
Abstract
Matrix metalloproteinase-7 (MMP-7) (also known as matrilysin-1) is secreted as a proenzyme (proMMP-7) and plays a key role in the degradation of various extracellular matrix (ECM) and non-ECM molecules after activation. To identify the binding proteins related to proMMP-7 activation, a human lung cDNA library was screened by yeast two-hybrid system using proMMP-7 as bait. We identified a candidate molecule CD151, which is a member of the transmembrane 4 superfamily. Complex formation of proMMP-7 with CD151 was demonstrated by immunoprecipitation of the molecules from CaR-1 cells, a human rectal carcinoma cell line, expressing both proMMP-7 and CD151, and CD151 stable transfectants incubated with proMMP-7. Yeast two-hybrid assays using deletion mutants of proMMP-7 and CD151 suggested an interaction between the propeptide of proMMP-7 and the COOH-terminal extracellular loop of CD151. The binding activity of 125I-labeled proMMP-7 to CD151 on the cell membranes was shown with CD151 stable transfectants. Laser-scanning confocal microscopy demonstrated that proMMP-7 colocalizes with CD151 on the cell membranes of CD151 stable transfectants and CaR-1 cells. In situ zymography using crosslinked carboxymethylated transferrin, a substrate of MMP-7, demonstrated proteinase activity on and around CD151 stable transfectants and CaR-1 cells, while the activity was abolished by their treatment with MMP inhibitors, anti-MMP-7 antibody or anti-CD151 antibody. In human lung adenocarcinoma tissues, colocalization of MMP-7 and CD151 was demonstrated on the carcinoma cells. Metalloproteinase activity was present in these tissues and could be inhibited by antibodies to MMP-7 or CD151. These data demonstrate for the first time that proMMP-7 is captured and activated on the cell membranes through interaction with CD151, and suggest the possibility that similar to the MT1-MMP/MMP-2 system, MMP-7 is involved in the pericellular activation mechanism mediated by CD151, a crucial step in proteolysis on the cell membranes under various pathophysiological conditions including cancer invasion and metastasis.
Similar content being viewed by others

Main
Matrix metalloproteinases (MMPs) are zinc-dependent endopeptidases capable of digesting extracellular matrix (ECM) and non-ECM molecules, and comprise a gene family of 23 different members in humans.1, 2 They are synthesized as proenzymes consisting of the three basic domains, that is, propeptide (PP), zinc-binding catalytic and COOH-terminal hemopexin-like domains. MMPs can be classified into secreted-type MMPs and membrane-type MMPs (MT-MMPs), and the latter possess a transmembrane region or a glycosylphosphatidylinositol-anchored tail next to the hemopexin-like domain.1, 2 Since MMPs are expressed in various types of cells and are capable of degrading almost all ECM components in tissues, they are believed to play a key role in the processes of physiological tissue remodeling and excessive ECM breakdown under pathological conditions including cancer invasion and metastases.1 Among the MMP members, MMP-7 (also known as matrilysin-1) is unique in its minimum MMP structure, its broad range of substrate specificity and its restricted production by normal mucosal and exocrine gland epithelial cells, as well as by carcinoma cells.3 Under physiological conditions, MMP-7 is secreted on the apical site of the epithelium and in the lumenal direction of the glands4, 5 and activates defensin, which functions as an antibacterial peptide.6 Thus, these data suggest that MMP-7 is involved in maintaining the patentability of the glandular lumen by digesting unknown non-ECM proteins in the lumen4 and/or defending bacterial infections by activating defensin.6 On the other hand, MMP-7 is overexpressed in human adenoma cells of the breast and colon,7, 8 as well as carcinoma cells in the human gastrointestinal, endometrial and lung adenocarcinomas.9, 10, 11, 12 Thus, MMP-7 is considered to contribute to early tumor development during carcinogenesis.7 However, since the expression and/or activation of MMP-7 zymogen (proMMP-7) correlates well with invasion and metastases in adenocarcinomas of the stomach, colon and endometrium,11, 13, 14 it is likely that MMP-7 is also implicated in cancer progression. Importantly, there is a large difference in the distribution of MMP-7 between adenomas and adenocarcinomas; MMP-7 is predominantly immunolocalized to the apical surface of dysplastic glands of the adenomas,7 whereas the cytoplasm of carcinoma cells at the invasive front is diffusely immunostained in human gastrointestinal and endometrial carcinomas.11, 14 Thus, the data, together with the broad range of substrate specificity of MMP-7,3, 15 suggest the possibility that different functions of this proteinase under various pathophysiological conditions may be dictated by vectorial secretion of MMP-7. Yu et al16 have demonstrated that CD44 heparan sulfate proteoglycan (CD44HSPG), which is selectively expressed on the apical site of gland cells, anchors active MMP-7 to the lumenal site of postpartum uterine and lactating mammary gland epithelium in normal mice, but that in CD44−/− mice MMP-7 is redistributed from the apical to the basal compartment of the epithelia. However, no studies have elucidated the molecular mechanism regulating MMP-7 localization to the basal epithelial compartment or the depolarized diffuse cytoplasmic distribution of MMP-7 in adenocarcinoma cells at the invasive front. Similar to other MMPs such as MMP-2, the activated form of MMP-7 is readily detected within both carcinoma11, 17 and normal intestinal tissues.6 Our previous in vitro biochemical studies have shown that proMMP-7 is fully activated by treatment with trypsin or MMP-3, and partially with plasmin or leukocyte elastase.15 However, little is known about the localization of proMMP-7 activation and the molecules involved in the activation processes.
CD151 (also known as PETA-3 or SFA-1) is a member of the transmembrane 4 superfamily (TM4SF) that is comprised of four transmembrane domains, two extracellular and one intracellular loops, and NH2- and COOH-terminal intracytoplasmic domains,18, 19 This molecule can form a complex with α3β1 integrin through the interaction of the extracellular loop of CD151 and the stalk region of α3 integrin.20, 21 Other integrins (α5β1, α6β1, α6β4 and αIIbβ3) are also known to interact with CD151 depending on cell types,22, 23, 24, 25 although information about the binding sites is not available. Since CD151 is located at the periphery of the thin invasive protrusions (invadopodia) of cancer cells and treatment of the cells with anti-CD151 monoclonal antibody inhibits metastasis,26 this molecule is considered to play an important role in invasion and metastasis. It has also been reported that the prognosis of patients with lung carcinoma is poorer in CD151-positive patients than in CD151-negative patients.27 However, limited information is available concerning the mechanisms of CD151-mediated metastasis or interaction of CD151 with proteins other than integrins.
In the present study, we sought the binding proteins to proMMP-7 by screening human lung cDNA library using a yeast two-hybrid system, and identified CD151. The data demonstrate that proMMP-7 is captured and activated on the cell membranes through interaction with CD151 in the presence of its substrate, and suggest the possibility that this pericellular activation of proMMP-7 may be involved in CD151-mediated progression of human cancers.
Materials and methods
Cell Lines and Antibodies
COS7 cells and CaR-1 human rectal carcinoma cells15 were maintained in Dulbecco's modified Eagle's medium (DMEM; Invitrogen Corp., Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Tissue Culture Biologicals, Tulare, CA, USA) and antibiotics. The following antibodies and nonimmune immunoglobulin G (IgG) were used in this study: anti-MMP-7 mouse monoclonal antibody (125-20H11; Daiichi Fine Chemical Co., Ltd, Takaoka, Japan) and anti-MMP-7 rabbit polyclonal antibodies (M8683; Sigma-Aldrich Co., St Louis, MO, USA), both of which recognize precursor and active forms of MMP-7; anti-CD151 mouse monoclonal antibodies (14A2.H1, BD Biosciences PharMingen, San Diego, CA, USA, and 11G5a, Serotec Ltd, Oxford, UK) reacting with the extracellular loop of CD151; anti-MMP-3 mouse monoclonal antibody (55-2A4; Daiichi Fine Chemical Co., Ltd), which reacts with precursor and active forms of MMP-3; negative control mouse IgG1κ (BD Biosciences PharMingen), nonimmune mouse IgG (DakoCytomation Denmark A/S, Glostrup, Denmark) and rabbit nonimmune IgG (DakoCytomation Denmark A/S).
Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
RT-PCR was used to examine the expression of CD151 and MMP-7 in CaR-1 cells, COS7 cells, mock transfectants and CD151 stable transfectants. Total RNA was isolated from the cells using ISOGEN (NIPPON GENE Co., Ltd, Tokyo, Japan) and reverse-transcribed with oligo-d(T)Primer (BD Biosciences Clontech, Palo Alto, CA, USA) and SuperScript II reverse transcriptase (Invitrogen Corp.). An aliquot of the reaction mixture was subjected to polymerase chain reaction (PCR) for amplification of CD151, MMP-7 and β-actin using 5 U TaKaLa Ex Taq™ DNA polymerase (Takara Shuzo Co., Ltd, Shiga, Japan). The primers used for the amplification were 5′-AAC ACG GAG CTC AAG GAG AA-3′ (forward primer) and 5′-AGA TGT TGG AGG CAT GGT CT-3′ (reverse primer) for CD151, 5′-GGT CAC CTA CAG GAT CGT ATC TAT-3′ (forward primer) and 5′-CAT CAC TGC ATT AGG ATC AGA GGA A-3′ (reverse primer) for MMP-7, and 5′-TGA CGG GGT CAC CCA CAC TGT GCC CAT CTA-3′ (forward primer) and 5′-CTA GAA GCA TTT GCG GTG GAC GAT GGA GGG-3′ (reverse primer) for β-actin. The nucleotide sequence of the amplified fragments was confirmed by cycle sequencing using a DYEnamic™ ET dye terminator cycle sequencing kit and MegaBASE™ 1000 DNA sequencer (Amersham Biosciences Corp., Piscataway, NJ, USA).
Yeast Two-Hybrid Screening
The cDNA encoding full-length proMMP-7 was prepared by RT-PCR from total RNA of CaR-1 cells. The primers employed for PCR were 5′-GGA ATT CCT GCC GCT GCC TCA GGA G-3′ (forward primer) and 5′-ACG CGT CGA CTT TCT TTC TTG AAT TAC TTC TC-3′ (reverse primer), which correspond to the NH2- and COOH-terminal regions of proMMP-7 incorporating EcoRI and SalI sites, respectively. The proMMP-7 cDNA was cloned into pAS2-1 (BD Biosciences Clontech), generating pAS2-proMMP-7 vector. To confirm the sequence of the insert, the vector was analyzed using the DNA sequencer as described above. Human lung cDNA library cloned in yeast activation domain plasmid pACT2 (BD Biosciences Clontech) was screened by cotransforming into Saccharomyces cerevisiae strain Y-190 (BD Biosciences Clontech) with pAS2-proMMP-7 vector according to the manufacturer's instructions. Positive clones were identified by growth on media lacking leucine (−Leu)/tryptophan (−Trp)/histidine (−His), but containing 15 mM 3-amino-1,2,4-triazole (+3AT). Positive clones grown on the −Leu/−Trp/−His/+3AT medium were further screened by X-α-galactosidase (X-α-gal) assay. 5-Bromo-4-chloro-3-indolyl-α-D-galactopyranoside (BD Biosciences Clontech) was spread on −Leu/−Trp/−His/+3AT medium to make indicator plates. Colonies were streaked out on the indicator plates and allowed to grow for a few days. Candidate colonies were selected by formation of blue colonies. This streaking step was repeated twice more. The plasmids isolated from yeast were used to transform Escherichia coli strain HB101 by electroporation. The cells harboring target plasmids were isolated by plating on M9 agar plates containing a mixture of amino acids except for leucine, 50 μg/ml ampicilline, 40 μg/ml proline and 1 mM thiamine hydrochloride. Inserts of each independent positive clone were sequenced using the DNA sequencer. A homology search was performed using BLAST2: Sequence Similarity Search served by GenomeNet (http://www.blast.genome.ad.jp/).
Immunoblotting (IB) and Immunoprecipitation
To examine the production of proMMP-7 by CaR-1 cells, COS7 cells, mock transfectants and CD151 stable transfectants, culture media harvested 2 days after culturing in serum-free DMEM were subjected to IB for proMMP-7. Proteins in the concentrated culture media (1 ml/lane) were electrophoresed on 12.5% SDS-PAGE under reducing conditions, and then electrotransferred onto polyvinylidene difluoride membranes (ATTO Corporation, Tokyo, Japan). The membranes were reacted with anti-MMP-7 antibody (10 μg/ml; 125-20H11) at 4°C for 12 h after blocking nonspecific reaction with 3% bovine serum albumin (BSA) in phosphate-buffered saline (PBS), and incubated with horseradish peroxidase-labeled secondary antibody (Vector Laboratories Inc., Burlingame, CA, USA). Immunoreactive bands were detected with ECL reagents (Amersham Bioscience Co.) using Lumivision HSII (Taitec Co. Ltd, Tokyo, Japan). For detection of CD151 in these cells, the cell lysates were prepared in a lysis buffer of 10 mM Tris-HCl buffer, pH 7.5, containing 0.5% NP-40 (IGEPAL CA-630; Sigma-Aldrich Co.), 1 mM EDTA, 150 mM NaCl and proteinase inhibitor cocktail (Complete™ Mini; Roche Diagnostics GmbH, Mannheim, Germany), and insoluble debris was removed by centrifugation. The lysates (2.0 × 107 cells/lane) were subjected to immunoprecipitation with anti-CD151 antibody (4 μg/ml; 14A2.H1), followed by incubation with Protein G Sepharose 4 Fast Flow beads (Amersham Biosciences Corp.) after preclearing with the beads according to our previous method.28 Beads carrying immune complexes were washed four times in the lysis buffer and boiled for 5 min in SDS reduction buffer. The immunoprecipitates were separated on 12.5% SDS-PAGE under reducing conditions. The proteins electrotransferred onto polyvinylidene difluoride membranes were immunoblotted with 125I-labeled anti-CD151 antibody (10 μg/ml; 11G5a) at 4°C for 12 h after blocking nonspecific reaction with 3% BSA in PBS. The bound antibody was detected by an imaging plate and BAS-2000 system (Fuji Photo Film Co., Ltd, Tokyo, Japan). As for a control, the cell lysates (1 × 106 cells/lane) of CaR-1 cells, COS7 cells, mock transfectants and CD151 stable transfectants were subjected to IB for β-actin using anti-β-actin antibody (1 μg/ml; A5316; Sigma-Aldrich Co.) and horseradish peroxidase-labeled secondary antibody.
In order to show the binding between CD151 and proMMP-7, immunoprecipitation was performed with the lysates of CaR-1 cells, COS7 cells, mock transfectants and CD151 stable transfectants. Since COS7 cells and CD151 stable transfectants did not produce MMP-7 (see below), these cells were incubated in the presence of proMMP-7 (1 μg/ml) for 2 h prior to immunoprecipitation. The cell lysates were prepared and subjected to immunoprecipitation with anti-CD151 antibody (4 μg/ml; 14A2.H1) or non-immune mouse IgG as described above. For some experiments, the cell lysates were incubated for 6 h at 4°C with active MMP-7 (0.5 μg/ml), which was prepared by activation of proMMP-7 as described below, and then subjected to immunoprecipitation. The immunoprecipitates were separated on 12.5% SDS-PAGE and proteins electrotransferred onto polyvinylidene difluoride membranes were incubated with 125I-labeled anti-MMP-7 antibody (10 μg/ml; 125-20H11) or 125I-labeled anti-CD151 antibody (10 μg/ml; 11G5a). As for a control, the cell lysates were incubated with proMMP-3 (2 μg/ml) and then subjected to immunoprecipitation using anti-CD151 antibody or nonimmune mouse IgG in a similar way. The immunoprecipitated proteins transferred on the membranes were immunoblotted with 125I-labeled anti-MMP-3 antibody (10 μg/ml; 55-2A4) or anti-CD151 antibody (10 μg/ml; 11G5a). The bound antibodies were detected by an imaging plate and BAS-2000 system (Fuji Photo Film Co., Ltd).
Yeast Two-Hybrid Assays to Examine Interaction Domains
The cDNA encoding full-length CD151 was prepared from human lung cDNA library by PCR using a set of primers 5′-CGG AAT TCA GGA TGG GTG AGT TCA ACG-3′ (forward primer) and 5′-ACG CGT CGA CTC AGT AGT GCT CCA GC-3′ (reverse primer), which correspond to the NH2- and COOH-terminal intracytoplasmic regions of CD151 with EcoRI and SalI sites, respectively. The cDNAs encoding deletion mutants of proMMP-7 and CD151 were generated from the full-length cDNAs by PCR using the corresponding primers (Table 1), and cloned into pAS2 or pACT2 vector (BD Biosciences Clontech). Sequences of the cDNAs were confirmed by cycle sequencing using the DNA sequencer. The resulting plasmids were designated as shown in Figure 1. pAS2-proMMP-7, pAS2-PP and pAS2-MMP-7 correspond to plasmids containing the cDNAs encoding full-length proMMP-7, PP of proMMP-7 and active MMP-7, respectively. On the other hand, pACT2-CD151, pACT2-C1, pACT2-EL1, pACT2-CL, pACT2-EL2 and pACT2-C2 indicate plasmids with the cDNAs encoding full-length CD151, NH2-terminal intracytoplasmic region, NH2-terminal extracellular loop, intracytoplasmic loop, COOH-terminal extracellular loop and COOH-terminal intracytoplasmic region, respectively. These plasmids for MMP-7 and CD151 species were cointroduced into yeast strain Y-187 (BD Biosciences Clontech) by the lithium acetate method.29 Positive clones were identified by growth on -Leu/-Trp/-His medium and X-α-gal assay.

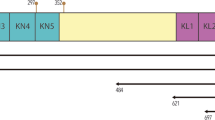
Schematic diagram of the inserted cDNA fragments used for yeast two-hybrid assays. Yeast plasmids were prepared by inserting cDNAs for full-length or deleted proMMP-7 into GAL4 DNA-BD vector and cDNAs for full-length or deleted CD151 into GAL4 AD vector. ProMMP-7 was divided to the PP and metalloproteinase domains. CD151 was split into five domains according to its structural features; the NH2-terminal intracytoplasmic region (C1), NH2-terminal extracellular loop (EL1), intracytoplasmic loop (CL), COOH-terminal extracellular loop (EL2) and COOH-terminal intracytoplasmic region (C2).
Establishment of Stable Transfectants Expressing CD151
The cDNA encoding full-length CD151 was inserted to pCI-neo mammalian expression vector (Promega Corporation, Madison, WI, USA), generating pCI-CD151 vector. Since COS7 cells express neither MMP-7 nor CD151 as reported previously30 and as confirmed by RT-PCR and IB in the present study (Figure 2a, lane 2), we used the cells to establish stable transfectants for CD151. COS7 cells were cultured in DMEM supplemented with 10% FBS in 10-cm dishes and transfected with 7.5 μg of pCI-CD151 vector or the original pCI-neo vector by DOTAP liposomal transfection (Roche Diagnostics GmbH) according to the manufacturer's instructions. Transfected cells were cultured in the presence of 300 μg/ml G418 (Invitrogen Corp.), and clones were established from the resistant cells. Stable transfectants were isolated and expression levels of CD151 were evaluated by flow cytometry.

Expression of proMMP-7 and CD151 in CaR-1 cells and CD151 stable transfectants and demonstration of the interaction between proMMP-7 and CD151. (a) Expression of proMMP-7 and CD151 in CaR-1 cells (lane 1), parental COS7 cells (lane 2), mock transfectants (lane 3) and stable transfectants with high expression of CD151 (clone 16; lane 4). The expression at the mRNA and protein levels in these cells was examined by RT-PCR and IB as described in Materials and methods. β-Actin was used as for a control. (b) Coimmunoprecipitation of proMMP-7 and CD151 from CaR-1 cells. Lysates of CaR-1 cells were subjected to immunoprecipitation with nonimmune mouse IgG (NI; lanes 1 and 3) or anti-CD151 monoclonal antibody (14A2.H1; lanes 2 and 4), and then immunoblotted with 125I-labeled anti-MMP-7 antibody (125-20H1; lanes 1 and 2) or 125I-labeled anti-CD151 antibody (14A2.H1; lanes 3 and 4) as described in Materials and methods. Note that proMMP-7 of 29 kDa (arrowhead) is coimmunoprecipitated with CD151 of 30 kDa (arrow). (c) Coimmunoprecipitation of proMMP-7 and CD151 from CD151 stable transfectants (clone 16) preincubated with proMMP-7. Cell lysates were subjected to immunoprecipitation with nonimmune mouse IgG (lanes 1 and 3) or anti-CD151 monoclonal antibody (lanes 2 and 4), and then immunoblotted with anti-MMP-7 antibody (lanes 1 and 2) or anti-CD151 antibody (lanes 3 and 4) as described above. Arrowhead and arrow indicate proMMP-7 and CD151, respectively. (d) Immunoprecipitation of proMMP-7 and CD151 in mock transfectants preincubated with proMMP-7. Cell lysates of mock transfectants were subjected to immunoprecipitation and subsequent IB as described above. Note that negligible bands are observed.
Flow Cytometry
Stable transfectants were dissociated from dishes by incubating with 0.25% trypsin and 0.02% EDTA (Immuno-Biological Laboratories Co., Ltd, Gumma, Japan) for 3 min at 37°C and suspended in PBS containing 1% FBS after blocking the activity of trypsin with 10% FBS. They were then incubated on ice for 30 min with anti-CD151 monoclonal antibody (14A2.H1; 1 μg/106 cells) or nonimmune mouse IgG1 κ (1 μg/106 cells). After being washed, the cells were incubated with fluorescein isothiocyanate (FITC)-conjugated F(ab′)2 of rabbit anti-mouse IgG (DakoCytomation Denmark A/S), and then analyzed by Courter EPICS XL-MCL digital flow cytometer (Beckman Courter, Inc., Fullerton, CA, USA).
Laser-Scanning Confocal Microscopy
For immunohistochemical staining, parental COS7 cells, mock transfectants and CD151 stable transfectants were cultured on Lab-Tek chamber slides (Nalge Nunc International, Rochester, NY) for 3 days and fixed for 5 min with 4% paraformaldehyde in PBS. After blocking nonspecific binding with 3% normal goat serum in PBS, cells were incubated at room temperature for 1 h with anti-CD151 monoclonal antibody (11G5a; 10 μg/ml), anti-MMP-7 rabbit polyclonal antibodies (M8683; 10 μg/ml), nonimmune mouse IgG (10 μg/ml) or nonimmune rabbit IgG (10 μg/ml). They were then reacted with FITC- or tetramethylrhodamine (TRITC)-conjugated secondary antibodies (DakoCytomation Denmark A/S) following a wash with PBS. For double immunohistochemical staining of CD151 and MMP-7, CD151 stable transfectants and CaR-1 cells were cultured on Lab-Tek chamber slides. After blocking nonspecific binding, they were incubated at room temperature for 1 h with anti-CD151 monoclonal antibody (11G5a; 10 μg/ml) and anti-MMP-7 rabbit polyclonal antibodies (M8683; 10 μg/ml), or nonimmune mouse IgG (10 μg/ml) and nonimmune rabbit IgG (10 μg/ml). They were then fixed and reacted with FITC- or TRITC-conjugated secondary antibodies as described above. Coverslips were mounted with fluorescent mounting medium (DakoCytomation California Inc., Carpinteria, CA, USA), and immunofluorescence was examined using a laser-scanning confocal microscope, Fluoview FV300 (Olympus Co., Ltd, Tokyo, Japan).
Purification of ProMMP-7 and ProMMP-3 and Iodination of Proteins
ProMMP-7 was purified from culture media of CaR-1 cells, and active MMP-7 was prepared by incubation of proMMP-7 with 1 mM aminophenylmercuric acetate for 4 h at 37°C according to our previous methods.15 ProMMP-3 was purified from culture media of rheumatoid synovial fibroblasts as described previously.31 ProMMP-7, active MMP-7, proMMP-3, anti-MMP-7 antibody (125-20H11) and anti-CD151 antibody (11G5a) were iodinated using IODOGEN® reagent (Pierece Chemical Company, Rockford, IL, USA).32
Binding Assay
Parental COS7 cells, mock transfectants and CD151 stable transfectants were cultured in DMEM supplemented with 10% FBS in 24-well dishes, washed twice with PBS and incubated with serum-free media containing 5% BSA at 37°C for 30 min. The media were replaced with PBS containing 125I-labeled proMMP-7 (1.8 μg/ml), 125I-labeled active MMP-7 (1.8 μg/ml) or 125I-labeled proMMP-3 (3 μg/ml) and incubated at 4°C for 1 h. The media were then harvested and cells were washed twice with PBS. These media and washes were combined as the unbound fractions. The cells were then lysed with 0.1 N NaOH and washed twice with PBS. The cell lysates and washes were combined as the bound fractions. Radioactivity of the bound and unbound fractions was counted by γ-counter, ARC-600 (ALOKA Co., Ltd, Tokyo, Japan). Inhibition experiments were performed by incubation of stable transfectants with nonlabeled proMMP-7 (0, 5 or 10 μg/ml), anti-CD151 antibody (10 μg/ml; 14A2.H1) or nonimmune mouse IgG (10 μg/ml) for 1 h at 4°C prior to the binding assay.
In Situ Zymography Using Crosslinked Carboxymethylated Transferrin (CCm-Tf) Films
CD151 stable transfectants and CaR-1 cells maintained in DMEM supplemented with 10% FBS were detached with 0.25% trypsin and 0.02% EDTA, and the activity of trypsin was completely blocked with 10% FBS in DMEM. After suspending the cells with serum-free DMEM containing 0.2% lactalbumin hydrolysate (Invitrogen Corp.), they were seeded on CCm-Tf films which were coated with CCm-Tf (2.5 μm thickness) on polyethylene telephthalate support (Fuji Photo Film Co., Ltd) according to our methods.33 Cells on the films were incubated in a 5% CO2 incubator for 6–16 h at 37°C, and then stained with 0.5% Biebrich Scarlet (Sigma-Aldrich Co.) and hematoxylin after being air-dried. For some experiments, the cells were also incubated on CCm-Tf films coated with 0.1 M o-phenanthroline or on the films in the presence of 20 μg/ml recombinant tissue inhibitor of metalloproteinases 1 (rTIMP-1),34 anti-MMP-7 antibody (M8683; 25 μg/ml), anti-CD151 antibody (14A2.H1; 25 μg/ml), nonimmune mouse IgG (25 μg/ml) or nonimmune rabbit IgG (25 μg/ml). Some of these cells were lysed in a lysis buffer and analyzed by IB with 125I-labeled anti-MMP-7 antibody (10 μg/ml; 125-20H11) as described above.
Immunohistochemistry and In Situ Zymography in Human Lung Adenocarcinoma and Normal Lung Tissues
Fresh carcinoma tissues were obtained from 10 patients with lung adenocarcinoma who underwent surgery at the Keio University Hospital, Tokyo, Japan. For immunohistochemical staining, carcinoma tissues and normal lung tissues remote from the tumor were snap-frozen in liquid nitrogen and the 4-μm sections were made with a cryostat microtome. In addition, normal bronchial tissues (three cases) were obtained at autopsy and frozen sections were prepared in a similar way. The sections were fixed for 5 min with 4% paraformaldehyde, and then incubated at room temperature for 1 h with anti-CD151 monoclonal antibody (11G5a; 10 μg/ml) and anti-MMP-7 rabbit polyclonal antibodies (M8683; 10 μg/ml), or nonimmune mouse IgG (10 μg/ml) and nonimmune rabbit IgG (10 μg/ml) after blocking nonspecific binding with 3% normal goat serum in PBS. They were then reacted with FITC- or TRITC-conjugated secondary antibodies as described above. Coverslips were mounted with fluorescent mounting medium, and immunofluorescence was examined using a laser-scanning confocal microscope, Fluoview FV300. The tissue sections were also observed by a phase-contrast microscope, CKX41 (Olympus Co. Ltd) with CCD camera (VB-7000, Keyence Corp., Osaka, Japan). For in situ zymography, serial frozen sections of the adenocarcinoma and normal lung and bronchial tissues were placed on CCm-Tf films or CCm-Tf films coated with 0.1 M o-phenanthroline, incubated for 10 h at 37°C, and then stained with 0.5% Biebrich Scarlet (Sigma-Aldrich Co.) and hematoxylin as described above. Serial sections were also stained with hematoxylin and eosin. In some experiments, the lung carcinoma tissues were sliced at 5-mm thickness and incubated with anti-CD151 antibody (14A2.H1; 25 μg/ml), anti-MMP-7 antibody (M8683; 25 μg/ml), nonimmune mouse IgG (25 μg/ml) or nonimmune rabbit IgG (25 μg/ml) in DMEM for 1 h at 4°C. The tissues were snap-frozen and frozen sections were placed on CCm-Tf films. After incubation for 10 h at 37°C, they were stained as mentioned above. For experimental use of the surgical specimens, the informed consent was obtained from the patients according to the hospital ethical guidelines.
Results
Screening of Proteins that Interact with ProMMP-7
To identify the proteins that interact with proMMP-7, 1 × 107 independent clones from the human lung cDNA library were screened by the yeast two-hybrid system using a cDNA encoding proMMP-7 as bait. On the basis of both nutritional and α-galactosidase activity, we evaluated five positive clones and sequenced (data not shown). Since they included human CD151, which is a membrane protein belonging to TM4SF, we focused on this molecule as a candidate-binding protein to proMMP-7.
Coimmunoprecipitation of ProMMP-7 and CD151
Since CaR-1 cells expressed both proMMP-7 and CD151 at the mRNA and protein levels (Figure 2a, lane 1), we first studied the interaction between proMMP-7 and CD151 by immunoprecipitation using CaR-1 cells. As shown in Figure 2b, proMMP-7 of 29 kDa and CD151 of 30 kDa were coimmunoprecipitated from the CaR-1 cell lysates with anti-CD151 antibody, while negligible immunoprecipitation was observed with nonimmune mouse IgG. Similarly, when CD151 stable transfectants, which expressed only CD151 (Figure 2a, lane 4), were subjected to immunoprecipitation with anti-CD151 antibody, coimmunoprecipitation of proMMP-7 and CD151 was found only after preincubation of the cells with exogenously added proMMP-7 (Figure 2c). Parental COS7 cells and mock transfectants showed no immunoprecipitation under similar conditions (Figure 2d for COS7 cells and data not shown for mock transfectants). On the other hand, no active MMP-7 species of 19 kDa was immunoprecipitated from the lysates of CaR-1 cells (Figure 2b) or CD151 stable transfectants that were preincubated with or without active MMP-7 prior to immunoprecipitation with anti-CD151 antibody (data not shown). In addition, proMMP-3 was not immunoprecipitated from the CaR-1 cell lysates that were preincubated with proMMP-3 (data not shown).
Interaction of ProMMP-7 with COOH-Terminal Extracellular Loop of CD151 by Yeast Two-Hybrid Assays
In order to examine the binding regions between proMMP-7 and CD151, yeast two-hybrid assays were performed by cointroducing plasmid DNAs encoding the full-length molecules and their deletion mutants into yeast (Figure 1). Among the yeast clones examined, the clone cotransformed with pAS2-proMMP-7 expressing proMMP-7 and pACT2-CD151 expressing full-length CD151 and the clone cotransformed with pAS2-PP expressing proMMP-7 PP and pACT2-CD151 grew on −Trp/−Leu/−His medium, although the combination of pAS2-MMP-7 expressing active MMP-7 and pACT2-CD151 produced no colonies (Figure 3a). On the other hand, using proMMP-7 as bait, clones expressing full-length CD151 (pACT2-CD151) or the COOH-terminal extracellular loop of CD151 (pACT2-EL2) formed blue-colored colonies on the medium (Figure 3b). However, the clones cotransformed by other combinations showed none or only a few white colonies as background growth (Figure 3b). In addition, the clone cotransformed with pAS2 PP and pACT2-EL2 could grow on the medium, but the combinations of proMMP-7 PP and each CD151 domain other than EL2 had only negligible colonies (data not shown). Thus, these data show that the PP of proMMP-7 interacts with the COOH-terminal extracellular loop of CD151.

Yeast two-hybrid assays showing interaction between the PP domain of proMMP-7 and the COOH-terminal extracellular loop domain of CD151. (a) Yeast strain Y-187 was cotransformed with prey plasmids encoding proMMP-7 (ProMMP-7), PP of proMMP-7 propeptide or active MMP-7 (Active MMP-7) in conjunction with bait plasmid encoding full-length CD151 (CD151-whole). As for controls, yeast was transformed with the vector encoding proMMP-7, PP of proMMP-7 or CD151-whole alone. Yeast transformants were assayed for bait–prey interactions by determining their ability to grow on –Leu/–Try/–His medium and to produce β-galactosidase as described in Materials and methods. (b) Yeast strain was cotransformed with prey plasmids encoding full-length CD151 (CD151-whole), NH2-terminal intracytoplasmic region (CD151-C1), NH2-terminal extracellular loop (CD151-EL1), intracytoplasmic loop (CD151-CL), COOH-terminal extracellular loop (CD151-EL2) or COOH-terminal intracytoplasmic region (CD151-C2) with plasmids for full-length proMMP-7 (ProMMP-7). Yeast transformants were assayed for bait–prey interactions as described above.
Establishment of Stable Transfectants for CD151 and Binding Assay
By transfection of COS7 cells with pCI-CD151 vector encoding human CD151, 60 stable transfectants were established. When the expression levels of CD151 in these transfectants were evaluated by flow cytometry, they showed a wide range of expression. Thus, we selected three clones with low expression (clones 2, 3 and 4) and three with high expression (clones 6, 16 and 17) for further study (Figure 4a). Using these CD151 stable transfectants with low or high expression of CD151, mock transfectants and parental COS7 cells, binding activity of 125I-labeled proMMP-7 was examined. As shown in Figure 4b, the binding capacity of 125I-proMMP-7 on the transfectants with high expression was significantly increased as compared with parental COS7 cells, mock transfectants or transfectants with low expression. The binding of 125I-proMMP-7 to CD151 stable transfectants was competitively inhibited with nonlabeled proMMP-7 in a dose-dependent manner (Figure 4c). In addition, similar inhibition was obtained by incubation of the transfectants with anti-CD151 antibody, but not with nonimmune mouse IgG prior to the binding assay (Figure 4c). On the other hand, only negligible background binding of 125I-labeled active MMP-7 or 125I-labeled proMMP-3 was observed with transfectants with high or low CD151 expression, mock transfectants or parental COS7 cells (data not shown).

Flow cytometry of CD151 stable transfectants and binding activity of proMMP-7 to CD151 stable transfectants. (a) Representative data of flow cytometrical analysis of parental COS7 cells and CD151 stable transfectants with low (Cl 2) or high expression (Cl 16). Solid and dotted lines indicate signals detected by anti-CD151 antibody and nonimmune IgG, respectively. Cl, clone. (b) Binding of proMMP-7 to COS7 cells, mock transfectants and CD151 stable transfectants. 125I-labeled proMMP-7 was added to parental COS7 cells, mock transfectants (data not shown) and stable transfectants with low (Cl 2, Cl 3 and Cl 4) or high expression of CD151 (Cl 6, Cl 16 and Cl 17), incubated for 1 h at 4°C, and relative binding ratios of proMMP-7 were calculated as described in Materials and methods. Bars indicate the mean±s.d. from three experiments. Cl, clone. *P<0.05; **P<0.01. (c) Inhibition of 125I-labeled proMMP-7 binding to CD151 stable transfectants by treatment with cold proMMP-7 and anti-CD151 antibody. Nonlabeled cold proMMP-7, nonimmune IgG (NI) or anti-CD151 antibody was added to transfectants with high expression of CD151 (clone 16) prior to the binding assay, and relative binding ratios were calculated as described in Materials and methods. Bars indicate the mean±s.d. from three experiments. *P<0.05; **P<0.01.
Colocalization of CD151 and ProMMP-7 on Cell Membranes by Laser-Scanning Confocal Microscopy
When CD151 was immunolocalized in COS7 cells, mock transfectants and CD151 stable transfectants with anti-CD151 antibody, it was localized to the cell membranes of only the stable transfectants with high expression of CD151 (data not shown). Thus, the stable transfectants with high expression were incubated with exogenous proMMP-7, and double immunostaining for CD151 and proMMP-7 was performed. As shown in Figure 5, both CD151 and proMMP-7 were localized to the cell membranes in a dotted pattern (Figure 5a and b, respectively), and a merged image of the double immunostaining showed yellow dots on the membranes (Figure 5c), suggesting that most of the two molecules coexist on the cell membranes. Immunostaining with nonimmune IgG showed no signals (Figure 5d). Similarly, when CaR-1 cells which express both CD151 and proMMP-7 were subjected to double immunostaining, CD151 and proMMP-7 were mostly colocalized on the cell membranes in a dotted pattern (Figure 5e–h).

Double immunostaining of CD151 and proMMP-7 on the cell membranes of CD151 stable transfectants and CaR-1 cells. CD151 stable transfectants (clone 16) (a–d) and CaR-1 cells (e–h) were reacted with anti-CD151 monoclonal antibody (11G5a) and anti-MMP-7 rabbit polyclonal antibodies (M8683) or nonimmune mouse IgG and nonimmune rabbit IgG prior to fixation, incubated with FITC- or TRITC-conjugated secondary antibodies after fixation, and observed by a laser-scanning confocal microscope as described in Materials and methods. (a, e), localization of CD151; (b, f), localization of MMP-7; (c, g), the merged image; (d, h), immunostaining with nonimmune IgG. Bar, 10 μm.
Detection of MMP-7 Activity around Cell Membranes of CD151-Expressing Cells
In order to examine the proteolytic activity of MMP-7, in situ zymography using polyethylene films coated with CCm-Tf, a substrate of MMP-7, was performed. When parental COS7 cells, which expressed neither proMMP-7 nor CD151, were incubated on CCm-Tf films, they showed no or only negligible proteolytic activity against CCm-Tf in the presence and absence of exogenous proMMP-7 (Figure 6a and b). In contrast, stable transfectants with high expression of CD151 digested pericellular CCm-Tf substrate only when proMMP-7 was added to the cells incubated on the films (Figure 6c and d). CaR-1 cells, which express both proMMP-7 and CD151, exhibited strong diffuse proteolytic activity on and around the cells when cultured on the CCm-Tf films (Figure 7a). This activity was almost completely abolished when the cells were incubated on CCm-Tf films coated with o-phenanthroline (Figure 7b). The activity was also blocked with rTIMP-1, a specific MMP inhibitor (Figure 7c). In addition, negligible activity was detected when anti-MMP-7 antibody was present in the incubation (Figure 7d). Importantly, treatment of the cells with anti-CD151 antibody also blocked the proteolytic activity (Figure 7e), although nonimmune IgG had no such effects (Figure 7f). As shown in Figure 8, active MMP-7 species of 19 kDa, together with proMMP-7 of 29 kDa, was detectable in the cell lysates of CaR-1 cells incubated on CCm-Tf films, but active species was not observed when CaR-1 cells were cultured on polyethylene films without CCm-Tf, CCm-Tf films coated with o-phenanthroline or plastic dishes as we have previously reported.15

In situ zymographical detection of CCm-Tf-degrading activity on and around the CD151 stable transfectants. Parental COS7 cells (a, b) and CD151 stable transfectants with high expression (clone 16) (c, d) were subjected to in situ zymography using CCm-Tf films in the absence (a, c) or presence of proMMP-7 (b, d) as described in Materials and methods. Note that CD151 stable transfectants show pericellular CCm-Tf-degrading activity only in the presence of proMMP-7 (arrows in d). Bar, 25 μm.

Demonstration of MMP-7 activity by in situ zymography of CaR-1 cells. CaR-1 cells were subjected to in situ zymography using CCm-Tf films (a), o-phenanthroline-coated CCm-Tf films (b), CCm-Tf films in the presence of rTIMP-1 (c), anti-MMP-7 antibody (M8683) (d), anti-CD151 antibody (14A2.H1) (e) or nonimmune rabbit IgG (f) as described in Materials and methods. Bar, 25 μm.

Detection of active form of MMP-7 in lysates of CaR-1 cells incubated on CCm-Tf films. Lysates of CaR-1 cells incubated on polyethylene films without CCm-Tf coating (lane 1), CCm-Tf films coated with o-phenanthroline (lane 2) or CCm-Tf films (lane 3) were subjected to IB analysis using 125I-labeled anti-MMP-7 antibody (125-20H11) as described in Materials and methods. A mixture of purified proMMP-7 and active MMP-7 is shown as a positive control in lane 4. Note that active MMP-7 of 19 kDa (arrow) as well as proMMP-7 of 29 kDa (arrowhead) is present only in the sample incubated on CCm-Tf films (lane 3).
Colocalization of CD151 and ProMMP-7 and Detection of Metalloproteinase Activity on Human Lung Adenocarcinoma Cells
When the lung adenocarcinoma tissues were double-immunostained with antibodies against CD151 and proMMP-7, CD151 was observed predominantly on cell membranes of the carcinoma cells (Figure 9a), and proMMP-7 was localized on cell membranes and within the cytoplasm of the carcinoma cells (Figure 9b). A merged image of the immunostaining showed that these two molecules are colocalized mainly on the cell membranes as yellow dots (Figure 9c). Only negligible signals were obtained on the sections incubated with nonimmune IgG (data not shown). In normal lung tissues, CD151 was localized on pneumocytes (Figure 9d), but proMMP-7 was undetectable (Figure 9e). Similarly, CD151 was immunostained on the basolateral surface of the peribronchial gland cells, but negligible immunostaining of proMMP-7 was observed (data not shown). By in situ zymography using CCm-Tf films, no proteolytic activity was observed with the normal lung and bronchial tissues (data not shown). On the other hand, strong diffuse proteolytic activity was detected around the carcinoma cell nests (Figure 10a) and the activity was abolished with o-phenanthroline (Figure 10b), indicating that the lysis is due to metalloproteinase activity. Importantly, this activity was completely inhibited by treatment of the carcinoma tissues with anti-MMP-7 antibody (Figure 10d) and partially blocked with anti-CD151 antibody (Figure 10e), although nonimmune IgG did not affect the activity (Figure 10f). Thus, these data strongly suggest that the CCm-Tf digestion by the lung adenocarcinoma cells is due to MMP-7 activity.

Double immunohistochemistry of CD151 and proMMP-7 in human lung adenocarcinoma and normal lung tissues. For double immunostaining of CD151 and proMMP-7, frozen sections of papillary adenocarcinoma (a–c) and normal lung tissues (d–f) were reacted with anti-CD151 monoclonal antibody (11G5a) and anti-MMP-7 rabbit polyclonal antibodies (M8683), incubated with FITC- or TRITC-conjugated secondary antibodies, and observed by a laser-scanning confocal microscope or a phase-contrast microscope as described in Materials and methods. (a, d), Localization of CD151; (b, e), localization of proMMP-7; (c), the merged image of lung adenocarcinoma tissue; (f) the image by a phase-contrast microscope of normal lung tissue. Bar, 30 μm.

Detection of MMP-7 activity in human lung adenocarcinoma tissues by in situ zymography. Serial frozen sections of the lung adenocarcinoma tissues were subjected to in situ zymography using CCm-Tf films (a) or o-phenanthroline-coated CCm-Tf films (b), or stained with hematoxylin and eosin (c). The carcinoma tissues were also treated with anti-MMP-7 antibody (d), anti-CD151 antibody (e) or nonimmune rabbit IgG (f), and then subjected to in situ zymography as described in Materials and methods. Note the CCm-Tf-degrading activity around the carcinoma cell nests (negative staining in panel a), which is abolished on CCm-Tf films coated with o-phenanthroline (panel b). The activity is also almost completely and partially blocked with anti-MMP-7 antibody (d) and anti-CD151 antibody (e), respectively, but not with nonimmune IgG (f). Bar, 50 μm.
Discussion
In the present study, we identified CD151 as a potential binding protein for proMMP-7. The interaction was first screened by yeast two-hybrid system and then demonstrated by immunoprecipitation with CaR-1 cells expressing both binding partners and CD151 stable transfectants incubated with proMMP-7. The data on proMMP-7 binding assay using the CD151 stable transfectants and colocalization of proMMP-7 and CD151 on the cell membranes by double immunostaining further support the notion that proMMP-7 and CD151 bind on the cell membranes. MMPs can be classified into two major subgroups according to the structural characteristics of the molecules, that is, secreted MMPs and membrane-associated MMPs including MT-MMPs. Secreted MMPs are thought to be synthesized and secreted from cells to digest ECM and non-ECM macromolecules in the extracellular milieu, although MT-MMPs localize and function on the cell membranes.1, 2, 35 However, recent studies have indicated the possibility that secreted MMPs may also function on the cell membranes through their cell surface docking.1 Actually, some secreted MMPs are known to interact with cell membrane proteins, which include CD147 (EMMPRIN) for MMP-1,36 αvβ3 integrin and caveolin-1 for MMP-2,37, 38 CD44HSPG for MMP-7,16 and CD44 for MMP-9.39 Among these interactions, the binding between MMP-7 and CD44HSPG is different from other combinations in that MMP-7 interacts with sulfated glycosaminoglycan chains of CD44HSPG but not with its core protein,16 whereas other MMPs associate with their binding molecules through a protein–protein interaction.36, 37, 38, 39 Thus, our data in the present study demonstrate for the first time that proMMP-7 also binds to CD151 through a protein–protein interaction.
ProMMP-7 is composed of PP and catalytic domains,3 and CD151 comprises four transmembrane domains, two extracellular and one intracellular loops, and NH2- and COOH-terminal intracytoplasmic domains.18 Thus, we carried out yeast two-hybrid assays to examine which domains of the molecule interact. Our results demonstrated that colonies cotransformed with the plasmids expressing full-length CD151 and those expressing proMMP-7 or proMMP-7 PP, but not active MMP-7, grow on medium, suggesting that the PP of proMMP-7 is implicated in the interaction. This was further confirmed by the data that proMMP-7 was coimmunoprecipitated with CD151 from the cells and bound to CD151 stable transfectants, although active MMP-7 or proMMP-3 had no such characteristics. On the other hand, since colonies cotransformed with plasmids expressing proMMP-7 or proMMP-7 PP and those expressing full-length CD151 or its COOH-terminal extracellular loop were formed on medium, it is strongly suggested that the extracellular loop of CD151 is involved in the interaction. In accordance with these data, exogenously added proMMP-7 could bind to CD151 stable transfectants and both proMMP-7 and CD151 were colocalized on the cell membranes.
One of the most important findings in the present study is that proMMP-7 is activated by interaction with CD151 on the cell membranes. This was demonstrated by in situ zymography using CCm-Tf films and IB. Since treatment of the cells with anti-CD151 antibody abolished the activation, it is very likely that CD151 plays a role in the activation process. However, as CD151 has no proteolytic activity, the molecule itself is not considered an activator of proMMP-7. Our previous biochemical studies15 demonstrated that proMMP-7 is proteolytically activated fully by trypsin or MMP-3, and partially by plasmin or leukocyte elastase. However, since these proteinases are considered not to be produced by COS7 and CaR-1 cells, it is unlikely that they are responsible for the activation. On the other hand, proMMP-7 has a propensity to autoactivate and degrade itself in a concentration-dependent manner.40, 41 Thus, it may be possible to speculate that CD151 captures proMMP-7 on the cell membranes at a higher concentration, and leads to spontaneous activation of proMMP-7 through conformational changes of the proenzyme. However, the activation process in the present study required the presence of a substrate of MMP-7, that is, CCm-Tf, since no activation was observed with CaR-1 cells cultured on polyethylene films or plastic dishes without substrate coating. Thus, these data suggest that the substrates may provide the cells with the necessary conditions to facilitate activation, probably through completion of the conformational changes of proMMP-7 caused by the interaction between the PP and the extracellular loop of CD151. The reason for the different digestion patterns of the CCm-Tf substrate by CD151 stable transfectants (a localized pericellular footprint pattern) and CaR-1 cells (a strong diffuse digestion pattern) is not clear at this point. However, it is reasonable to speculate that CaR-1 cells continuously secrete and simultaneously activate proMMP-7 in the presence of its substrate, probably because they are endowed with more efficient machinery for proMMP-7 activation through interaction with CD151 and/or activator. Although our data provide the evidence of pericellular activation of proMMP-7, further studies are needed to understand the precise mechanism of CD151-mediated proMMP-7 activation.
Previous studies have reported the vectorial secretion of MMP-7. In normal mucosal and gland epithelium and in polarized monolayer cultures, MMP-7 is preferentially secreted to the apical surface.3, 5 Yu et al16 demonstrated that CD44HSPG, which is expressed predominantly in the apical surface of glands, functions as a docking molecule for MMP-7 and plays a key role in the localization of MMP-7 to the lumenal epithelial surface of the uterine and mammary glands. The authors also showed that, in CD44−/− mice, MMP-7 is redistributed from the apical to the basal compartment of the uterine endometrial gland cells and mammary gland epithelial cells. Although they speculated that in such mice, MMP-7 may bind to other HSPG molecules expressed on the basal surface of epithelial cells and in the basement membrane,16 no study was performed to elucidate which molecule determined MMP-7 localization to the basolateral compartment. On the other hand, localization and expression of MMP-7 protein in the basolateral compartment of epithelial cells are reported in various pathological conditions. They include epithelial cells migrating over injured airway,5 rat corneal squamous epithelium during wound healing after excimer keratectomy42 and carcinoma cells in human cancer tissues such as colon adenocarcinomas,14 infiltrative basal cell carcinomas and squamous cell carcinomas of the skin.43 CD151 is localized to basal cells of stratified squamous cell epithelium22, 24, 44, 45 and the basolateral layer of enterocytes on villi.22 Although information about localization in carcinoma cells is limited, our immunohistochemical and in situ zymographical studies on the human lung adenocarcinoma tissues have demonstrated colocalization of CD151 and MMP-7 on the carcinoma cell membranes and MMP-7 activity at the carcinoma cell nests. Thus, it is tempting to speculate that CD151 may function as a possible docking molecule for proMMP-7 activation in such cells under pathological conditions.
Several lines of evidence have indicated that CD151 is implicated in invasion and metastasis. Transfection of CD151 cDNA into cancer cell lines enhances invasion of Matrigel, cell motility and pulmonary metastasis in an experimental animal model46 and, conversely, treatment of CD151-overexpressing cancer cells with anti-CD151 antibodies markedly inhibits cell migration and metastasis.26, 46 Similar inhibitory effects on endothelial cell migration and angiogenesis with anti-CD151 antibodies are also known.20, 47 In addition, anti-CD151 monoclonal antibody strongly inhibits neurite outgrowth on laminin-5.48 Furthermore, the overall survival rate of patients with CD151-positive non-small-cell lung cancer is reported to be much lower than that of CD151-negative patients, suggesting that high CD151 gene expression is a marker of poor prognosis in human lung carcinomas.27 All these data support the notion that CD151 is a positive effecter of metastasis. As CD151 is able to bind with various integrin species, especially α3β1 and α6β4 integrins, it has been suggested that CD151–integrin protein complexes may play a role in cell motility probably through modulation of the interaction between integrin and ECM such as laminin-524 and also activation of focal adhesion kinase.46 However, these studies have completely lacked information about proteinases, which are also essential to tumor cell invasion and metastasis.1 In the present study, we have shown that proMMP-7 is captured and activated on the membranes of cultured cells by complex formation with CD151, and that the MMP-7 activity is detected around the human lung adenocarcinoma cell nests, in which CD151 and MMP-7 are colocalized. Thus, these data strongly suggest a hypothesis that MMP-7 may be involved in CD151-mediated progression in human lung carcinomas. The precise studies to demonstrate our hypothesis are now under way in our laboratory.
References
Sternlicht MD, Werb Z . How matrix metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol 2001;17:463–516.
Okada Y . Proteinases and Matrix Degradation. Kelley's Textbook of Rheumatology. Elsevier Saunders: Philadelphia, 2005, pp 63–81.
Wilson CL, Matrisian LM . Matrilysin: an epithelial matrix metalloproteinase with potentially novel functions. Int J Biochem Cell Biol 1996;28:123–136.
Saarialho-Kere UK, Crouch EC, Parks WC . Matrix metalloproteinase matrilysin is constitutively expressed in adult human exocrine epithelium. J Invest Dermatol 1995;105:190–196.
Dunsmore SE, Saarialho-Kere UK, Roby JD, et al. Matrilysin expression and function in airway epithelium. J Clin Invest 1998;102:1321–1331.
Wilson CL, Ouellette AJ, Satchell DP, et al. Regulation of intestinal alpha-defensin activation by the metalloproteinase matrilysin in innate host defense. Science 1999;286:113–117.
Wilson CL, Heppner KJ, Labosky PA, et al. Intestinal tumorigenesis is suppressed in mice lacking the metalloproteinase matrilysin. Proc Natl Acad Sci USA 1997;94:1402–1407.
Takeuchi N, Ichikawa Y, Ishikawa T, et al. Matrilysin gene expression in sporadic and familial colorectal adenomas. Mol Carcinogen 1997;19:225–229.
Yoshimoto M, Itoh F, Yamamoto H, et al. Expression of MMP-7(PUMP-1) mRNA in human colorectal cancers. Int J Cancer 1993;54:614–618.
Mori M, Barnard GF, Mimori K, et al. Overexpression of matrix metalloproteinase-7 mRNA in human colon carcinomas. Cancer 1995;75:1516–1519.
Ueno H, Yamashita K, Azumano I, et al. Enhanced production and activation of matrix metalloproteinase-7 (matrilysin) in human endometrial carcinomas. Int J Cancer 1999;84:470–477.
Kawano N, Osawa H, Ito T, et al. Expression of gelatinase A, tissue inhibitor of metalloproteinases-2, matrilysin, and trypsin(ogen) in lung neoplasms: an immunohistochemical study. Hum Pathol 1997;28:613–622.
Yamashita K, Azumano I, Mai M, et al. Expression and tissue localization of matrix metalloproteinase 7 (matrilysin) in human gastric carcinomas. Implications for vessel invasion and metastasis. Int J Cancer 1998;79:187–194.
Adachi Y, Yamamoto H, Itoh F, et al. Clinicopathologic and prognostic significance of matrilysin expression at the invasive front in human colorectal cancers. Int J Cancer 2001;95:290–294.
Imai K, Yokohama Y, Nakanishi I, et al. Matrix metalloproteinase 7 (matrilysin) from human rectal carcinoma cells. Activation of the precursor, interaction with other matrix metalloproteinases and enzymic properties. J Biol Chem 1995;270:6691–6697.
Yu WH, Woessner Jr JF, McNeish JD, et al. CD44 anchors the assembly of matrilysin/MMP-7 with heparin-binding epidermal growth factor precursor and ErbB4 and regulates female reproductive organ remodeling. Genes Dev 2002;16:307–323.
Knox JD, Wolf C, McDaniel K, et al. Matrilysin expression in human prostate carcinoma. Mol Carcinogen 1996;15:57–63.
Fitter S, Tetaz TJ, Berndt MC, et al. Molecular cloning of cDNA encoding a novel platelet-endothelial cell tetra-span antigen, PETA-3. Blood 1995;86:1348–1355.
Hasegawa H, Utsunomiya Y, Kishimoto K, et al. SFA-1, a novel cellular gene induced by human T-cell leukemia virus type 1, is a member of the transmembrane 4 superfamily. J Virol 1996;70:3258–3263.
Yanez-Mo M, Alfranca A, Cabanas C, et al. Regulation of endothelial cell motility by complexes of tetraspan molecules CD81/TAPA-1 and CD151/PETA-3 with alpha3 beta1 integrin localized at endothelial lateral junctions. J Cell Biol 1998;141:791–804.
Yauch RL, Kazarov AR, Desai B, et al. Direct extracellular contact between integrin alpha(3)beta(1) and TM4SF protein CD151. J Biol Chem 2000;275:9230–9238.
Sincock PM, Mayrhofer G, Ashman LK . Localization of the transmembrane 4 superfamily (TM4SF) member PETA-3 (CD151) in normal human tissues: comparison with CD9, CD63, and alpha5beta1 integrin. J Histochem Cytochem 1997;45:515–525.
Serru V, Le Naour F, Billard M, et al. Selective tetraspan-integrin complexes (CD81/alpha4beta1, CD151/alpha3beta1, CD151/alpha6beta1) under conditions disrupting tetraspan interactions. Biochem J 1999;340:103–111.
Sterk LM, Geuijen CA, Oomen LC, et al. The tetraspan molecule CD151, a novel constituent of hemidesmosomes, associates with the integrin alpha6beta4 and may regulate the spatial organization of hemidesmosomes. J Cell Biol 2000;149:969–982.
Fitter S, Sincock PM, Jolliffe CN, et al. Transmembrane 4 superfamily protein CD151 (PETA-3) associates with beta 1 and alpha IIb beta 3 integrins in haemopoietic cell lines and modulates cell–cell adhesion. Biochem J 1999;338:61–70.
Testa JE, Brooks PC, Lin JM, et al. Eukaryotic expression cloning with an antimetastatic monoclonal antibody identifies a tetraspanin (PETA-3/CD151) as an effector of human tumor cell migration and metastasis. Cancer Res 1999;59:3812–3820.
Tokuhara T, Hasegawa H, Hattori N, et al. Clinical significance of CD151 gene expression in non-small cell lung cancer. Clin Cancer Res 2001;7:4109–4114.
Inoki I, Shiomi T, Hashimoto G, et al. Connective tissue growth factor binds vascular endothelial growth factor (VEGF) and inhibits VEGF-induced angiogenesis. FASEB J 2002;16:219–221.
Guthrine C, Fink GR . Guide to Yeast Genetics and Molecular Biology. Methods in Enzymology. Academic Press: San Diego, 1991, pp 1–932.
Yauch RL, Hemler ME . Specific interactions among transmembrane 4 superfamily (TM4SF) proteins and phosphoinositide 4-kinase. Biochem J 2000;351:629–637.
Okada Y, Harris Jr ED, Nagase H . The precursor of a metalloendopeptidase from human rheumatoid synovial fibroblasts. Purification and mechanisms of activation by endopeptidases and 4-aminophenylmercuric acetate. Biochem J 1988;254:731–741.
Okada Y, Nagase H, Harris Jr ED . A metalloproteinase from human rheumatoid synovial fibroblasts that digests connective tissue matrix components. Purification and characterization. J Biol Chem 1986;261:14245–14255.
Nemori R, Yamamoto M, Kataoka F, et al. Development of in situ zymography to localize active matrix metalloproteinase-7 (matrilysin-1). J Histochem Cytochem 2005, 13 June [Epub ahead of print].
Hayakawa T, Yamashita K, Ohuchi E, et al. Cell growth-promoting activity of tissue inhibitor of metalloproteinases-2 (TIMP-2). J Cell Sci 1994;107:2373–2379.
Sato H, Takino T, Okada Y, et al. A matrix metalloproteinase expressed on the surface of invasive tumour cells. Nature 1994;370:61–65.
Guo H, Li R, Zucker S, et al. EMMPRIN (CD147), an inducer of matrix metalloproteinase synthesis, also binds interstitial collagenase to the tumor cell surface. Cancer Res 2000;60:888–891.
Brooks PC, Stromblad S, Sanders LC, et al. Localization of matrix metalloproteinase MMP-2 to the surface of invasive cells by interaction with integrin alpha v beta 3. Cell 1996;85:683–693.
Puyraimond A, Fridman R, Lemesle M, et al. MMP-2 colocalizes with caveolae on the surface of endothelial cells. Exp Cell Res 2001;262:28–36.
Yu Q, Stamenkovic I . Localization of matrix metalloproteinase 9 to the cell surface provides a mechanism for CD44-mediated tumor invasion. Genes Dev 1999;13:35–48.
Crabbe T, Willenbrock F, Eaton D, et al. Biochemical characterization of matrilysin. Activation conforms to the stepwise mechanisms proposed for other matrix metalloproteinases. Biochemistry 1992;31:8500–8507.
Lopez-Boado YS, Wilson CL, Hooper LV, et al. Bacterial exposure induces and activates matrilysin in mucosal epithelial cells. J Cell Biol 2000;148:1305–1315.
Lu PC, Ye H, Maeda M, et al. Immunolocalization and gene expression of matrilysin during corneal wound healing. Invest Ophthalmol Vis Sci 1999;40:20–27.
Karelina TV, Goldberg GI, Eisen AZ . Matrilysin (PUMP) correlates with dermal invasion during appendageal development and cutaneous neoplasia. J Invest Dermatol 1994;103:482–487.
Yauch RL, Berditchevski F, Harler MB, et al. Highly stoichiometric, stable, and specific association of integrin alpha3beta1 with CD151 provides a major link to phosphatidylinositol 4-kinase, and may regulate cell migration. Mol Biol Cell 1998;9:2751–2765.
Penas PF, Garcia-Diez A, Sanchez-Madrid F, et al. Tetraspanins are localized at motility-related structures and involved in normal human keratinocyte wound healing migration. J Invest Dermatol 2000;114:1126–1135.
Kohno M, Hasegawa H, Miyake M, et al. CD151 enhances cell motility and metastasis of cancer cells in the presence of focal adhesion kinase. Int J Cancer 2002;97:336–343.
Sincock PM, Fitter S, Parton RG, et al. PETA-3/CD151, a member of the transmembrane 4 superfamily, is localised to the plasma membrane and endocytic system of endothelial cells, associates with multiple integrins and modulates cell function. J Cell Sci 1999;112:833–844.
Stipp CS, Hemler ME . Transmembrane-4-superfamily proteins CD151 and CD81 associate with alpha 3 beta 1 integrin, and selectively contribute to alpha 3 beta 1-dependent neurite outgrowth. J Cell Sci 2000;113:1871–1882.
Acknowledgements
This work was supported by Scientific Research on Priority Areas 11240101 from the Ministry of Education, Science and Culture of Japan (to YO) and Keio University Grant-in-Aid for Encouragement of Young Medical Scientists (to TS). We thank Ms Michiko Uchiyama and Mr Hitoshi Abe for their skillful technical assistance. We are also grateful to Dr Atsushi Suzuki for critical review of our manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Shiomi, T., Inoki, I., Kataoka, F. et al. Pericellular activation of proMMP-7 (promatrilysin-1) through interaction with CD151. Lab Invest 85, 1489–1506 (2005). https://doi.org/10.1038/labinvest.3700351
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.3700351
Keywords
This article is cited by
-
CD151 in cancer progression and metastasis: a complex scenario
Laboratory Investigation (2014)
-
CD151
AfCS-Nature Molecule Pages (2011)
-
Functional interplay between tetraspanins and proteases
Cellular and Molecular Life Sciences (2011)
-
Assessment of myocardial blood perfusion improved by CD151 in a pig myocardial infarction model
Acta Pharmacologica Sinica (2009)
-
An in vitro model of light chain deposition disease
Kidney International (2009)
