
Abstract
The human sialidase, NEU4, has emerged as a possible regulator of neuronal differentiation and its overexpression has been demonstrated to promote the acquisition of a stem cell-like phenotype in neuroblastoma cells. In this paper, we demonstrated that glioblastoma stem cells (GSCs) isolated from glioblastoma multiforme (GBM) cell lines and patients’ specimens as neurospheres are specifically marked by the upregulation of NEU4; in contrast, the expression of NEU4 is very low in non-neurosphere-differentiated GBM cells. We showed that NEU4 silencing by miRNA or a chemical inhibitor of its catalytic activity triggered key events in GSCs, including (a) the activation of the glycogen synthase kinase 3β, with the consequent inhibition of Sonic Hedgehog and Wnt/β-catenin signalling pathways; (b) the decrease of the stem cell-like gene expression and marker signatures, evidenced by the reduction of NANOG, OCT-4, SOX-2, CD133 expression, ganglioside GD3 synthesis, and an altered protein glycosylation profile; and (c) a significant decrease in GSCs survival. Consistent with this finding, increased NEU4 activity and expression induced in the more differentiated GBM cells by the NEU4 agonist thymoquinone increased the expression of OCT-4 and GLI-1. Thus, NEU4 expression and activity appeared to help to determine the molecular signature of GSCs and to be closely connected with their survival properties. Given the pivotal role played by GSCs in GBM lethality, our results strongly suggest that NEU4 inhibition could significantly improve current therapies against this tumour.
Similar content being viewed by others
Main
Glioblastoma multiforme (GBM) is the most lethal and least successfully treated brain tumour,1 with a median survival of 15 months.2 Many studies have revealed that GBM includes a heterogeneous mixture of both cancer stem cells that possess the property of self-renewal, as well as more differentiated cancer cells.3, 4 GBM stem cells (glioblastoma stem cells (GSCs)) are believed to be responsible for GBM development, progression, recurrence, and therapeutic resistance.5, 6 GSCs share the expression of many markers, such as CD133 and nestin (NES),7 and core signalling pathways, such as Sonic Hedgehog (SHH)-GLI, Notch-1,8 and Wnt/β-catenin,9 with normal neural stem cells. These signalling pathways are known to sustain the long-term self-renewal and propagation of GSCs.8, 9 To identify GSCs features, recent papers have focused on the glycosylation status of some markers including CD133.10, 11 Furthermore, several glycoprotein markers involved in the differentiation status of GSCs were identified through a multi-lectin affinity chromatography and quantitative glycoproteomics approach.12 Regarding glycolipids, b-series gangliosides such as GD2 (Klassen et al.13) and GD3 (Yanagisawa et al.14 and Nakatani et al.15) have been previously demonstrated to be highly expressed in normal neural stem cells. In mouse neural stem cells, GD3 interacts with epidermal growth factor receptor (EGFR) stimulating its downstream signalling to support cell self-renewal capability.16 During mammalian brain development, the levels of simple gangliosides, that is, GD3 and GM3, decrease, while the synthesis of a-series gangliosides (GM1 and GD1a) increases.17 As the ganglioside pattern of neural stem cells seems to reflect their undifferentiated status and their self-renewal ability, it could be hypothesized that GSCs could also display an altered synthesis of gangliosides. The stage-specific embryonic antigenic-1 (CD15), a glycosylated epitope carried by glycoproteins and by glycolipids was associated with GSCs.18 Sialidases are enzymes critically involved in the control of protein and lipid glycosylation and altered in cancer.19, 20, 21, 22, 23 Sialidases (EC 3.2.1.18) catalytically remove sialic acid24 and, among many crucial cell events, in the nervous system they are involved in neuronal differentiation, neuritogenesis, and axonal growth.25, 26, 27 Among the four sialidases described so far, that is, NEU1, NEU2, NEU3, and NEU4, the latter appeared to be intriguingly related to the fate of neural cells. The level of NEU4 gene expression decreases during the development of the mouse brain and during retinoic acid-induced neuronal differentiation.28 Moreover, we previously reported that the NEU4 overexpression enhances an undifferentiated stem cell-like phenotype and cell proliferation in human neuroblastoma cells22 and, recently, it has been demonstrated that mouse neural stem cells highly express NEU4.29
Despite these findings, the role of sialidases in GBM pathogenesis remains to be firmly established. We first analysed sialidase expression in GSCs and we identified a significant increase of NEU4 expression in comparison with more differentiated GBM cells. Then, through NEU4 silencing and its chemical inhibition in GSCs, we demonstrated that (a) NEU4 is connected to the inhibition of glycogen synthase kinase-3β (GSK-3β) and, therefore, to the activation of the downstream pathways SHH and Wnt/β-catenin; (b) NEU4 expression is associated with the expression of the transcriptional factors, GLI-1, NANOG, OCT-4, and SOX-2; and (c) the silencing or chemical inhibition of NEU4 changes the entire glycosylation pattern of proteins and lipids, making it more similar to that of differentiated GBM cells and drastically reduces GSCs survival.
Results
Sialidase NEU4 expression is higher in GSCs isolated from GBM cell lines than in non-neurospheres differentiated GBM cells
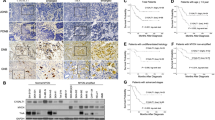
Human GBM cell lines U87MG, U138MG, and T98G were cultivated in selective medium, for 4 weeks, to promote the enrichment of GSCs, as previously reported.30 As shown in Figure 1a, after an initial steady phase of about 4 days, some U87MG and U138MG cells began to grow in suspension forming typical aggregates, referred to as neurospheres (U87MG duplication rate: 7 days; U138MG duplication rate: 14 days) (Figure 1b). We demonstrated that neurospheres formed by both U87MG and U138MG cells were highly enriched by GSCs through the evaluation of the expression of the stem cell markers CD133 (5.6-fold increase in U87MG cells and 4-fold increase in U138MG cells, after 4 weeks) and nestin (9.5-fold increase in U87MG cells and 1.8-fold increase in U138MG cells, after 4 weeks) (Figures 1c and d). After 4 weeks, the expression of these two markers did not further increase. Moreover, the GSC phenotype was further confirmed by functional assays of Hoechst 33342 dye exclusion and self-renewal (serial neurosphere formation) (data not shown). Instead, T98G cells were not able to survive in the selective medium, and after 14 days all cells died (Figure 1a). Accordingly, CD133 and nestin expression did not significantly increase (Figure 1e), indicating the absence of GSCs among T98G cells able to survive in these culture conditions. In parallel, we also determine the expression of sialidases NEU1, NEU3, and NEU4 in all three GBM cell lines and in neurospheres isolated from them. The cytosolic sialidase NEU2 was not expressed. NEU1 and NEU3 expression did not significantly change between U87MG-adherent cells and GSC-enriched neurospheres, or between U138MG-adherent cells and GSC-enriched neurospheres. Instead, NEU4 expression that was scarcely detectable in U87MG and in U138MG cells, increased by 14-fold comparing U87MG- and U138MG-adherent cells with the corresponding GSCs, after 4 weeks in selective medium (Figures 1f and g). We did not record the increment of NEU4 expression in T98G cells cultivated in the same selective medium (Figure 1h).

NEU4 expression in GBM cell lines and GSCs. (a) Growth curves of U87MG, U138MG, and T98G GBM cells cultivated in DMEM F12 medium plus supplements for up to 4 weeks. Values are the mean±S.D. of four independent experiments. Significance is based on the Student’s t-test: ***P<0.001 (values compared with that on Day 0). (b) Microphotographs of representative GBM cells and neurospheres formed by U87MG cells after 4 weeks of culture in the selective medium. Original magnification × 20 (Olympus Ix50). (c) Real-time PCR analysis of CD133 and nestin expression in U87MG cells; (d) U138MG cells; (e) T98G cells during for weeks of culture in DMEM F12 medium plus supplements. (f) Real-time PCR analysis of NEU1, NEU3, and NEU4 sialidases expression in U87MG cells; (g) U138MG cells; (h) T98G cells during 4 weeks of culture in DMEM F12 medium plus supplements. Values are the mean±S.D. of four independent experiments. Significance is based on the Student’s t-test: ***P<0.001 (values compared with starting cell lines)
NEU4 silencing impairs U87MG-GSCs survival
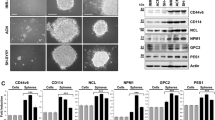
To determine the role of sialidase NEU4 and the significance of its increase in GSCs, after 4 weeks of culture in selective medium, GSCs isolated from U87MG cells (referred to as U-GSCs) were transfected with pcDNA 6.2-GW/EmGFP-miR carrying a miRNA specifically designed towards NEU4. The silencing effect was verified by real-time PCR (-60% as NEU4 mRNA expression) (Figure 2a). NEU4 silencing strongly impaired U-GSC survival: in fact, 6 days after the end of selection, mock U-GSCs began to grow (+25%); instead, 70% of NEU4 silencing U-GSCs (referred to as iNEU4 U-GSCs) died and, after 12 days, almost the 87% of cells died (Figure 2b). The clonogenic potential was determined performing limiting dilution assays: neurospheres were readily formed after 5 days by mock U-GSCs but not by iNEU4 U-GSCs (Figure 2c). Hoechst 33342 staining of mock U-GSCs and iNEU4 U-GSCs revealed the condensation of chromatin in iNEU4 U-GSCs, indicating an apoptotic phenotype (Figure 2d). The configuration of key proteins involved in cell cycle control revealed that iNEU4 U-GSCs were mainly blocked in the G2 phase: in fact, phospho-RB (Ser807/811) was not detected in contrast to mock U-GSCs, cyclin B1 was markedly accumulated, and inactive phospho-CDC2 (Tyr15) decreased (63%) (P<0.01) (Figure 2e). Moreover, we detected a marked downregulation of activated AKT (45%; P<0.01) and the appearance of the active fragment derived from the cleavage of caspase 3 in iNEU4 U-GSCs (Figure 2e), confirming the inhibition of pro-survival pathways and, instead, the parallel activation of signalling linked to cell death.

Effects of NEU4 silencing on U-GSCs survival. (a) Real-time PCR analysis of NEU4 sialidase expression in mock and iNEU4 U-GSCs. (b) Cell viability assessed through Trypan Blue after NEU4 silencing. (c) Neurosphere assay assessed after 6 days after NEU4 silencing. Original magnification × 20 (Olympus Ix50). (d) Hoechst 33342 staining in mock and iNEU4 U-GSCs. Pseudo-colouring approach used for this image. (e) Western blot analysis of P-RB (Ser 807/811), cyclin B1, P-CDC2 (Tyr15), P-AKT (Ser473), and active caspase 3 in mock and iNEU4 U-GSCs. β-Actin was used as loading control. Values are the mean±S.D. of four independent experiments. Significance is based on the Student’s t-test: **P<0.01, ***P<0.001
NEU4 silencing reduces the activation of signalling pathways related to stemness in U87MG-GSCs
Through the investigation of the molecular pathways altered by NEU4 silencing in GSCs and mainly related to their survival and maintenance, we identified a 74% decrease of phospho-glycogen synthase kinase-3β (GSK-3β) (Ser9) (inhibitory phosphorylation) in iNEU4 U-GSCs (Figure 3a) (P<0.001). This event was clearly related to many important subsequent effects: first, we detected a decrease in the mRNA expression of SHH (82%), protein patched homolog 1 (PTCH1) (70%), and GLI-1 (41.7%), clearly related to the activation of SHH pathway, and β-catenin (55.6%) and axin 2 (82%), clearly related to the activation of Wnt/β-catenin pathway, in iNEU4 U-GSCs (Figure 3b). Activated GSK-3β is a well-known inhibitor of both SHH31, 32 and Wnt/β-catenin33 signalling pathways. As SHH and Wnt/β-catenin pathways have a central role in GSCs behaviour by promoting an embryonic stem cell-like gene expression signature,34, 35 we investigated the expression of the known stem cell markers NANOG, OCT-4, and SOX-2. As shown in Figure 3c, in iNEU4 U-GSCs, NANOG expression decreased by 17%, OCT-4 by 71.5%, and SOX-2 by 42%, confirming the partial inhibition of the signalling pathways that fuel stem cell growth. Accordingly, the expression of the stemness marker CD133 decreased in iNEU4 U-GSCs (30%) (Figure 3c). Interestingly, also the content of the EGFR, which is usually constitutively activated in GSCs, decreased in iNEU4 U-GSCs (−74%; P<0.001) (Figure 3d).

Effects of NEU4 silencing on signalling pathways related to stemness. (a) Western blot analysis of P-GSK-3β (Ser9) in mock and iNEU4 U-GSCs. β-Actin was used as loading control. (b) Real-time PCR analysis of SHH, PTCH1, GLI-1, β catenin, axin 2, and (c) NANOG, OCT-4, SOX-2, and CD133 in mock and iNEU4 U-GSCs. (d) Western blot analysis of EGFR in mock and iNEU4 U-GSCs. β-Actin was used as loading control. Values are the mean±S.D. of five independent experiments. Significance is based on the Student’s t-test: *P<0.05, ***P<0.001
NEU4 silencing alters the sialo-glycoconjugate profile of U87MG-GSCs
As gangliosides and sialo-glycoproteins could be potential substrates of NEU4, we investigated sphingolipid pattern of GSCs after NEU4 silencing. It was evaluated by metabolic labelling with [3-3H]sphingosine. After a 2-h pulse followed by a 24-h chase, a metabolic steady state was obtained. The ganglioside profile of iNEU4 U-GSCs was clearly different from that of mock U-GSCs (Figures 4a and b). The content of all ganglioside, except GM3 and GM2, decreased in iNEU4 U-GSCs; in particular, GD1a, GD3, and GM1 underwent a 29, 74, and 55% decrease, respectively (Figure 4b). It is worth noting that these modifications were associated with alterations concerning the expression of GD3 synthase (−59%) (Figure 4c), which is regulated at a transcriptional level by Sp1 factors (−52.3% in iNEU4 U-GSCs) (Figure 4d). Significantly, after NEU4 silencing, GD3 synthase expression was reduced to the level shown by more differentiated U87MG cells (Figure 4d).

Modifications induced by NEU4 silencing on U-GSC sphingolipid pattern. (a) HPTC separation of mock and iNEU4 U-GSCs gangliosides. The doublets are due to the heterogeneity of the ceramide moiety. Solvent system: chloroform/methanol/0.2% aqueous CaCl2 60 : 40 : 9 (v/v). The image was acquired by radiochromatoscanning (Beta Imager 2000). (b) The ganglioside content of mock and iNEU4 U-GSCs. (c) Real-time PCR analysis of GD3 synthase and (d) Sp1 expression in mock and iNEU4 U-GSCs. (e) HPTLC separation of mock and iNEU4 U-GSCs neutral sphingolipids. Solvent system: chloroform/methanol/H2O 55 : 20 : 3 (v/v). The image was acquired by radiochromatoscanning (Beta Imager 2000). (f) The neutral sphingolipid content of mock and iNEU4 U-GSCs. The values are the mean±S.D. of five independent experiments. Significance is based on the Student’s t-test: **P<0.01, ***P<0.001
Among neutral sphingolipids, in iNEU4 U-GSCs, we detected the decrease of lactosylceramide (LacCer) (−30%), which is considered a pro-survival sphingolipid, and the parallel increase of ceramide (Cer) (+50%), which is known as a pro-apoptotic sphingolipid;36 glycosylceramide (GlcCer) did not change (Figures 4e and f).
We also investigated the α2-3 sialoglycoprotein profile of mock U-GSCs and iNEU4 U-GSCs (Figure 5a). Overall, the level of sialylation displayed by iNEU4 U-GSCs appeared to be reduced in comparison with mock U-GSCs with the exception of some glycoproteins of ∼50–60 kDa, which showed an increased content of sialic acid (Figure 5a). As for sphingolipids, it should be underlined that other enzymes involved in the sialylation of glycoproteins changed their expression following NEU4 silencing. Interestingly, α2-3 silayltransferase (ST3GalIII), which encodes a α2-3 sialyltransferase, was significantly increased as mRNA expression in U-GSCs than in U87MG cells (+70%), but decreased in iNEU4 U-GSCs (−70% in comparison with mock U-GSCs) reverting to the levels found in more differentiated U87MG cells (Figure 5b).

Modifications induced by NEU4 silencing on U-GSC sialoglycoprotein pattern. (a) The α2-3 sialoglycoprotein profile was assessed using western blotting and MAA (Maackia amurensis agglutinin) lectin staining of mock and iNEU4 U-GSCs. An equal content of protein was loaded in each lane. (b) Real-time PCR analysis of ST3GalIII expression in mock and iNEU4 U-GSCs. The values are the mean±S.D. of four independent experiments. Significance is based on the Student’s t-test: **P<0.01
Chemical inhibition or activation of NEU4 changes the expression of stem cell-like genes
To further corroborate the effects recorded after NEU4 silencing in U-GSCs, we tested the effects of a selective inhibitor of NEU4 previously described37 and referred to as C.6 below. We first checked the inhibition level caused by C.6 in assays of U-GSC sialidase activity. The assay was not selective and therefore included contributions of NEU1, NEU3, and NEU4 activities (NEU2 expression was not detected). As shown in Figures 6a, C.6 induced only a 16% inhibition of the total sialidase activity, likely to be due to its selectivity for NEU4.37 Under similar conditions, the nonspecific inhibitor, DANA (2,3-deydro-2-deoxy-N-acetylneuraminic acid), inhibited the 37% of total sialidase activity. We next cultivated U-GSCs in the presence of C.6 for 4 days and recorded the effects induced on the expression of stem-cell marker genes; C.6 treatment of U-GSCs decreased the expression of β-catenin (−28%), GLI-1 (−68%), NANOG (−75%), and OCT-4 (−84%) (Figure 6b), and the amount of phospho-GSK-3β (Ser9; 66.4%; P<0.01) (Figure 6c), reproducing the same effects seen for NEU4 silencing (Figures 3a–c). Moreover, the proliferation and survival capabilities of U-GSCs were impaired by C.6 treatment, similar to our observations in iNEU4 U-GSCs (Figure 2b); after 4 days of treatment, we observed that the initial number of U-GSCs was reduced by 44%, in contrast to non-treated U-GSCs that increased their number by 94% in the same period of culture (Figure 6d).

Effects induced by the chemical inhibition of NEU4 in U-GCS or activation in GBM cells. (a) In vitro sialidase activity of U-GSCs assayed towards 4-MU-NeuAc as substrate, with the inhibitors C.6 (20 μM) and DANA (20 μM). (b) Real-time PCR analysis of β-catenin, GLI-1, NANOG, and OCT-4 expression after in vivo treatment of U-GSCs with C.6 (0.7 mM) for 4 days. (c) Western blot analysis of P-GSK-3β (Ser9) in C.6-treated and non-treated U-GSCs. β-Actin was used as loading control. (d) Cell viability assessed through Trypan Blue after 4 days of GSCs treatment with C.6 (0.7 mM). (e) Real-time PCR analysis of OCT-4 and GLI-1 expression in U87MG GBM cells after 30 min of TQ treatment (0.3 mM). The values are the mean±S.D. of four independent experiments. Significance is based on the Student’s t-test: **P<0.01, ***P<0.001
In addition, we tested the effects of thymoquinone (TQ) on U87MG cells. TQ derived from the black cumin oil has been reported to be an agonist of NEU4 sialidase activity in live cells.38 Consistent with this, the expression of GLI-1 and OCT-4 increased by 775% and 700%, respectively, after treatment with TQ (Figure 6e).
NEU4 high expression characterizes GSCs isolated from human surgical specimens and regulates the activation of the stemness pathways
In order to confirm our data in cells directly isolated from patients and, therefore, to exclude the possibility that they could be related to long-term cultures of GBM cell lines, we assayed sialidase expression in GBM tissues and in GSCs isolated from six human surgical specimens, as previously described.39, 40 These GBM-derived GSCs were demonstrated to be endowed with long-term self-renewal, high in vivo tumorigenic potential, multipotency, and increased ability to establish and expand GBM-like tumours in serial transplantation.39 Among sialidases, NEU4 expression underwent the most significant change, increasing 40.3-fold in GBM-derived GSCs in comparison with GBM tissues. Instead, NEU1 expression showed a slight decrease in GSCs (1.6-fold) and NEU3 increased (13.4-fold) (Figure 7a).

Effects of NEU4 silencing and chemical inhibition in LO627 cells. (a) Real-time PCR of NEU1, NEU3, and NEU4 expression in GSCs isolated from six different surgical specimens and in five GBM tissues. Mean values of sialidase expression in GSCs and in GBM tissues were compared. (b) Cell viability assessed through Trypan Blue after NEU4 silencing in LO627 cells and (c) after 4 days of LO627 cells treatment with C.6 (0.7 mM). (d) Real-time analysis of SHH, PTCH1, GLI-1, β-catenin, axin 2, NANOG, OCT-4, SOX-2, and CD133 in mock and iNEU4 LO627. (e) Real-time PCR analysis of β-catenin, GLI-1, NANOG, and OCT-4 expression after in vivo treatment of LO627 cells with C.6 (0.7 mM) for 4 days. (f) Western blot analysis of P-GSK-3β (Ser9) in C.6-treated and non-treated LO627 cells. β-Actin was used as loading control. Significance is based on the Student’s t-test: *P<0.05; **P<0.01, ***P<0.001
In order to confirm the role of NEU4 in regulating the ‘stemness’ pathways emerged from the studies performed in U87MG cells, we silenced NEU4 in GSCs isolated from a surgical specimen (L0627 cells39). We achieved a 50% NEU4 silencing as mRNA. As observed for U87MG GSCs, NEU4 silencing strongly impaired LO627 GSC survival; in fact, 6 days after the end of selection mock LO627 cells began to grow (+48%); 18% of NEU4 silencing LO627-GSCs (referred to as LO627 iNEU4) died and, after 12 days almost 52% of LO627 iNEU4 cells died (Figure 7b). Moreover, C.6 treatment of LO627 cells reduced the initial number by 14%, in contrast to non-treated LO627 cells that increased their number by 100% after 4 days of culture (Figure 7c). Similar to U87MG cells, we identified in LO627 iNEU4 cells the decrease in mRNA expression of genes related to the SHH pathway (SHH: 83.5%; GLI-1: 59%), to the Wnt/β catenin pathway (β catenin: 30%; axin 2: 40%), and to ‘stemness’ transcriptional factors and markers (NANOG: 30%; OCT-4: 40%; SOX-2: 40%; CD133: 30%) (Figure 7d). Accordingly, C.6 treatment of LO627 cells reduced the expression of GLI-1 (37%), NANOG (37%), OCT-4 (20%) (Figure 7e), and phospho-GSK-3β (Ser9) content (65%; P<0.01) (Figure 7f).
Discussion
The sialidase NEU4 is the most recently identified sialidase isoenzyme;41 its physiological role inside the cell is still largely cryptic. Some recent papers have identified a strong involvement of NEU4 in neuronal differentiation and, in particular, its expression appeared to be elevated during the immature stage of neuronal cells or in undifferentiated neuroblastoma cells.22, 28, 29, 42 The data presented here are the first to demonstrate that NEU4 expression is high in GSCs isolated from both GBM cell lines and patients, and that its upregulation appears to be intimately correlated with the capability to form neurospheres. Cells that were not able to form neurospheres, such as T98G cells, did not show NEU4 upregulation. In contrast, in the bulk of more differentiated non-neurosphere GBM cells and in GBM surgical specimens, NEU4 expression is very low. NEU4 appears to be closely interconnected with the key pathways activated in GSCs responsible for their stemness and self-renewal properties. In fact, through silencing in U87MG-derived GSCs, we demonstrated that NEU4 activity affects the activation status of GSK-3β, inducing its inhibition (phosphorylation) when NEU4 is fully expressed. GSK-3β inhibition has been previously shown to significantly reduce the stem-like CD133+ fraction of a human GBM cell line, as well as the expression of the stem-cell markers, SOX-2 and nestin, while, in parallel, it increased the expression of differentiation markers, including glial fibrillary acidic protein, β-tubulin III and 2′, 3′-cyclic nucleotide 3′-phospohodiesterease.43 GSK-3β inhibition by administration of lithium or through specific inhibitors such as SB216763 has been demonstrated to impair neurosphere formation.43 The focal role had by GSK-3β in U-GSCs could be easily explained by interactions with the main signalling pathways, which sustain GSC self-renewal and propagation, SHH/GLI and Wnt/β-catenin.44, 45 Importantly, NEU4 silencing radically changed the molecular signature of U-GSCs, decreasing the expression of SHH, PTCH-1, and GLI-1 (key components of the SHH pathway) of β-catenin and axin 2 (key components of the Wnt/β-catenin pathway), and of genes regulated by them, such as NANOG, OCT-4, and SOX-2. We found that the expression of CD133 decreased, possibly because this gene is subjected to the transcriptional control of OCT-4 and SOX-2.46 In addition, the level of EGFR was significantly reduced. EGFR and β-catenin signalling are known to interact: intratumoral administration of β-catenin siRNA into subcutaneous gliomas in nude mice has been shown to transcriptionally downregulate EGFR.47 Therefore, we hypothesized that the decrease of β-catenin induced by NEU4 silencing could be responsible for the reduction in EGFR content. EGFR strongly regulates proliferation and migration of neural stem cells. GSCs constitutively activate EGFR, leading to the activation of AKT. Numerous studies have revealed the crucial role of the EGFR signalling cascade in the maintenance of GSCs, wherein EGF is capable of promoting neurosphere formation and self-renewal.48 Moreover, CD133 has been demonstrated to interact with the phosphoinositide 3-kinase 85 kDa regulatory subunit, resulting in the preferential activation of AKT in GSCs.49 Therefore, we speculate that the decrease of EGFR and CD133 in NEU4-silenced U-GSCs may act in synergy to reduce AKT signalling, and thus its pro-survival drive. The molecular revolution of U-GSCs phenotype subsequent to NEU4 silencing involved also the expression of GD3 synthase (ST8SiaI) that is clearly related to the presence of ganglioside GD3. GD3 is present in neural stem cells where it interacts with EGFR, stimulating its activation and its downstream signalling to maintain the self-renewal capability.16 In this study, we demonstrated that a high expression of GD3 synthase leads to a subsequent higher content of GD3 as a hallmark of GSCs. In normal neural stem cells, GD3 is linked to their self-renewal.15 GD3 synthase upregulation in GSCs appeared to be linked to the overexpression of Sp-1 factors,50 which, in turn, could be related also to EGFR activation.51 NEU4 silencing reverted the upregulation of GD3 synthase, decreasing the expression of Sp-1. We demonstrated that another feature of GSCs is the upregulation of ST3GalIII, which is involved in the de novo synthesis of carbohydrate antigens on different glycoproteins, and in GSCs it could be responsible for an altered pattern of glycosylation, already previously described.52 NEU4 silencing decreased ST3GalIII expression in U-GSCs, possibly decreasing the expression of Sp-1 factors that have been demonstrated to regulate the transcription of this gene.53 In this way, NEU4 silencing changed the glycosylation pattern of GSCs. Summing up, NEU4 silencing induced a sequence of interrelated molecular events, summarized in Figure 8, which significantly changed GSCs signalling and phenotype, leading to a significant decrease of their maintenance, as recorded by the inhibition of pro-survival pathways (AKT), the activation of caspase-3, the increase of the pro-apoptotic molecule, ceramide, and the decrease of the anti-apoptotic glycolipid, LacCer. Consequently, NEU4 silencing impaired the survival of U-GSCs inducing a block in the G2 phase and cell death.

Schematic picture of pathways regulated by NEU4 in GSCs
Importantly, the effects induced by NEU4 silencing on signalling pathways related to U-GSCs self-renewal and propagation, and on cell proliferation were confirmed in LO627 cells, that is, human short-term GSCs isolated from a surgical specimen, further confirming the key role had by NEU4.
Thus, in this emerging model, NEU4 appears to be intrinsically connected to the stem cell-like signalling of GSCs. As a further evidence of this, the treatment of U87MG GBM cells with TQ, an agonist of NEU4 activity,38 strongly enhanced the expression of the stem cell-like gene expression signature. Unfortunately, it is still unclear how NEU4 could interact with GSK-3β. On the basis of these data and our previous results that appeared to exclude the action of NEU4 on gangliosides and that identified, on the other hand, its efficient recognition of glycoproteins,22 we strongly suspect that also in GSCs NEU4 primarily acts on glycoproteins, because ganglioside profile did not change accordingly to NEU4 silencing. In fact, we recorded a decrease of GD1a and GD3, while GM3 remained unchanged after NEU4 silencing, instead of an increase, as expected, if these gangliosides were really substrates of NEU4 in GSCs. These ganglioside modifications could be related to alterations induced in the expression of other biosynthetic enzymes such as GD3 synthase, as discussed above. Notably, inspection of the sialo-glycoprotein pattern of NEU4-silenced GSCs showed dramatic changes from control GSCs. We identified some bands from 50 to 65 kDa that were more sialylated in NEU4-silenced GSCs. These sialoglycoproteins could therefore interact with GSK-3β, modulating its activation status based on the level of sialylation. On the other hand, in literature, several examples of sialoglycoproteins, including mucins, able to interact with GSK-3 have been described.54
Finally, we tested the effects of a previously identified specific inhibitor of NEU4 (Albohy et al.37) on both GSCs isolated from U87MG cell lines and in LO627 cells. We found that chemical inhibition of the enzyme resulted in similar effects to NEU4 silencing. Treatment of GSCs with the C.6 inhibitor markedly resembled NEU4 silencing, as it reduced the expression of β-catenin, GLI-1, NANOG, and OCT-4 stem-cell markers. Most significantly, we observed that treatment of GSCs with the NEU4 inhibitor, C.6, impaired cell proliferation and survival. Together, these finding confirms that the specific inhibition of NEU4 can be explored to suppress the growth and stem-cell phenotype of GSCs, and suggest that this strategy could form the basis of future therapeutic strategies for GBM.
Materials and Methods
Cell cultures
U87MG, U138MG, and T98G cells were purchased from ECACC (Sigma Aldrich, St. Louis, MO, USA) and cultured in RPMI 1640 supplemented with 10% (v/v) fetal bovine serum and 2 mM glutamine (Sigma Aldrich). GSCs were isolated from U87MG, U138MG, and T98G cells by plating 2 × 103 cells/cm2 in the selective medium DMEM/F12 (Sigma Aldrich) plus 10 ng/ml basic fibroblast growth factor (Millipore, Billerica, MA, USA), 20 ng/ml EGF (Sigma Aldrich), B27 supplement minus vitamin A (Life Technology, Carlsbad, CA, USA), and 2 mM glutamine, and culturing them for 3 weeks, at 37 °C, 5% CO2.
GSCs isolated from human post-surgical specimens and GBM tissues isolated from patients admitted at S. Raffaele Hospital (Milan, Italy) were provided by Dr R Galli. LO627 cells were cultured as previously described.39
NEU4 silencing in GSCs isolated from U87MG cells (U-GSCs) and in LO627 cells
To stably silence NEU4, an miRNA targeting the human NEU4 gene sequence was designed employing the BlockIT RNAi Designer software (Life Technologies) and inserted into the pcDNA 6.2-GW/EmGFP-miR vector (Life Technologies). U87-GSCs (5 × 105) were transfected overnight, using Lipofectamine LTX and Plus Reagent (Life Technologies), according to the manufacturer’s procedure. To stably silence NEU4 in LO627 cells, we employed the BlockIT lentiviral PolII miR RNAi Expression System (Life Technologies) and infected cells at an MOI of 5 according to the manufacturer’s instructions. Silenced U-GSCs and LO627 cells were isolated after selection with 10 μg/ml blasticidin for a week.
Chemical inhibition and activation of sialidase NEU4
The NEU4-specific inhibitor, 5-acetamido-9-[4-hydroxymethyl-1,2,3-triazol-1-y1]-2,3,5,9-tetradeoxy-D-glycerol-D-galacto-2-nonulopyranosonic acid, referred to as compound 6 (C.6) here, was synthesized by Albohy et al.37, as previously reported. C.6 (0.7 mM) was added to 4 × 105 U87-GSCs or LO627 cells in the presence of Lipofectamine LTX (Life Technologies) to allow the entry of the inhibitor inside the cells. In parallel, control cells were treated only with Lipofectamine LTX. Cells were collected after 4 days.
In order to stimulate NEU4 sialidase expression and enzymatic activity, 6 × 105 U87MG cells were treated with 0.3 mM TQ (Sigma Aldrich) for 30 min at 37 °C.
Cellular death assays
To determine the cell growth and viability, 5 × 104 mock U-GSCs, NEU4-silenced U-GSCs, mock LO627, NEU4-silenced LO627, C.6-treated U87-GSCs and C.6-treated LO627 cells were seeded in 96-well culture plates. Six (for NEU4-silenced cells) or four (for C.6-treated cells) days later, viable cells were counted with Trypan blue.
To test the clonogenic potential of mock and NEU4-silenced U-GSCs, a limiting dilution assay was performed plating 10–20 cells/well after neurosphere dissociation and counting cells in neurospheres newly formed after 4 days.
For Hoechst 33342 staining, 5 × 104 mock and NEU4-silenced U-GSCs were washed twice with PBS and incubated for 15 min with 10 μg/ml Hoechst 33342 in the dark at room temperature. The cells were then observed using an inverted fluorescence microscope (IX50 Olympus, Tokyo, Japan) and imaged.
Real-time PCR
Real-time PCR was performed as previously reported.19 Primer sequences were the following: NEU1: forward primer, 5′-CCTGGATATTGGCACTGAA-3′, reverse primer, 5′-CATCGCTGAGGAGACAGAAG-3′; NEU2: forward primer: 5′-TGCGCAGAGGAGACTACGA-3′, reverse primer, 5′-GTCTGCGCGTCATACAAGG-3′; NEU3: forward primer, 5′-GGCTTGTTTGGGTGTTTGTT-3′, reverse primer, 5′-TTTTGAATTGGCTTGGGTTC-3’; NEU4: forward primer, 5′-ACCGCCGAGAGTGTTTTGG-3′, reverse primer, 5′-CGTGGTCATCGCTGTAGAAGG-3′; NANOG: forward primer, 5′-GGTCCCAGTCAAGAAACAGA-3′, reverse primer, 5′-GAGGTTCAGGATGTTGGAGA-3′; OCT-4: forward primer, 5′-AGGAGAAGCTGGAGCAAAA-3′, reverse primer, 5′-GGCTGAATACCTTCCCAAA-3′; CD133: forward primer, 5′-CTAGCCTGCGGTCATCTCTC-3′, reverse primer, 5′-AGGCCATCCAAATCTGTCCT-3′; NES: forward primer, 5′-CCAAGAACTGGCTCAGGAAA-3′, reverse primer, 5′-TCTCCCTGCTCTACCACCTC-3′; ST3GALIII: forward primer, 5′-CATCATCGTGGGCAATGGAG-3′, reverse primer, 5′-TCAAAGCCTTTCACTGGTGC-3′; β catenin: forward primer, 5′-CATCATCGTGGGCAATGGAG-3′, reverse primer, 5′-GCACGAACAAGCAACTGAAC-3′; GLI-1: forward primer, 5′-TGAGCTGGACATGCTGGTT-3′, reverse primer, 5′-GATTCAGGCTCACGCTTCTC-3′; SHH: forward primer: 5′-CCCCAATTACAACCCCGACA-3′, reverse primer, 5′-AGTTTCACTCCTGGCCACTG-3′; PTCH1: forward primer: 5′-TGCCATTTCTCGCTCTTGGT-3′, reverse primer: 5′-GAATTGGGATTAACGCGGCC-3′; axin 2: forward primer: 5′-GGCTCCAGAAGATCACAAAG-3′, reverse primer: 5′-TATGGAATTTCTTCCCCACA-3′; GD3 synthase: forward primer, 5′-CCCTGAACCAGTTCGATGTT-3′, reverse primer, 5′-CAGCATAATTCGGCAAAGGT-3′; SOX-2: forward primer, 5′-GCACATGAACGGCTGGAGCAACG-3′, reverse primer, 5′-TGCTGCGAGTAGGACATGCTGTAGG-3′; β actin: forward primer, 5′-CGACAGGATGCAGAAGGAG-3′, reverse primer, 5′-ACATCTGCTGGAAGGTGGA-3′.
Western blotting
Proteins (25 μg) were separated on 10% SDS-PAGE, and transferred onto PVDF membrane. The following antibodies were used for the assays: anti-β actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and anti-P-AKT (Ser473), anti-EGFR, anti-cleaved caspase 3, anti-P-RB (Ser807/811), anti-cyclin B1, P-CDC2 (Tyr15) and anti-GSK-3β (Ser9) (Cell Signaling, Danvers, MA, USA). Blots were quantified using GS-700 calibrated densitometer and the Quantity One software (Bio-Rad, Hercules, CA, USA).
Metabolic labelling of cell sphingolipids
Sphingolipid analysis was carried out through cell metabolic labelling with [3-3H]sphingosine (PerkinElmer, Waltham, MA, USA), as previously reported.20 Ganglioside and neutral sphingolipid extracts were analysed by HPTLC carried out with the solvent systems chloroform/methanol/0.2% CaCl2 55 : 45 : 6 (v/v) and chloroform/methanol/water 110 : 40 : 6 (v/v), respectively.
Sialoglycoprotein analysis
Proteins (30 μg) of mock and NEU4-silenced U-GSCs were separated on a 10% SDS-PAGE gel and transferred onto a PVDF membrane. Sialoglycoproteins were visualized employing the DIG Glycan Differentiation kit (Roche Applied Science, Indianapolis, IN, USA) as previously reported.22 Densitometric analysis was performed using Quantity One software.
Sialidase activity
The sialidase activity present in the particulate fraction of U-GSCs was assayed using 4-MU-NeuAc (4-methylumbelliferyl-N-acetylneuraminic acid) (Sigma Aldrich) substrate, as previously reported.42 To test the inhibitory effect of C.6 or that of DANA, the inhibitors (20 μM) were added to the enzymatic mixture. One unit of sialidase activity is defined as the amount of enzyme liberating 1 μmol of product per min.
Statistical analysis
The values are presented as the mean±S.D. The statistical analyses were performed using Student’s t-test.
Abbreviations
- GBM:
-
glioblastoma multiforme
- GSC:
-
glioblastoma stem cell
- SHH:
-
Sonic Hedgehog
- EGFR:
-
epidermal growth factor receptor
- PTCH1:
-
protein patched homolog 1
- TQ:
-
thymoquinone
- DANA:
-
2,3-deydro-2-deoxy-N-acetylneuraminic acid
- GSK-3β:
-
glycogen synthase kinase 3β
References
Furnari FB, Fenton T, Bachoo RM, Mukasa A, Stommel JM, Stegh A et al. Malignant astrocytic glioma: genetics, biology, and paths to treatment. Genes Dev 2007; 21: 2683–2710.
Das P, Puri T, Jha P, Pathak P, Joshi N, Suri V et al. A clinicopathological and molecular analysis of glioblastoma multiforme with long-term survival. J Clin Neurosci 2011; 18: 66–70.
Visvader JE, Lindeman GJ . Cancer stem cells: current status and evolving complexities. Cell Stem Cell 2012; 10: 717–728.
Akhtar K, Bussen W, Scott SP . Cancer stem cells - from initiation to elimination, how far have we reached? (Review). Int J Oncol 2009; 34: 1491–1503.
Sanai N, Alvarez-Buylla A, Berger MS . Neural stem cells and the origin of gliomas. N Engl J Med 2005; 353: 811–822.
Cheng L, Bao S, Rich JN . Potential therapeutic implications of cancer stem cells in glioblastoma. Biochem Pharmacol 2010; 80: 654–665.
Perez Castillo A, Aguilar-Morante D, Morales-Garcia JA, Dorado J . Cancer stem cells and brain tumors. Clin Transl Oncol 2008; 10: 262–267.
Kristoffersen K, Villingshoj M, Poulsen HS, Stockhausen MT . Level of Notch activation determines the effect on growth and stem cell-like features in glioblastoma multiforme neurosphere cultures. Cancer Biol Ther 2013; 14: 625–637.
Bell D, Miele L . A magnifying glass on glioblastoma stem cell signaling pathways. Cancer Biol Ther 2011; 11: 765–768.
Kemper K, Sprick MR, de Bree M, Scopelliti A, Vermeulen L, Hoek M et al. The AC133 epitope, but not the CD133 protein, is lost upon cancer stem cell differentiation. Cancer Res 2010; 70: 719–729.
Lehnus KS, Donovan LK, Huang X, Zhao N, Warr TJ, Pilkington GJ et al. CD133 glycosylation is enhanced by hypoxia in cultured glioma stem cells. Int J Oncol 2013; 42: 1011–1017.
He J, Liu Y, Zhu TS, Xie X, Costello MA, Talsma CE et al. Glycoproteomic analysis of glioblastoma stem cell differentiation. J Proteome Res 2011; 10: 330–338.
Klassen H, Schwartz MR, Bailey AH, Young MJ . Surface markers expressed by multipotent human and mouse neural progenitor cells include tetraspanins and non-protein epitopes. Neurosci Lett 2001; 312: 180–182.
Yanagisawa M, Nakamura K, Taga T . Roles of lipid rafts in integrin-dependent adhesion and gp130 signalling pathway in mouse embryonic neural precursor cells. Genes Cells 2004; 9: 801–809.
Nakatani Y, Yanagisawa M, Suzuki Y, Yu RK . Characterization of GD3 ganglioside as a novel biomarker of mouse neural stem cells. Glycobiology 2010; 20: 78–86.
Wang J, Yu RK . Interaction of ganglioside GD3 with an EGF receptor sustains the self-renewal ability of mouse neural stem cells in vitro. Proc Natl Acad Sci USA 2013; 110: 19137–19142.
Yanagisawa M . Stem cell glycolipids. Neurochem Res 2011; 36: 1623–1635.
Son MJ, Woolard K, Nam DH, Lee J, Fine HA . SSEA-1 is an enrichment marker for tumor-initiating cells in human glioblastoma. Cell Stem Cell 2009; 4: 440–452.
Tringali C, Lupo B, Anastasia L, Papini N, Monti E, Bresciani R et al. Expression of sialidase Neu2 in leukemic K562 cells induces apoptosis by impairing Bcr-Abl/Src kinases signaling. J Biol Chem 2007; 282: 14364–14372.
Tringali C, Lupo B, Cirillo F, Papini N, Anastasia L, Lamorte G et al. Silencing of membrane-associated sialidase Neu3 diminishes apoptosis resistance and triggers megakaryocytic differentiation of chronic myeloid leukemic cells K562 through the increase of ganglioside GM3. Cell Death Differ 2009; 16: 164–174.
Tringali C, Lupo B, Silvestri I, Papini N, Anastasia L, Tettamanti G et al. The plasma membrane sialidase NEU3 regulates the malignancy of renal carcinoma cells by controlling beta1 integrin internalization and recycling. J Biol Chem 2012; 287: 42835–42845.
Tringali C, Cirillo F, Lamorte G, Papini N, Anastasia L, Lupo B et al. NEU4L sialidase overexpression promotes beta-catenin signaling in neuroblastoma cells, enhancing stem-like malignant cell growth. Int J Cancer 2012; 131: 1768–1778.
Miyagi T, Wada T, Yamaguchi K, Shiozaki K, Sato I, Kakugawa Y et al. Human sialidase as a cancer marker. Proteomics 2008; 8: 3303–3311.
Monti E, Preti A, Venerando B, Borsani G . Recent development in mammalian sialidase molecular biology. Neurochem Res 2002; 27: 649–663.
Proshin S, Yamaguchi K, Wada T, Miyagi T . Modulation of neuritogenesis by ganglioside-specific sialidase (Neu 3) in human neuroblastoma NB-1 cells. Neurochem Res 2002; 27: 841–846.
Da Silva JS, Hasegawa T, Miyagi T, Dotti CG, Abad-Rodriguez J . Asymmetric membrane ganglioside sialidase activity specifies axonal fate. Nat Neurosci 2005; 8: 606–615.
Kappagantula S, Andrews MR, Cheah M, Abad-Rodriguez J, Dotti CG, Fawcett JW . Neu3 sialidase-mediated ganglioside conversion is necessary for axon regeneration and is blocked in CNS axons. J Neurosci 2014; 34: 2477–2492.
Shiozaki K, Koseki K, Yamaguchi K, Shiozaki M, Narimatsu H, Miyagi T . Developmental change of sialidase neu4 expression in murine brain and its involvement in the regulation of neuronal cell differentiation. J Biol Chem 2009; 284: 21157–21164.
Itokazu Y, Yu RK . Amyloid beta-peptide 1-42 modulates the proliferation of mouse neural stem cells: upregulation of fucosyltransferase IX and notch signaling. Mol Neurobiol 2014; e-pub ahead of print 17 January 2014.
Beier D, Hau P, Proescholdt M, Lohmeier A, Wischhusen J, Oefner PJ et al. CD133(+) and CD133(-) glioblastoma-derived cancer stem cells show differential growth characteristics and molecular profiles. Cancer Res 2007; 67: 4010–4015.
Price MA, Kalderon D . Proteolysis of the Hedgehog signaling effector Cubitus interruptus requires phosphorylation by glycogen synthase kinase 3 and casein kinase 1. Cell 2002; 108: 823–835.
Jia J, Amanai K, Wang G, Tang J, Wang B, Jiang J . Shaggy/GSK3 antagonizes Hedgehog signalling by regulating Cubitus interruptus. Nature 2002; 416: 548–552.
Ferkey DM, Kimelman D . GSK-3: new thoughts on an old enzyme. Dev Biol 2000; 225: 471–479.
Santoni M, Burattini L, Nabissi M, Morelli MB, Berardi R, Santoni G et al. Essential role of Gli proteins in glioblastoma multiforme. Curr Protein Pept Sci 2013; 14: 133–140.
Rossi M, Magnoni L, Miracco C, Mori E, Tosi P, Pirtoli L et al. Beta-catenin and Gli1 are prognostic markers in glioblastoma. Cancer Biol Ther 2011; 11: 753–761.
Morad SA, Cabot MC . Ceramide-orchestrated signalling in cancer cells. Nat Rev Cancer 2013; 13: 51–65.
Albohy A, Zhang Y, Smutova V, Pshezhetsky AV, Cairo CW . Identification of selective nanomolar inhibitors of the human neuraminidase, NEU4. ACS Med Chem Lett 2013; 4: 532–537.
Finlay TM, Jayanth P, Amith SR, Gilmour A, Guzzo C, Gee K et al. Thymoquinone from nutraceutical black cumin oil activates Neu4 sialidase in live macrophage, dendritic, and normal and type I sialidosis human fibroblast cells via GPCR Galphai proteins and matrix metalloproteinase-9. Glycoconj J 2010; 27: 329–348.
Galli R, Binda E, Orfanelli U, Cipelletti B, Gritti A, De Vitis S et al. Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res 2004; 64: 7011–7021.
Mazzoleni S, Politi LS, Pala M, Cominelli M, Franzin A, Sergi Sergi L et al. Epidermal growth factor receptor expression identifies functionally and molecularly distinct tumor-initiating cells in human glioblastoma multiforme and is required for gliomagenesis. Cancer Res 2010; 70: 7500–7513.
Monti E, Bassi MT, Bresciani R, Civini S, Croci GL, Papini N et al. Molecular cloning and characterization of NEU4, the fourth member of the human sialidase gene family. Genomics 2004; 83: 445–453.
Bigi A, Tringali C, Forcella M, Mozzi A, Venerando B, Monti E et al. A proline-rich loop mediates specific functions of human sialidase NEU4 in SK-N-BE neuronal differentiation. Glycobiology 2013; 23: 1499–1509.
Korur S, Huber RM, Sivasankaran B, Petrich M, Morin P Jr, Hemmings BA et al. GSK3beta regulates differentiation and growth arrest in glioblastoma. PLoS One 2009; 4: e7443.
Dahmane N, Sanchez P, Gitton Y, Palma V, Sun T, Beyna M et al. The Sonic Hedgehog-Gli pathway regulates dorsal brain growth and tumorigenesis. Development 2001; 128: 5201–5212.
Clement V, Sanchez P, de Tribolet N, Radovanovic I, Ruiz i Altaba A . HEDGEHOG-GLI1 signaling regulates human glioma growth, cancer stem cell self-renewal, and tumorigenicity. Curr Biol 2007; 17: 165–172.
Iida H, Suzuki M, Goitsuka R, Ueno H . Hypoxia induces CD133 expression in human lung cancer cells by up-regulation of OCT3/4 and SOX2. Int J Oncol 2012; 40: 71–79.
Yue X, Lan F, Yang W, Yang Y, Han L, Zhang A et al. Interruption of beta-catenin suppresses the EGFR pathway by blocking multiple oncogenic targets in human glioma cells. Brain Res 2010; 1366: 27–37.
Paul I, Bhattacharya S, Chatterjee A, Ghosh MK . Current understanding on EGFR and Wnt/beta-catenin signaling in glioma and their possible crosstalk. Genes Cancer 2013; 4: 427–446.
Wei Y, Jiang Y, Zou F, Liu Y, Wang S, Xu N et al. Activation of PI3K/Akt pathway by CD133-p85 interaction promotes tumorigenic capacity of glioma stem cells. Proc Natl Acad Sci USA 2013; 110: 6829–6834.
Furukawa K, Horie M, Okutomi K, Sugano S, Furukawa K . Isolation and functional analysis of the melanoma specific promoter region of human GD3 synthase gene. Biochim Biophys Acta 2003; 1627: 71–78.
Xu K, Shu HK . EGFR activation results in enhanced cyclooxygenase-2 expression through p38 mitogen-activated protein kinase-dependent activation of the Sp1/Sp3 transcription factors in human gliomas. Cancer Res 2007; 67: 6121–6129.
Cheray M, Petit D, Forestier L, Karayan-Tapon L, Maftah A, Jauberteau MO et al. Glycosylation-related gene expression is linked to differentiation status in glioblastomas undifferentiated cells. Cancer Lett 2011; 312: 24–32.
Taniguchi A, Saito K, Kubota T, Matsumoto K . Characterization of the promoter region of the human Galbeta1,3(4)GlcNAc alpha2,3-sialyltransferase III (hST3Gal III) gene. Biochim Biophys Acta 2003; 1626: 92–96.
Schroeder JA, Adriance MC, Thompson MC, Camenisch TD, Gendler SJ . MUC1 alters beta-catenin-dependent tumor formation and promotes cellular invasion. Oncogene 2003; 22: 1324–1332.
Acknowledgements
This study was supported by grant AIRC (IG-13131) to BV and CT.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by A Stephanou
Rights and permissions
Cell Death and Disease is an open-access journal published by Nature Publishing Group. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article

Cite this article
Silvestri, I., Testa, F., Zappasodi, R. et al. Sialidase NEU4 is involved in glioblastoma stem cell survival. Cell Death Dis 5, e1381 (2014). https://doi.org/10.1038/cddis.2014.349
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cddis.2014.349
This article is cited by
-
Insights into the role of sialylation in cancer progression and metastasis
British Journal of Cancer (2021)
-
Sialidase neu4 deficiency is associated with neuroinflammation in mice
Glycoconjugate Journal (2021)
-
The multifaceted NF-kB: are there still prospects of its inhibition for clinical intervention in pediatric central nervous system tumors?
Cellular and Molecular Life Sciences (2021)
-
Sialylation: an Avenue to Target Cancer Cells
Pathology & Oncology Research (2016)
