C1110.
Patarroyo M, Prieto J, Rincon J, et al. Leukocyte-cell adhesion: a molecular process fundamental in leukocyte physiology. Immunol Rev 1990;114:67–108.
Hiller G, Sundler R . Activation of arachidonate release and cytosolic phospholipase A2 via extracellular signal-regulated kinase and p38 mitogen-activated protein kinase in macrophages stimulated by bacteria or zymosan. Cell Signal 1999;11:863–869.
Karsdal MA, Fjording MS, Foged NT, et al. Transforming growth factor-beta-induced osteoblast elongation regulates osteoclastic bone resorption through a p38 mitogen-activated protein kinase- and matrix metalloproteinase-dependent pathway. J Biol Chem 2001;276:39350–39358.
Murray EJ, Tram KK, Spencer MJ, et al. PTH-mediated osteoblast retraction: possible participation of the calpain pathway. Miner Electrolyte Metab 1995;21:184–188.
Sylvester FA, Wyzga N, Hyams JS, et al. Effect of Crohn's disease on bone metabolism in vitro: a role for interleukin-6. J Bone Miner Res 2002;17:695–702.
Gaudin P, Berthier S, Barro C, et al. Proteolytic potential of human neutrophil membranes. Eur J Cell Biol 1997;72:345–351.
Owen CA, Campbell MA, Sannes PL, et al. Cell surface-bound elastase and cathepsin G on human neutrophils: a novel, non-oxidative mechanism by which neutrophils focus and preserve catalytic activity of serine proteinases. J Cell Biol 1995;131:775–789.
Perez-Amodio S, Beertsen W, Everts V . (Pre-)osteoclasts induce retraction of osteoblasts before their fusion to osteoclasts. J Bone Miner Res 2004;19:1722–1731.
Rochon YP, Kavanagh TJ, Harlan JM . Analysis of integrin (CD11b/CD18) movement during neutrophil adhesion and migration on endothelial cells. J Microsc 2000;197:15–24.
Albertine KH, Cerasoli Jr F, Gee MH, et al. Morphological analysis of the activation of adherent neutrophils in vitro. Tissue Cell 1988;20:519–530.
Kamata T, Puzon W, Takada Y . Identification of putative ligand-binding sites of the integrin alpha 4 beta 1 (VLA-4, CD49d/CD29). Biochem J 1995;305 (Pt 3):945–951.
Bouchard L, Naccache PH, Poubelle PE . Promotion of neutrophil adherence to human osteoblasts by microcrystals and f-Met-Leu-Phe. Biochem Biophys Res Commun 2002;296:759–764.
Ling CJ, Owen Jr WF, Austen KF . Human fibroblasts maintain the viability and augment the functional response of human neutrophils in culture. J Clin Invest 1990;85:601–604.
Jung Y, Wang J, Havens A, et al. Cell-to-cell contact is critical for the survival of hematopoietic progenitor cells on osteoblasts. Cytokine 2005;32:155–162.
Lee DH, Lim BS, Lee YK, et al. Effects of hydrogen peroxide (H2O2) on alkaline phosphatase activity and matrix mineralization of odontoblast and osteoblast cell lines. Cell Biol Toxicol 2006;22:39–46.
Naccache PH, Grimard M, Roberge CJ, et al. Crystal-induced neutrophil activation. I. Initiation and modulation of calcium mobilization and superoxide production by microcrystals. Arthritis Rheum 1991;34:333–342.
Poubelle PE, De Medicis R, Naccache PH . Monosodium urate and calcium pyrophosphate crystals differentially activate the excitation-response coupling sequence of human neutrophils. Biochem Biophys Res Commun 1987;149:649–657.
Cepinskas G, Sandig M, Kvietys PR . PAF-induced elastase-dependent neutrophil transendothelial migration is associated with the mobilization of elastase to the neutrophil surface and localization to the migrating front. J Cell Sci 1999;112 (Pt 12):1937–1945.
Patcha V, Wigren J, Winberg ME, et al. Differential inside-out activation of beta2-integrins by leukotriene B4 and fMLP in human neutrophils. Exp Cell Res 2004;300:308–319.
Denkert C, Furstenberg A, Daniel PT, et al. Induction of G0/G1 cell cycle arrest in ovarian carcinoma cells by the anti-inflammatory drug NS-398, but not by COX-2-specific RNA interference. Oncogene 2003;22:8653–8661.
Xu H, Izon DJ, Loftin C, et al. The COX-2 inhibitor NS-398 causes T-cell developmental disruptions independent of COX-2 enzyme inhibition. Cell Immunol 2001;214:184–193.
Chung R, Cool JC, Scherer MA, et al. Roles of neutrophil-mediated inflammatory response in the bony repair of injured growth plate cartilage in young rats. J Leukoc Biol 2006;80:1272–1280.
Kuwabara H, Wada T, Oda T, et al. Overexpression of the granulocyte colony-stimulating factor gene impairs bone morphogenetic protein responsiveness in mice. Lab Invest 2001;81:1133–1141.
Poubelle PE, Chakravarti A, Fernandes MJ, et al. Differential expression of RANK, RANK-L, and osteoprotegerin by synovial fluid neutrophils from patients with rheumatoid arthritis and by healthy human blood neutrophils. Arthritis Res Ther 2007;9:R25.
Seo SM, McIntire LV, Smith CW . Effects of IL-8, Gro-alpha, and LTB(4) on the adhesive kinetics of LFA-1 and Mac-1 on human neutrophils. Am J Physiol Cell Physiol 2001;281:C1568–C1578.
Acknowledgements
We thank L Bouchard for her technical assistance, J Garneau for human bone pieces, M Dufour for cytofluorometry analyses, J-C Lévesque for her assistance in confocal microscopy, and C Gilbert for her useful comments. This work was supported by the Canadian Institutes for Health Research and the National Institutes of Health, USA (#NIH/NIAMS 1 R01 AR52614-01).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Bone with urate tophi shows altered remodeling and cellular distribution. Osteoblasts exposed to urate promote neutrophil adherence; and neutrophils, through membrane elastase, induce retraction of confluent osteoblasts. Neutrophil adherence is mediated by the CD29/CD49d-fibronectin peptide CS1 receptors and cPLA2α-derived metabolites, and impacts osteoblast and osteoclast functions.
Rights and permissions
About this article
Cite this article
Allaeys, I., Rusu, D., Picard, S. et al. Osteoblast retraction induced by adherent neutrophils promotes osteoclast bone resorption: implication for altered bone remodeling in chronic gout. Lab Invest 91, 905–920 (2011). https://doi.org/10.1038/labinvest.2011.46
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.2011.46
Keywords
This article is cited by
-
Association of neutrophil to lymphocyte ratio with bone mineral density in post-menopausal women: a systematic review and meta-analysis
BMC Women's Health (2024)
-
Immune microenvironment: novel perspectives on bone regeneration disorder in osteoradionecrosis of the jaws
Cell and Tissue Research (2023)
-
Neutrophil extracellular traps induce the bone erosion of gout
BMC Musculoskeletal Disorders (2022)
-
Study on the relationship between FFA and gout flare
Clinical Rheumatology (2020)
-
NLRP3 promotes autophagy of urate crystals phagocytized by human osteoblasts
Arthritis Research & Therapy (2013)
