
Abstract
A and B alleles at the ABO genetic locus specify A and B glycosyltransferases that catalyze the biosynthesis of A and B oligosaccharide antigens, respectively, of blood group ABO system which is important in transfusion and transplantation medicine. GBGT1 gene encodes Forssman glycolipid synthase (FS), another glycosyltransferase that produces Forssman antigen (FORS1). Humans are considered to be Forssman antigen-negative species without functional FS. However, rare individuals exhibiting Apae phenotype carry a dominant active GBGT1 gene and express Forssman antigen on RBCs. Accordingly, FORS system was recognized as the 31st blood group system. Mouse ABO gene encodes a cis-AB transferase capable of producing both A and B antigens. This murine enzyme contains the same GlyGlyAla tripeptide sequence as FSs at the position important for the determination of sugar specificity. We, therefore, transfected the expression construct into appropriate recipient cells and examined whether mouse cis-AB transferase may also exhibit FS activity. The result was positive, confirming the crosstalk between the ABO and FORS systems. Further experiments have revealed that the introduction of this tripeptide sequence to human A transferase conferred some, although weak, FS activity, suggesting that it is also involved in the recognition/binding of acceptor substrates, in addition to donor nucleotide-sugars.
Similar content being viewed by others

Introduction
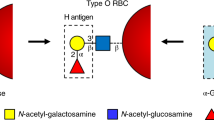
The ABO system is one of the most important blood group systems in transfusion medicine1,2. This system is composed of A and B oligosaccharide antigens expressed on red blood cells (RBCs), and anti-A and anti-B antibodies against those antigens in serum. Although both antigens and antibodies are involved, this hereditary trait is specified by a single genetic locus of ABO. Functional A and B alleles encode blood group A and B glycosyltransferases with similar, but distinct, specificities. A transferase transfers an N-acetyl-D-galactosamine (GalNAc) to oligosaccharide acceptor substrate, H substance (Fucα1-2Gal-), to produce A antigen, whereas B transferase transfers a galactose to the same acceptor to produce B antigen. The immunodominant structures of A and B antigens are GalNAcα1-3(Fucα1-2)Gal- and Galα1-3(Fucα1-2)Gal-, respectively. O allele-encoded proteins are enzymatically inactive and do not possess either of the transferase activities. Accordingly, H substance remains without further modifications in blood group O individuals. Presence or absence of the antibodies is determined secondarily to the absence or presence of the corresponding antigens. In other words, individuals expressing A and/or B antigens do not possess the antibodies against those antigens, however, individuals not expressing A and/or B antigens possess the antibodies against those antigens (Landsteiner’s Law)3,4. We previously elucidated the molecular genetic basis of ABO system5. A and B alleles encode proteins that are different by 4 amino acid residues at codons 176, 235, 266, and 268. They are Arg, Gly, Leu, and Gly in A transferase, and Gly, Ser, Met, and Ala in B transferase. The latter 2 codons of the 4 amino acid substitutions determine sugar specificities of A and B transferases, whereas the first and second codons exhibit no and slight effects, respectively. Many O alleles are inactive due to a single nucleotide deletion (261delG)5, although inactivating missense mutations can also be found (Gly268Arg, for instance)6.
The FORS system is another system of RBC polymorphism expressing heterophilic Forssman antigen (FORS1) at low-prevalence. Forssman antigen was initially noted by Frouin and re-discovered by Forssman when antibodies produced by injecting rabbits with a suspension of kidney tissue from guinea pig were found to lyse sheep erythrocytes in the presence of the complement proteins7. Those antibodies were later used to classify animals into Forssman antigen-positive and Forssman antigen-negative species, depending on the antigen expression8,9. For instance, guinea pig, sheep, and mouse are Forssman-positive, whereas rabbit and human are Forssman-negative. Chemical determination showed that Forssman antigen is carried on a pentasaccharide glycolipid with the structure of GalNAcα1-3GalNAcβ1-3Galα1-4Galβ1-4Glcβ1-Cer10. Forssman glycolipid synthase (FS) catalyzes the final step of its biosynthesis11,12. Canine GBGT1 gene cDNA encoding FS was cloned by Haslam and Baenziger, and GBGT1 and ABO genes were found to be paralogous genes that were derived from the same ancestral gene by gene duplication and subsequent divergence13.
Forssman antigen has only lately been taken into consideration for blood transfusion. It was known before that RBCs from rare individuals exhibiting the phenotype named Apae reacted strongly with Helix pomatia lectin, weakly with polyclonal anti-A antibodies, but not with monoclonal anti-A antibodies14. Because of reactivity to polyclonal anti-A antibodies, Apae was thought to be an A subgroup. However, recent chemical characterization of glycolipids on Apae RBCs has identified Forssman glycolipid15,16. The molecular genetic analysis demonstrated that Apae individuals had a GBGT1 gene containing the Arg296Gln substitution when compared with FS protein of ordinary non-Apae individuals. Forssman glycolipid is immunologically and chemically distinct from blood group A and B structures, GBGT1 gene is different from ABO gene, and anti-Forssman antigen antibodies exist. Therefore, the International Society of Blood Transfusion (ISBT) has recognized FORS system as the 31st blood group system with one low-prevalence allele expressing Forssman antigen (FORS1).
Excluding special case of Apae individuals, humans are thought to be Forssman antigen-negative species. Strangely, however, Forssman antigen has been detected in a variety of human tumors, including metastatic tumor of biliary adenocarcinoma in liver17, stomach and colon tumors18,19,20, and lung tumors21. Forssman antigen expression has also been observed in human cancer cell lines of lung, breast, and stomach22,23. Why and how Forssman glycan is expressed in cancer remains a mystery.
Over the last 2 decades, we have been investigating ABO genes in other species than humans. We first studied primate ABO genes, and found that the latter 2 of the 4 amino acid substitutions between human A and B transferases are conserved in primates, depending on A or B specificity24. We then cloned porcine A transferase cDNA and demonstrated that the major portion of A transferase coding sequence is missing in the O allele of pig AO system25. We also cloned ABO gene cDNAs from mouse species, and demonstrated that mouse ABO gene encodes cis-AB transferase, an enzyme that is capable of producing both A and B antigens26. In recent years, we have also expanded our studies to the other members of α1,3-Gal(NAc) transferase family of genes. In addition to GBGT1, those genes include GGTA1 genes, which encode α1,3-galactosyltransferases to synthesize α1,3-galactosyl epitope (Galα1-3Galβ1-4GlcNAcβ-), A3GALT2 genes, which encode isogloboside b3 synthase to synthesize isogloboside b3 (Galα1-3Galβ1-4Glcβ1-Cer), and GLT6D1 (glycosyltransferase 6 domain containing 1) genes, whose transferase activity remains to be examined.
The gene we initially investigated was GBGT1. We compared amino acid sequences of FS proteins between Forssman-positive and Forssman-negative species. We then prepared structural chimeras between functional mouse FS and non-functional human FS protein, determined FS activity by DNA transfection of the expression constructs into COS1 African green monkey kidney cells followed by immunostaining with anti-Forssman monoclonal antibody27. We narrowed down the regions responsible for the human inactivity. We then prepared amino acid substitution constructs by in vitro mutagenesis, and performed additional DNA transfection-immunostaining experiments. We found that two substitutions (c.688G > A [p.Gly230Ser] and c.887A > G [p.Gln296Arg]) are responsible for the inactivation of human FS. Codon 230 of human/mouse FS proteins corresponds to codon 235 of human A/B transferase, and the glycine residue at this position was previously shown to be important for GalNAc specificity of A transferase28. We also demonstrated that the reversion of one of the two substitutions in human FS protein conferred weak FS activity whereas the reversion of the two restored FS activity to a level equivalent of mouse FS. In conclusion, we demonstrated the molecular mechanism that resulted in Forssman negativity in humans. In the same paper we also characterized probable mechanisms of inactivation in several other Forssman-negative species including cow, rat, and rabbit27. We also investigated the evolution of α1,3-Gal(NAc) transferase family of genes and produced a phylogenetic tree of those glycosyltransferase genes29. One important finding from that study was that the tripeptide sequence corresponding to codons 266–268 of human A and B transferases is conserved to be GlyGlyAla in GBGT1 genes from all the species that were examined except for cow, 3 Old World monkeys, and several fish species, suggesting that this tripeptide sequence is important for FS activity. Surprisingly, however, the same GlyGlyAla tripeptide was also found in mouse ABO gene-encoded cis-AB transferase. We, therefore, examined whether or not mouse cis-AB transferase may also possess FS activity, in addition to A and B transferase activities.
Results
Conservation of GlyGlyAla (GGA) tripeptide sequence in the majority of GBGT1 gene at the position corresponding to codons 266–268 of human blood group A and B transferases
Among 69 Ensembl species, 46 species were found to possess GBGT1 gene(s), whereas the remaining 23 have none. Excluding fishes where variations are frequent, GBGT1 genes from the majority of other species contain GlyGlyAla (GGA) tripeptide sequence at the equivalent position of codons 266–268 of human A and B transferases (Supplemental Table 1). The exceptions are cow containing GlyArgAla (GRA) and three Old World monkeys (macaque, olive baboon, and Vervet Monkey or African Green Monkey (vervet-AGM)) containing GlyGlyLys (GGK). There is a deletion of 33 nucleotides (11 amino acids) at 12 nucleotides (4 amino acids) downstream of the lysine residue in the bovine GBGT1 gene. Because this region of protein is critical for the catalytic activity of FS proteins, the deletion seems to render cow FS protein enzymatically non-functional. GBGT1 genes from the 3 primates are unlikely to be functional, either, considering that the substitution of codon 268 of human A and B transferases by the lysine residue abolished either of the transferase activities30. It should also be noted that several fish GBGT1 genes also contain the GlyGlyAla tripeptide. Taken together, it was hypothesized that the conserved GlyGlyAla tripeptide sequence plays a significant role for GBGT1 gene-encoded FS activity.
Presence of GlyGlyAla (GGA) tripeptide sequence in the corresponding position of mouse cis-AB transferase
The amino acid sequences of blood group A/B/cis-AB transferases and FS proteins were MEGA-aligned, and are shown in Fig. 1. The proteins analyzed are human A transferase (H_ABO-A), human B transferase (H_ABO-B), human cis-AB transferase (H_ABO-AB), mouse cis-AB transferase (M_ABO-AB), human GBGT1-encoded non-functional FS protein (H_GBGT1), human GBGT1 FS with Arg296Gln substitution responsible for Apae phenotype (H_GBGT1 + ), and mouse GBGT1 FS (M_GBGT1). The amino acids corresponding to codons 176, 235, 266, and 268 of human A and B transferases, as well as codon 296 of FS proteins, are shown in bold type. Those amino acids that are conserved in all the FS proteins and also in mouse cis-AB transferase (M_ABO-AB) are highlighted.

Amino acid sequences of the coding regions of human A, B, and cis-AB transferases encoded by A101 (H_ABO-A), B101 (H_ABO-B), and cis-AB01 (H_ABO-AB) alleles of ABO gene, mouse cis-AB transferase encoded by mouse ABO gene (M_ABO-AB), human FS protein encoded by Forssman-negative GBGT1 gene (H_GBGT1) from ordinary individuals and that of Forssman-positive GBGT1 gene (H_GBGT1+) from rare Apae individuals, and mouse FS encoded by GBGT1 gene (M_GBGT1) were aligned and are shown. When amino acids are conserved in all the proteins, they are marked above with an equal symbol (=). 116 out of 274 amino acids (42.3%) in the last 2 coding exons are conserved in all those proteins. When amino acids of mouse cis-AB transferase are identical to all the other human A/B/cis-AB transferases, they are marked above with a plus symbol (+). Similarly, when amino acids are conserved among FS proteins, they are marked below with an asterisk (*). The seven amino acids that are shared between mouse cis-AB transferase and all FS proteins are highlighted. Crucial amino acids corresponding to codons 176, 235, 266, and 268 of human A/B/cis-AB transferases, as well as codon 296 of FS proteins are indicated in bold type. Murine cis-AB transferase has the same tripeptide sequence of GlyGlyAla(GGA) as that of FS proteins.
Evidently, mouse cis-AB transferase is more homologous to human A/B/cis-AB transferases than mouse and human FS proteins. Excluding the N-terminal sequences where alignment was difficult, there are only 7 amino acids that are uniquely conserved between mouse cis-AB transferase and mouse/human FS proteins, with two situated at codons 245 and 247 of mouse cis-AB transferase, which correspond to codons 266 and 268 of human A/B transferases. The tripeptide sequence of GlyGlyAla (GGA) at codons 245–247 of mouse cis-AB transferase is identical to the corresponding sequence at codons 261–263 of FS proteins. It should be noted that human cis-AB transferase coded by cis-AB01 allele possesses LeuGlyAla (LGA) and not GlyGlyAla (GGA).
Enhanced sensitivity of FS activity detection using COS1(B3GALNT1) cells as demonstrated by the re-examination of inactivating mutations in human GBGT1 gene
We compared the immunostaining results obtained using COS1 cells and those obtained using COS1(B3GALNT1) cells. Four expression constructs of FS proteins were analyzed. Results are shown in Fig. 2. Using COS1 cells, only the double revertant H_GBGT1(S230G&R296Q) construct exhibited FS activity. However, by using COS1(B3GALNT1) cells, not only H_GBGT1(S230G&R296Q) construct, but also H_GBGT1(R296Q) construct exhibited FS activity, suggesting that enhanced availability of globoside, the enzymatic reaction product of B3GALNT1 gene-encoded β1,3-N-acetyl-galactosaminyltransferase 1 and the acceptor substrate for FS, complemented the deficiency due to the Gly230Ser substitution in human GBGT1 gene. In other words, under the condition where a higher concentration of globoside is available, one amino acid substitution of Arg296Gln sufficiently confers human protein with FS activity.

Results of anti-Forssman antibody immunostaining after viral infection are compared between COS1 and COS1(B3GALNT1) cells. Cells were imaged using a Leica DMI6000B microscope equipped with a 20x/0.30 PH1 objective lens. One representative optical field was selected from individual experiments, and the cell pictures were taken in grey scale at the same magnification using monochrome DFC420 camera and Application Suite V3 software. All pictures are shown in 2 different magnifications. The right pictures correspond to the right bottom quarter of the left pictures at a higher magnification (2x). Figures were created using Adobe Photoshop software. The left panel shows the results using COS1 cells, and the right panel shows those using COS1(B3GALNT1) cells. The viral expression constructs examined are original human GBGT1 gene (H_GBGT1), human GBGT1 with Arg296Gln substitution (H_GBGT1-R296Q), human GBGT1 with Ser230Gly and Arg296Gln substitutions (H_GBGT1-S230G&R296Q), and human GBGT1 with Ser230Gly substitution (H_GBGT1-S230G) from top to bottom. Because the cells infected with the viral vectors were FACS-selected for their expression of coupled BFP and GFP, all the cells in a single experiment exhibited similar immunostaining results; positive or negative, depending on the constructs. Forssman antigen positivity is characterized not only by darker grey but also by cell surface DAB pigmentation.
Forssman glycolipid synthase activity of mouse ABO gene-encoded cis-AB transferase
As mentioned above, mouse ABO gene-encoded cis-AB transferase contains the GlyGlyAla tripeptide sequence, which is also found in the majority of GBGT1 genes. Therefore, we examined FS activity of mouse cis-AB transferase construct (M_ABO-AB), using COS1(B3GALNT1) cells with a higher globoside substrate concentration. In addition to mouse GBGT1 construct (M_GBGT1) as a positive control, we also analyzed human A, B, and cis-AB transferase constructs (H_ABO-A, H_ABO-B, and H_ABO-AB, respectively). Results of immunostaining are shown in Panel (I) of Table 1. We could detect cell surface expression of Forssman antigen on some of COS1(B3GALNT1) cells transfected with M_ABO-AB expression construct. Only the mouse cis-AB transferase with the GlyGlyAla tripeptide sequence, but not the human cis-AB transferase with the LeuGlyAla, exhibited FS activity equivalent of mouse GBGT1 gene-encoded FS.
Abolished FS activity of mouse GBGT1 gene-encoded FS and mouse ABO gene-encoded cis-AB transferase by substituting GlyGlyAla tripeptide with LeuGlyGly or MetGlyAla, and acquired weak FS activity of human A transferase by substituting LeuGlyGly with GlyGlyAla
We next prepared the following amino acid substitution constructs of ABO gene-encoded transferases by in vitro mutagenesis: M_ABO-AB(LGG), M_ABO-AB(MGA), and H_ABO-A(GGA). Similarly, we also prepared the substitution constructs of mouse GBGT1 gene-encoded FS: M_GBGT1(LGG) and M_GBGT1(MGA). The LeuGlyGly and MetGlyAla tripeptide sequences are found in A and B transferases from various species. Together with H_ABO-A, H_ABO-A(MGA), M_ABO-AB, and M_GBGT1, those constructs were transfected to COS1(B3GALNT1) cells. Results of immunostaining after DNA transfection are shown in Panel (II) of Table 1. Several photographs of the immunostained cells transfected with different constructs are shown for comparison in Fig. 3.

Results of anti-Forssman antibody immunostaining of COS1(B3GALNT1) cells after DNA transfection are shown. Cells were imaged in color using a Leica DMIL LED microscope equipped with D-LUX3 camera. All images were taken at the same magnification using a 10x/0.25 PH1 objective. Figures were created using Adobe Photoshop software. The eukaryotic expression constructs prepared in the pSG5 plasmid vector were used. The results are: mouse original GBGT1 gene (M_GBGT1) having GlyGlyAla tripeptide and its in vitro mutagenized amino acid substitution constructs with either LeuGlyGly or MetGlyAla tripeptide in the top row, mouse original ABO-AB gene (M_ABO-AB) with GlyGlyAla tripeptide and its substitution constructs having LeuGlyGly or MetGlyAla in the middle row, and human A allele of ABO gene (H_ABO-A) with LeuGlyGly tripeptide and its substitution constructs having GlyGlyAla or MetGlyAla tripeptide in the bottom row. Under the experimental conditions used, cells became confluent and formed a monolayer. Strong positive staining with anti-Forssman antibody was observed by DNA transfection of original M_GBGT1 and M_ABO-AB expression constructs. Additionally, COS1(B3GALNT1) cells transfected with H_ABO-A(GGA), the H_ABO-A construct with GlyGlyAla tripeptide sequence, exhibited some positive staining. It should be noted that not all cells were stained positive by DNA transfection as opposed to the viral infection-FACS sorting results shown in Fig. 2. Bluish background color was due to DAB and nickel ions in color development solution.
The substitution of GlyGlyAla sequence of mouse cis-AB transferase by either A transferase-specific LeuGlyGly or B transferase-specific MetGlyAla eliminated FS activity, suggesting that the GlyGlyAla tripeptide sequence is important for FS activity of mouse cis-AB transferase. The GlyGlyAla substitution of mouse FS by LeuGlyGly or MetGlyAla also abolished FS activity, indicating that this sequence is also important for FS activity of GBGT1 gene-encoded FSs. On the other hand, the substitution of LeuGlyGly of human A transferase by FS-specific GlyGlyAla conferred some FS activity although the activity was less than mouse FS or cis-AB transferase. In order to obtain more quantitative data, we also performed immunocytometry of the cells transfected with the selected constructs. After DNA transfection, cells were resuspended and immunostained. In combination with anti-Forssman antibody, Alexa Fluor 488-conjugated secondary antibody was used for fluorescence detection. Forssman antigen-positive cell percentages and Median Fluorescence Intensity (MFI) values of positive cell populations were determined. Results are shown in Table 2, confirming that some of the COS1(B3GALNT1) cells transfected with the H_ABO-A(GGA) construct, but not with the H_ABO-A construct, expressed Forssman antigen. The positive cell percentages, as well as the MFI values, of the H_ABO-A(GGA) construct were much lower than those of M_GBGT1 or M_ABO-AB. We, therefore, concluded that the GlyGlyAla tripeptide sequence alone is not sufficient for human A transferase to acquire a full FS activity. Some other determinants must be necessary.
Discussion
Immunochemical determination of glycolipid fractions of RBCs from individuals exhibiting Apae phenotype led to the discovery of a novel blood group system of FORS16. However, our results presented here suggest that the boundary between ABO and Forssman systems may not be as strictly delineated as previously thought.
Both ABO and GBGT1 genes are members of evolutionary related α1,3-Gal(NAc) transferase family of genes, deriving from the same ancestral gene29. ABO genes seem to have appeared during/after the separation of amphibians from fish. FUT1/FUT2/Sec genes encoding α1,2-fucosyltransferases that catalyze the last biosynthetic pathway of H substance, the acceptor substrate of A/B transferases, also seem to have emerged in amphibians. Accordingly, it is more logical to hypothesize that ABO gene-encoded A/B transferases have acquired the capability of utilizing H substance, a fucosylated acceptor substrate, than to assume that GBGT1 gene-encoded FSs have lost the ability to utilize fucosylated acceptor substrates. Even with this acquisition, however, ABO gene-encoded transferases may still retain some ability to utilize unfucosylated substrates if certain conditions are met. Obviously, the GlyGlyAla tripeptide sequence, which allows ABO gene-encoded glycosyltransferases to catalyze the biosynthesis of the FORS1 antigen, is such an example. The evolutionary link between ABO and GBGT1 genes and the structural similarity between A/B transferases and FSs make distinct blood group systems of ABO and FORS closely connected one another.
We have previously shown that two missense mutations resulting in Gly230Ser and Gln296Arg substitutions are responsible for inactivity of human GBGT1 gene-encoded FS protein27. Re-examination of those inactivating mutations using COS1(B3GALNT1) cells has revealed that Arg296Gln substitution alone might bestow human GBGT1 gene-encoded protein with FS activity strong enough to present what is called Apae phenotype under the condition where sufficient globoside concentration was provided by the forced expression of β1,3-N-acetyl-galactosaminyltransferase 1. Actually, an augmented FS activity by B3GALNT1 gene expression was previously reported by Svensson and colleagues, who observed a striking increase in Forssman antigen-positive cells after co-transfecting a hematopoietic cell line with B3GALNT1 gene construct and GBGT1 gene construct with Arg296Gln substitution16. Obviously, enhanced sensitivity of FS activity detection is not unique to COS1(B3GALNT1) cells. Extrapolating from this observation, deficiency in human GBGT1 gene-encoded FS protein caused by Gln296Arg substitution may also be overcome by currently unknown differential physiological/pathological conditions.
As mentioned earlier in the Introduction section, Forssman antigen has been detected in human cancer cells and tissues17,18,19,20,21,22,23. In addition to immunohistochemistry and immunocytometry, antigens reactive to anti-Forssman antibodies have also been detected by immunostaining of glycolipids separated through thin-layer chromatography17,23,31. One may question the specificity of anti-Forssman antibodies used in those experiments because polyclonal antibodies were used in many of the studies. However, in certain cases of human cancer Forssman glycolipid was demonstrated to exist by chemical determination of the antigenic structures18. It should be noted that in addition to regular pentasaccharide structure, Forssman antigen with unusual core structures, such as ceramide trisaccharide and a compound longer than a pentasaccharide ceramide, have also been detected in some tumors23,32. Similarly, Forssman antigens carried on glycoproteins seem to exist33,34,35.
The allele frequency of single nucleotide polymorphism (SNP) (rs375748588) responsible for the Apae phenotype is very low: 0.000008 in Exome Aggregation Consortium - ExAC Browser and 0.00008 in NHLBI Grand Opportunity Exome Sequencing Project (GO-ESP) database. Because this trait is dominant, the phenotype frequency of Apae may be of a similar value. Accordingly, this low allele frequency does not explain more frequent observation of Forssman antigen expression in cancer. COSMIC (Catalogue of Somatic Mutations in Cancer) database lists 60 mutations found in GBGT1 gene in various cancers, but none of them are missense mutations at codon 230 or 296. No SNPs have been found of the second nucleotide (T) of the triplet encoding Leu (CTG) or Met (ATG) at codon 266 of the ABO gene. Therefore, individuals with GlyGlyAla may be very rare. From regular A transferase with LeuGlyGly or B transferase with MetGlyAla, at least 3 or 2 nucleotide substitutions are necessary to change to GlyGlyAla, respectively, and actually, no mutations at the tripeptide sequence resulting in LeuGlyGly266–268GlyGlyAla or MetGlyAla266–268GlyGlyAla substitution have been reported of ABO gene in cancer, either. In summary, missense mutations at the tripeptide sequence are unlikely to explain Forssman antigen expression in cancer.
However, there are other potential mechanisms that remain to be examined. These include over-expression of GBGT1 gene mRNA and FS protein and increased stability of mRNA and protein. Splicing variations and post-translational protein modifications may also change the specificity and activity of FS protein. Even if no changes are observed in the structure and quantity of FS protein, extraordinarily higher concentrations of UDP-GalNAc and/or globoside substrates than threshold levels in the Golgi apparatus may also achieve the FS function. The finding that increased availability of globoside complemented the Gly230Ser deficiency supports this argument. Additionally, certain tumors were shown to contain higher concentrations of UDP-GalNAc and UDP-GlcNAc36. An increased level of intracellular UDP-GlcNAc was shown to activate O-GlcNAc transferase (OGT) and lead to enhanced O-GlcNAcylation of target proteins37. Therefore, a similar mechanism based on high donor nucleotide-sugar concentration may also influence the Forssman antigen expression. Forssman-positive cells tend to appear after DNA transfection on dish edges where cells are more crowded, suggesting that cell density may affect the antigen expression or antibody binding. Our results have proven that Forssman antigen might also be produced by other glycosyltransferases than FS. The observation that mouse ABO gene encodes a protein with FS activity warrants further exploration of the possibility that blood group ABO gene-encoded transferases may be involved in the biosynthesis of Forssman antigen in human cancer by yet-to-be defined molecular mechanisms.
Methods
Retrieval of ABO and GBGT1 gene/protein sequences and comparison of amino acid sequences of human A/B/cis-AB transferases, mouse cis-AB transferase, mouse Forssman glycolipid synthase (FS), and human FS proteins
The Mega file (Mouse_GBGT1_orthologues.med.txt) that aligned amino acid sequences of GBGT1 genes from a variety of species was downloaded from Ensembl database. The amino acid sequences of human A, B, and cis-AB transferases encoded by A101, B101, and cis-AB01 alleles at the ABO genetic locus were modified from the protein sequence of Ensembl ABO-202 transcript (ENST00000611156) by adding 3 nucleotides at the exon-intron splicing junction, which were missing in the annotated sequence, and then introducing allele-specific amino acid differences that were deposited at the dbRBC - BGMUT - System - ABO – NCBI database (http://www.ncbi.nlm.nih.gov/gv/rbc/xslcgi.fcgi?cmd=bgmut/systems_info&system=abo). The amino acid sequences of mouse cis-AB transferase, as well as human and mouse FS proteins, were retrieved from Ensembl, and directly used for sequence alignment, using ClustralW software of MEGA5 that was available online38.
Preparation of COS1(B3GALNT1) cells by retroviral introduction of human B3GALNT1 gene
African green monkey kidney COS1 cells were originally obtained from American Type Culture Collection (ATCC), and have been maintained in the laboratory. In order to improve the sensitivity of detection of FS activity, we created COS1(B3GALNT1) cells that produce increased number of globoside, the acceptor substrate of FS. For this purpose, we first prepared retroviral construct pMigR1b-B3GALNT1 by inserting human B3GALNT1 (β1,3-N-acetyl-galactosaminyltransferase 1) gene cDNA into pMigR1b (mTagBFP) vector at the cloning site preceding the internal ribosome entry sequence (IRES) element followed by blue fluorescent protein (BFP) gene cDNA. This vector construct was previously used to demonstrate the feasibility of preparing custom-made murine cells exhibiting specific glycolipid profiles by modular expression of extraneously introduced glycosyltransferase genes39. The pMigR1b-B3GALNT1 construct was transfected into packaging Phoenix-AMPHO cells. Then Phoenix-AMPHO medium containing the viral particles was recovered, filtered, and added to COS1 cells. The cells infected were allowed to grow to confluence, detached/suspended with trypsin/EDTA, and cells expressing coupled BFP were FACS-sorted. Those selected cells, COS1(B3GALNT1), were later used as hosts of subsequent viral infection and recipients of DNA transfection.
Preparation and infection of viral vectors of human GBGT1 gene and its derivatives
The expression constructs prepared in pSG5 (Stratagene) plasmid vector of human GBGT1 (H_GBGT1) and human GBGT1 with Ser230Gly, Arg296Gln, or both substitutions (H_GBGT1-S230G, H_GBGT1-R296Q, and H_GBGT1-S230G&R296Q, respectively), were previously described27. Those inserts were subcloned into pMigR1 (EGFP) viral vector having coupled enhanced green fluorescent protein (EGFP) gene cDNA, and the constructs were then transfected into Phoenix-AMPHO cells. The viral particles were produced and infected to COS1 and COS1(B3GALNT1) cells using the same strategy explained above. EGFP positive cells were selected of COS1 cells by FACS sorting. Double positive cells with EGFP and mTagBFP were selected of COS1(B3GALNT1) cells. The isolated populations were amplified and then seeded onto glass coverslips for Forssman antigen immunodetection.
Construction of in vitro mutagenized plasmid expression constructs
The constructs were prepared in pSG5. The original expression constructs of human A and B transferases (H_ABO-A and H_ABO-B), mouse cis-AB transferase (M_ABO-AB), and mouse FS (M_GBGT1), as well as human A transferase construct with Leu266Met and Gly268Ala substitutions (H_ABO-A(MGA)), were previously described26,27,28. The in vitro mutagenized constructs were prepared through a two-round PCR amplification using specific primers with nucleotide substitutions. The names and nucleotide sequences of primers, as well as detailed PCR protocols, can be found in Supplemental Information. The constructs without any additional unintended non-synonymous mutations were used in the following experiments.
DNA transfection
COS1(B3GALNT1) cells were used as recipients of DNA transfection. Lipofectamine 3000 reagents were used, following the manufacturer’s protocol (Life Technologies). For immunological detection of Forssman antigen by peroxidase staining procedure, two sets of expression constructs were DNA transfected: Set 1. Human A, B, and cis-AB transferase constructs (H_ABO-A, H_ABO-B, and H_ABO-AB, respectively), mouse cis-AB transferase construct (M_ABO-AB), and mouse GBGT1 construct (M_GBGT1). Set 2. Mouse cis-AB transferase construct (M_ABO-AB), mouse cis-AB transferase constructs with either GlyGlyAla245-247LeuGlyGly or GlyGlyAla245-247MetGlyAla substitution, M_ABO-AB(LGG) and M_ABO-AB(MGA), respectively, mouse GBGT1 construct (M_GBGT1), mouse GBGT1 constructs with either GlyGlyAla261-263LeuGlyGly or GlyGlyAla261-263MetGlyAla substitution, M_GBGT1(LGG) and M_GBGT1(MGA), respectively, and human A transferase construct (H_ABO-A), and human A transferase constructs with LeuGlyGly266-268GlyGlyAla or LeuGlyGly266-268MetGlyAla substitution, H_ABO-A(GGA) and H_ABO-A(MGA), respectively. In order to facilitate normalization of DNA transfection efficiency, a fixed amount of pEGFP construct expressing the enhanced GFP was co-transfected. GFP-positive cells were counted under the fluorescence microscope 3 days after DNA transfection just before cell fixation with 4% paraformaldehyde. The experiments were repeated 4 times for Set 2. For immunological detection of Forssman antigen by cytometry, only the selected expression constructs were DNA transfected: Set 3. H_ABO-A, H_ABO-A(GGA), M_ABO-AB, M_ABO-AB(LGG), M_GBGT1, and M_GBGT1(LGG).
Immunological detection of Forssman antigen
Immunostaining was performed using clone FOM-1 anti-Forssman Rat IgM monoclonal antibody from BMA Biomedicals. For immunoperoxidase staining procedure, cells fixed on culture plates were first incubated with the primary antibody, then with a biotinylated Goat Anti-Rat IgG + IgM(H + L) secondary antibody (Jackson ImmunoResearch Laboratories), followed by the treatment with Vectastain ABC reagents (Vector Laboratories). DAB reagents were used for peroxidase-mediated color development. Positive cell numbers were counted under the microscope and then normalized using cell numbers positive with co-transfected GFP. The values in percentage were then obtained, calculating the results of transfection with mouse FS construct (M_GBGT1) and a negative control to be 100 and 0, respectively. Pictures of the cells were also taken. For cytometry, DNA-transfected cells were detached from culture plates with EDTA, washed, and subjected to immunostaining, first with the anti-Forssman antibody and then with Alexa Fluor 488-conjugated Goat Anti-Rat IgM(μ Chain Specific) secondary antibody (Thermo Scientifics). BD LSRFortessa Analyzer (BD Biosciences) was used to determine Forssman antigen-expressing cell percentages and the B530-A Median Fluorescence Intensity (MFI) values of those expressing cell populations.
Additional Information
How to cite this article: Yamamoto, M. et al. Crosstalk between ABO and Forssman (FORS) blood group systems: FORS1 antigen synthesis by ABO gene-encoded glycosyltransferases. Sci. Rep. 7, 41632; doi: 10.1038/srep41632 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Schenkel-Brunner, H. Human blood groups: chemical and biochemical basis of antigen specificity. 2nd completely revised edn, (Springer-Verlag, 2000).
Daniels, G. Human blood groups. 2nd edn, (Blackwell Science, 2002).
Landsteiner, K. Zur Kenntnis der antifermentativen, lytischen und agglutinierenden Wirkungen des Blutserums und der Lymphe. Zentralblatt Bakteriol. 27, 357–362 (1900).
Landsteiner, K. Über Agglutinationserscheinungen normalen menschlichen Blutes. Wiener Klin Wochenschrift 14, 1132–1134 (1901).
Yamamoto, F., Clausen, H., White, T., Marken, J. & Hakomori, S. Molecular genetic basis of the histo-blood group ABO system. Nature 345, 229–233 (1990).
Yamamoto, F. et al. Molecular genetic analysis of the ABO blood group system: 4. Another type of O allele. Vox Sang. 64, 175–178 (1993).
Forssman, J. Die Herstellung hochwertiger spezifischer Schafhämonysine ohne Verwendung von Schafblut. Ein Beitrag zur Lehre von heterologer Antikörpebildung. Biochem Z. 37, 78–115 (1911).
Bushbinder, L. Heterophile phenomena in immunology. Arch Path. 19, 841–880 (1935).
Tanaka, N. & Leduc, E. H. A study of the cellular distribution of Forssman antigen in various species. J Immunol. 77, 198–212 (1956).
Siddiqui, B. & Hakomori, S. A revised structure for the Forssman glycolipid hapten. J Biol Chem. 246, 5766–5769 (1971).
Kijimoto, S., Ishibashi, T. & Makita, A. Biosynthesis of Forssman hapten from globoside by alpha-N-acetylgalactosaminyltransferase of guinea pig tissues. Biochem Biophys Res Commun. 56, 177–184 (1974).
Das, K. K., Basu, M., Basu, S. & Evans, C. H. Biosynthesis in vitro of a globoside containing a 2-acetamido-2-deoxy-beta-D-galactopyranosyl group (1–3)-linked and Forssman glycolipid by two N-acetylgalactosaminyltransferases from chemically transformed guinea pig cells. Carbohydr Res. 149, 119–135 (1986).
Haslam, D. B. & Baenziger, J. U. Expression cloning of Forssman glycolipid synthetase: a novel member of the histo-blood group ABO gene family. Proc Natl Acad Sci USA 93, 10697–10702 (1996).
Stamps, R., Sokol, R. J., Leach, M., Herron, R. & Smith, G. A new variant of blood group A. Apae . Transfusion 27, 315–318 (1987).
Svensson, L. et al. The structural basis of blood group A-related glycolipids in an A3 red cell phenotype and a potential explanation to a serological phenomenon. Glycobiology 21, 162–174 (2011).
Svensson, L. et al. Forssman expression on human erythrocytes: biochemical and genetic evidence of a new histo-blood group system. Blood 121, 1459–1468 (2013).
Kawanami, J. The appearance of Forssman hapten in human tumor. J Biochem. 72, 783–785 (1972).
Hakomori, S., Wang, S. M. & Young, W. W. Jr. Isoantigenic expression of Forssman glycolipid in human gastric and colonic mucosa: its possible identity with “A-like antigen” in human cancer. Proc Natl Acad Sci USA 74, 3023–3027 (1977).
Uemura, K., Hattori, H., Ono, K., Ogata, H. & Taketomi, T. Expression of Forssman glycolipid and blood group-related antigens A, Le(x), and Le(y) in human gastric cancer and in fetal tissues. Jpn J Exp Med. 59, 239–249 (1989).
Ono, K. et al. Expression of Forssman antigen in human large intestine. J Histochem Cytochem. 42, 659–665 (1994).
Yoda, Y., Ishibashi, T. & Makita, A. Isolation, characterization, and biosynthesis of Forssman antigen in human lung and lung carcinoma. J Biochem. 88, 1887–1890 (1980).
Mori, T. et al. Forssman antibody levels in sera of cancer patients. Immunol Commun. 11, 217–225 (1982).
Mori, E., Mori, T., Sanai, Y. & Nagai, Y. Radioimmuno-thin-layer chromatographic detection of Forssman antigen in human carcinoma cell lines. Biochem Biophys Res Commun. 108, 926–932 (1982).
Kominato, Y. et al. Animal histo-blood group ABO genes. Biochem Biophys Res Commun. 189, 154–164 (1992).
Yamamoto, F. & Yamamoto, M. Molecular genetic basis of porcine histo-blood group AO system. Blood 97, 3308–3310 (2001).
Yamamoto, M. et al. Murine equivalent of the human histo-blood group ABO gene is a cis-AB gene and encodes a glycosyltransferase with both A and B transferase activity. J Biol Chem. 276, 13701–13708 (2001).
Yamamoto, M., Cid, E. & Yamamoto, F. Molecular genetic basis of the human Forssman glycolipid antigen negativity. Sci Rep. 2, 975, doi: 10.1038/srep00975 (2012).
Yamamoto, F. & Hakomori, S. Sugar-nucleotide donor specificity of histo-blood group A and B transferases is based on amino acid substitutions. J Biol Chem. 265, 19257–19262 (1990).
Yamamoto, F. et al. An integrative evolution theory of histo-blood group ABO and related genes. Sci Rep. 4, 6601, doi: 10.1038/srep06601 (2014).
Yamamoto, F. & McNeill, P. D. Amino acid residue at codon 268 determines both activity and nucleotide-sugar donor substrate specificity of human histo-blood group A and B transferases: In vitro mutagenesis study. J Biol Chem. 271, 10515–10520 (1996).
Taniguchi, N., Yokosawa, N., Narita, M., Mitsuyama, T. & Makita, A. Expression of Forssman antigen synthesis and degradation in human lung cancer. J Natl Cancer Inst 67, 577–583 (1981).
Yokota, M., Warner, G. A. & Hakomori, S. Blood group A-like glycolipid and a novel Forssman antigen in the hepatocarcinoma of a blood group O individual. Cancer Res. 41, 4185–4190 (1981).
Mori, T., Mori, E., Sudo, T. & Kano, K. Expression of heterophil Forssman antigen as glycoprotein on transformed rat cell lines: shedding of the antigen from the cells. J Natl Cancer Inst. 73, 1179–1186 (1984).
Katagiri, T., Mori, T., Nakano, T., Ueno, K. & Kano, K. Aberrant expression of Forssman and Paul-Bunnell antigens on lymph node cells of MRL/Mp-lpr/lpr mice. J Immunol. 133, 3143–3148 (1984).
Kijimoto-Ochiai, S. et al. Forssman antigen expressed on lymph node cells of MRL/MpJ-lpr/lpr mice is of a glycoprotein nature. Microbiol Immunol. 34, 299–309 (1990).
Gribbestad, I. S., Petersen, S. B., Fjosne, H. E., Kvinnsland, S. & Krane, J. 1H NMR spectroscopic characterization of perchloric acid extracts from breast carcinomas and non-involved breast tissue. NMR Biomed. 7, 181–194 (1994).
Hart, G. W., Housley, M. P. & Slawson, C. Cycling of O-linked beta-N-acetylglucosamine on nucleocytoplasmic proteins. Nature 446, 1017–1022 (2007).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 28, 2731–2739 (2011).
Cid, E., Yamamoto, M., Buschbeck, M. & Yamamoto, F. Murine cell glycolipids customization by modular expression of glycosyltransferases. PLoS One 8, e64728, doi: 10.1371/journal.pone.0064728 (2013).
Acknowledgements
We are grateful to Silvia Rayo and David Izquierdo for their technical assistance. We also thank Marco Antonio Fernández Sanmartín and Gerard Requena Fernández at the IGTP Cytometry Core Facility for their help with the FACS sorting/analysis. The majority of work was performed at the now defunct Institut de Medicina Predictiva i Personalitzada del Càncer (IMPPC). The study was supported by the Spanish Health Research Foundation (Instituto de Salud Carlos III) grant (PI11/00454), the fund from Agència de GestiÓ d’Ajuts Universitaris i de Recerca (2014 SGR 1269), and the institutional start-up funds from IMPPC and IJC-“la Caixa” Foundation to F.Y. IJC and IGTP are CERCA centers supported by CERCA Programme/Generalitat de Catalunya. IJC is also supported by the Josep Carreras Foundation.
Author information
Authors and Affiliations
Contributions
M.Y. and F.Y. conceived and designed the experiment. M.Y. and E.C. performed the experiments. F.Y. wrote the manuscript with contributions from all the co-authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Yamamoto, M., Cid, E. & Yamamoto, F. Crosstalk between ABO and Forssman (FORS) blood group systems: FORS1 antigen synthesis by ABO gene-encoded glycosyltransferases. Sci Rep 7, 41632 (2017). https://doi.org/10.1038/srep41632
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep41632
This article is cited by
-
The stem region of group A transferase is crucial for its specificity, and its alteration promotes heterologous Forssman synthase activity
Scientific Reports (2023)
-
ABO blood group A transferase and its codon 69 substitution enzymes synthesize FORS1 antigen of FORS blood group system
Scientific Reports (2019)
-
Amino acid substitutions at sugar-recognizing codons confer ABO blood group system-related α1,3 Gal(NAc) transferases with differential enzymatic activity
Scientific Reports (2019)
-
Differential recognition of natural and remodeled glycotopes by three Diocleae lectins
Glycoconjugate Journal (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
