
Abstract
Microbial symbionts are known to assist exotic pests in their colonization of new host plants. However, there has been little evidence linking symbiotic invasion success to mechanisms for mitigation of native plant resistance. The red turpentine beetle (RTB) was introduced with a fungus, Leptographium procerum, to China from the United States and became a destructively invasive symbiotic complex in natural Pinus tabuliformis forests. Here, we report that three Chinese-resident fungi, newly acquired by RTB in China, induce high levels of a phenolic defensive chemical, naringenin, in pines. This invasive beetle-fungus complex is suppressed by elevated levels of naringenin. However, cryptic microbiotas in RTB galleries strongly degrade naringenin, and pinitol, the main soluble carbohydrate of P. tabuliformis, is retained in L. procerum-infected phloem and facilitate naringenin biodegradation by the microbiotas. These results demonstrate that cryptic microbiota mitigates native host plant phenolic resistance to an invasive symbiotic complex, suggesting a putative mechanism for reduced biotic resistance to symbiotic invasion.
Similar content being viewed by others


Introduction
Successful single-species invasions have been attributed to reduced biotic resistance in native communities1,2. Especially where a series of strong, nested interactions exists, biotic resistance can become a highly important determinant in their invasions3,4 as suggested by the ‘new associations’ or ‘increased susceptibility’ hypotheses5,6. Recent research highlights several cases of pest invasions that are facilitated by symbiotic microbes7,8. Although this form of invasion seems successful, the limitation of invaders under condition of biotic resistance is suggested to be more severe if their interdependent microbial symbionts are quenched9,10. Symbiotic invasion might be more susceptible to biotic resistance than single-species invasions. However, there has been little evidence linking symbiotic invasion success to mechanisms for reduced biotic resistance.
Studies have reported that compromised plant defences have contributed to the success of invasive insect pests on their new hosts11,12,13. Even though it is considered as an important barrier for colonization, the role of plant chemical defence as mediated by an invader’s microbial symbionts has received little attention. For coniferous trees, the main products of chemical defence in the bark are terpenoid resins and phenolics14. Volatile terpenoids are known to serve as olfactory cues for host recognition, mediate tritrophic interactions and act as precursors for bark beetle pheromone signalling15,16,17. Non-volatile terpenoids like diterpene resin acids are suggested to mainly inhibit fungal associates with no obvious negative effects on bark beetles18. Phenolics are also abundant and chemically diverse (e.g. monoaryls, flavonoids and stilbenes) in conifer bark19,20,21, within which compounds of the stilbenes are reported to have both the antifeedant and antifungal properties to bark beetles and blue-staining fungal associates22,23. Bark beetles are commonly associated with ophiostomatoid fungi24 and also with diverse bacterial and yeast species25,26. Conifer defensive chemicals were shown to be regulated by either type of these microbial associates27,28,29,30,31, yet how these symbiotic partners jointly mediate the bark beetle-conifer defence interactions has been little evidenced empirically.
The red turpentine beetle (RTB), Dendroctonus valens, was accidently introduced into China from the United States about three decades ago and, following a latency period, suddenly became a widespread tree killer, devastating forests and killing over ten million healthy Pinus tabuliformis trees32. Female pioneer beetles initiate attacks, construct galleries and perform monogamous behaviour that one male is attracted by one female for reproduction33,34; and during these processes, ophiostomatoid fungal associates are inoculated into host pines. The fungal community of Chinese RTB contained 20 ophiostomatoid fungal species, great majority (15 species) of which were not found associated with the US RTB and are herein referred to “Chinese-resident” fungi. The rest of fungi were associated with RTB both in China and US: Leptographium procerum is the primary fungal associate of Chinese RTB, which had all alleles shared by US RTB-associated L. procerum, with no allele unique in Chinese populations, this demonstrating that L. procerum associated with RTB in China originated from those with RTB in US and thus is referred to as “Chinese-invasive”; the other species that had no population genetic evidence yet are temporarily classified as “shared” fungal species. Overall, the fungal community composition has shifted from species associated with RTB in the United States to species newly acquired in China (See Supplementary Table S1).
The multispecies interactions between RTB, ophiostomatoid fungi and host pine P. tabuliformis have been suggested to facilitate invasion success. After their introduction into China, L. procerum evolved several novel genotypes that were more pathogenic to host pines and induced the release of higher amounts of 3-carene, a volatile terpenoid which is the primary attractant for the beetle vector35,36. RTB and L. procerum act as an invasive symbiotic complex in China32. However, L. procerum could be outcompeted by newly acquired Chinese-resident fungi37. From the benefit of induced rosin defence by resident fungi, their suppression of L. procerum was largely alleviated in the presence of the non-volatile terpenoid, diterpene resin acid37. Phenolics have the potential in targeting both beetles and ophiostomatoid fungi, and in return, it is fascinating to know in here how this invasive beetle-fungus complex could protect itself against this type of defensive chemical in pine phloem. Thus in this study, we first investigated whether fungal associates of RTB in China differentially induced phenolic resistance by conducting fungal inoculations on P. tabuliformis seedlings. Then, we assessed in vitro whether performance of this invasive symbiotic complex would be affected under increased phenolic resistance. Last, we tested whether harmful phenolic compounds could be decreased by associated bacteria or yeasts, symbionts that may exist cryptically in RTB gallery, through phenolic biodegradation.
Results
High concentrations of naringenin induced in P. tabuliformis seedlings by Chinese-resident fungi associated with D. valens
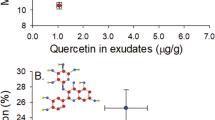
Among the Chinese-invasive, shared, and Chinese-resident fungal groups, one phenolic compound of the flavonoids, naringenin, showed significant induction by the Chinese-resident group, compared to controls 21 d after inoculation (Fig. 1 inset; Kruskal-Wallis test,  = 11.517, P < 0.01). Other phenolics including two monoaryls ρ-coumaric acid and ferulic acid, three stilbenes resveratrol, pinosylvin and pinosylvin monomethyl ether and another flavonoid taxifolin were not significantly induced by any fungal groups (Supplementary Table S2). Three Chinese-resident species, H. pinicola, L. truncatum, and L. sinoprocerum, among the representative 11 fungal species, induced significantly higher concentrations of naringenin compared to controls (Fig. 1; Kruskal-Wallis test,
= 11.517, P < 0.01). Other phenolics including two monoaryls ρ-coumaric acid and ferulic acid, three stilbenes resveratrol, pinosylvin and pinosylvin monomethyl ether and another flavonoid taxifolin were not significantly induced by any fungal groups (Supplementary Table S2). Three Chinese-resident species, H. pinicola, L. truncatum, and L. sinoprocerum, among the representative 11 fungal species, induced significantly higher concentrations of naringenin compared to controls (Fig. 1; Kruskal-Wallis test,  = 33.153, P < 0.001). The three Chinese-resident fungi and the Chinese-invasive L. procerum were then chosen for subsequent inoculation experiments.
= 33.153, P < 0.001). The three Chinese-resident fungi and the Chinese-invasive L. procerum were then chosen for subsequent inoculation experiments.

Among the ophiostomatoid fungal community, Chinese-resident fungi induce high concentrations of naringenin in pine seedlings.
Mean concentrations of naringenin (+SEM, n = 3–5) in 4-5-yr-old P. tabuliformis seedlings were obtained from inoculation with the 11 fungal associates of Chinese RTB after 21 d. Asterisks indicate significant differences between controls and isolates (*P < 0.05). Inset: Mean concentrations of naringenin (+SEM) induced by the Chinese-invasive, shared, and Chinese-resident fungal groups.
There were significant differences in naringenin concentrations 3, 6, 9, 12, 18, and 24 d after inoculation among all the treatments (Fig. 2A; two-way ANOVA; isolate, F4, 224 = 300.756, P < 0.0001; time, F5, 224 = 39.359, P < 0.0001; isolate × time, F20, 224 = 26.799, P < 0.0001). H. pinicola and L. truncatum induced higher concentrations of naringenin than mock inoculation 3, 6, 9, 12, 18, and 24 d after inoculation and L. sinoprocerum induced higher concentrations of naringenin than mock inoculation 6, 9, 12, 18, and 24 d after inoculation (Fig. 2A). However, there were no significant differences in naringenin concentrations between L. procerum and mock inoculation at all the sampled points except 6 and 12 d after inoculation (Fig. 2A). Neither were there significant inductions in the concentrations of other phenolic compounds by any fungal associates compared to controls (Supplementary Figs S1–S7).

Three Chinese-resident fungi induce high concentrations of naringenin over time and across stem diameters of pine seedlings.
(A) Mean concentrations of naringenin (±SEM, n = 7–10) from 4-5-yr-old P. tabuliformis seedlings inoculated with three Chinese-resident fungi and L. procerum after 3, 6, 9, 12, 18, and 24 d. Different letters indicate significant differences between treatments (P < 0.05). (B–E) Relationships between stem diameter and naringenin concentration in pine seedlings (n = 37–40) inoculated with three Chinese-resident fungi and L. procerum after 24 d.
Diameter of P. tabuliformis seedlings was a significant predictor of naringenin induction for the four fungal associates (Fig. 2B–E; H. pinicola, r2 = 0.486, F1, 38 = 35.958, P < 0.0001; L. truncatum, r2 = 0.343, F1, 35 = 18.295, P = 0.0001; L. sinoprocerum, r2 = 0.199, F1, 36 = 8.922, P = 0.005; L. procerum, r2 = 0.140, F1, 36 = 5.854, P = 0.021). The standardized slopes of regression between L. procerum and each of the three Chinese-resident fungi were significantly different (Fig. 2B–E; L. procerum, β = 0.374; H. pinicola, β = 0.697, F1, 74 = 32.94, P < 0.0001; L. truncatum, β = 0.586, F1, 71 = 6.81, P = 0.0111; L. sinoprocerum, β = 0.446, F1, 72 = 5.14, P = 0.0264).
Adverse effects of elevated naringenin on the invasive beetle-fungus complex
Female beetle tunnelling length was significantly reduced with elevated naringenin concentrations over 150 min (Fig. 3A; linear mixed-effects model test; treatment, F6, 118.520 = 14.364, P < 0.0001; time, F1, 156.073 = 48.084, P < 0.0001). Female body weight was significantly decreased with elevated naringenin concentrations after 6 h (Fig. 3B; one-way ANOVA, F5, 166 = 10.143, P < 0.0001). Larval beetles were significantly repelled from diet plugs with elevated naringenin concentrations after 6 h (Fig. 3C; 50 μg·g−1,  = 0.067, P = 0.796; 100 μg·g−1,
= 0.067, P = 0.796; 100 μg·g−1,  = 5.400, P = 0.020; 200 μg·g−1,
= 5.400, P = 0.020; 200 μg·g−1,  = 17.067, P < 0.0001; 500 μg·g−1,
= 17.067, P < 0.0001; 500 μg·g−1,  = 17.067, P < 0.0001; 1000 μg·g−1,
= 17.067, P < 0.0001; 1000 μg·g−1,  = 26.667, P < 0.0001; 2000 μg·g−1,
= 26.667, P < 0.0001; 2000 μg·g−1,  = 29.400, P < 0.0001). Larval feeding area (Fig. 3D; Kruskal-Wallis test,
= 29.400, P < 0.0001). Larval feeding area (Fig. 3D; Kruskal-Wallis test,  = 84.586, P < 0.0001) and body weight (Fig. 3E; two-way ANOVA; treatment, F6, 124 = 2.346, P = 0.035; block, F3, 124 = 6.559, P < 0.001) were significantly decreased with elevated naringenin concentrations after 6 h. Larval boring rate (Fig. 3F; Mantel-Cox test,
= 84.586, P < 0.0001) and body weight (Fig. 3E; two-way ANOVA; treatment, F6, 124 = 2.346, P = 0.035; block, F3, 124 = 6.559, P < 0.001) were significantly decreased with elevated naringenin concentrations after 6 h. Larval boring rate (Fig. 3F; Mantel-Cox test,  = 21.13, P < 0.0001) and survival rate (Fig. 3G; Mantel-Cox test,
= 21.13, P < 0.0001) and survival rate (Fig. 3G; Mantel-Cox test,  = 23.58, P < 0.0001) were significantly decreased with the elevated naringenin concentrations over 12 d.
= 23.58, P < 0.0001) were significantly decreased with the elevated naringenin concentrations over 12 d.

Elevated naringenin concentrations inhibit the beetle-fungus complex.
(A,B) Mean tunnelling length (+SEM, n = 18) and body weight change (+SEM, n = 26–30) of female RTB in relation to naringenin. Inset of (A) Mean tunnelling length (±SEM) over time. (C) Choice of RTB larvae (n = 60) for diet with only solvent or with naringenin. (D,E) Mean feeding area (+SEM, n = 22) and body weight change (+SEM, n = 18–20) of RTB larvae expose to naringenin. (F,G) Boring and survival rates of RTB larvae (n = 45) in the presence of naringenin. (H–K) Mean radial growth rates (+SEM, n = 4–6) of Chinese-resident fungi and L. procerum in the presence of naringenin. In (A,B,D,E), different letters indicate significant differences between treatments (P < 0.05). In (C,F–K), asterisks indicate significant differences between controls and treatments (*P < 0.05, **P < 0.01, ***P < 0.001).
The radial growth rate of each of the four fungal associates was significantly decreased with elevated naringenin concentrations (Fig. 3H–K; H. pinicola, one-way ANOVA, F6, 35 = 21.112, P < 0.0001; L. truncatum, one-way ANOVA, F6, 35 = 12.224, P < 0.0001; L. sinoprocerum, Brown-Forsythe’s one-way ANOVA, F6, 19.628 = 44.363, P < 0.0001; L. procerum, one-way ANOVA, F6, 21 = 51.393, P < 0.0001).
Naringenin biodegradation by cryptic microbiota with enhanced activity in pinitol, the main soluble carbohydrate of P. tabuliformis
Crude extracts from D. valens galleries caused significantly more naringenin biodegradation than those from healthy phloem after 72 h, but these differences were not significant after just 6 h (Fig. 4A; Kruskal-Wallis tests, 6 h,  = 7.079, P = 0.069; 72 h,
= 7.079, P = 0.069; 72 h,  = 23.184, P < 0.0001). Filtration through 0.22 μm filters significantly reduced naringenin biodegradation after 72 h (Fig. 4B; paired t test; t16 = 10.292, P < 0.001), indicating that naringenin biodegradation of D. valens gallery is conferred by cryptic microbiota. OD600 values and dry weights of gallery microbiotas with naringenin were significantly higher than those without naringenin (Fig. 4C; paired t test; t19 = −6.68, P < 0.001; Supplementary Fig. S8A; paired t test; t19 = −5.45, P < 0.001) when grown with pinitol, the main soluble carbohydrate in the phloem of P. tabuliformis (Supplementary Fig. S9).
= 23.184, P < 0.0001). Filtration through 0.22 μm filters significantly reduced naringenin biodegradation after 72 h (Fig. 4B; paired t test; t16 = 10.292, P < 0.001), indicating that naringenin biodegradation of D. valens gallery is conferred by cryptic microbiota. OD600 values and dry weights of gallery microbiotas with naringenin were significantly higher than those without naringenin (Fig. 4C; paired t test; t19 = −6.68, P < 0.001; Supplementary Fig. S8A; paired t test; t19 = −5.45, P < 0.001) when grown with pinitol, the main soluble carbohydrate in the phloem of P. tabuliformis (Supplementary Fig. S9).

Gallery microbiota degrades naringenin.
(A) Mean naringenin degradation activities (+SEM, replicates see Experiment 7 in Methods) of healthy phloem and galleries. (B) Mean quantities of naringenin (+SEM, n = 17) remaining after treatment with gallery crude extract with or without filtration. (C) Mean OD600 values (+SEM, n = 20) of gallery microbiota with or without naringenin in the presence of pinitol. (D) Mean concentrations of pinitol (±SEM, n = 7–10) in 4-5-yr-old P. tabuliformis seedlings inoculated with the three Chinese-resident fungi and the L. procerum after 3, 6, 9, 12, 18, and 24 d. (E,F) Mean OD600 values of gallery microbiota and quantities of naringenin (+SEM, n = 18) remaining after treatment with gallery microbiota with or without pinitol in the presence of naringenin. In (A,D), different letters indicate significant differences between treatments (P < 0.05). In (B,C,E,F), asterisks indicate significant differences between two treatments (*P < 0.05, ***P < 0.001).
There were significant differences of pinitol concentrations in the phloem of P. tabuliformis seedlings 3, 6, 9, 12, 18, and 24 d after inoculation (Fig. 4D; two-way ANOVA, isolate, F4, 234 = 144.864, P < 0.0001; time, F5, 234 = 51.939, P < 0.0001; isolate × time, F20, 234 = 6.328, P < 0.0001). L. procerum retained higher concentrations of pinitol than the three Chinese-resident fungi 24 d after inoculation (Fig. 4D; Supplementary Fig. S10). OD600 values and dry weights of gallery microbiotas with pinitol were significantly higher than those without pinitol (Fig. 4E; paired t test, t17 = −9.58, P < 0.001; Supplementary Fig. S8B; paired t test, t17 = −4.96, P < 0.001) when grown with naringenin. Gallery microbiotas with pinitol caused significant naringenin reduction compared to those without pinitol after 72 h (Fig. 4F; Wilcoxon signed ranks test, Z18 = −2.24, P = 0.025).
Fourteen bacterial and 8 yeast species were isolated from gallery microbiotas of 269 galleries (Supplementary Tables S3 and S4), within which 3 bacterial (Supplementary Fig. S11; Brown-Forsythe one-way ANOVA, F14, 33.939 = 1518.565, P < 0.0001) and 6 yeast species (Supplementary Fig. S12; Brown-Forsythe one-way ANOVA, F8, 4.597 = 1257.845, P < 0.0001) caused significant naringenin reductions. Among the 22 microbial species, 5 bacterial and 5 yeast species with pinitol caused significant naringenin reduction compared to the strain without pinitol after 72 h (Supplementary Fig. S13).
Discussion
Recent studies have demonstrated various essential roles of microbial symbionts for invasive pests in colonizing new host plants7,38,39,40, yet the dynamic interactions between invasive pest, microbial symbionts and new host plant appear to be more complicated than previously thought. In our system, we present evidence that the invasive RTB-L. procerum symbiotic complex appears to be challenged by host pine inducible phenolic resistance but this resistance can be mitigated by cryptic microbiotas in the beetle galleries. H. pinicola, L. truncatum and L. sinoprocerum, species responsible for the induced phenolic resistance, have not been reported as the fungal associates of RTB in the US, whereas seem to be unfortunately acquired by Chinese RTB populations41,42,43 (Fig. 5, lines 1.1–1.3; Supplementary Table S1). The ecological roles of flavonoids have been paid less attention than primarily studied stilbenes in bark beetle interactions; here, the increased naringenin present in host pines is shown to suppress the performance of both RTB and the fungal associate L. procerum (Fig. 5, line 2). This induced defensive flavonoid then, however, would be degraded by RTB gallery microbiota (Fig. 5, line 3.1). RTB populations might have tended to loosely associate with those resident fungi during invasion, considering that the invasive fungus L. procerum presents more cooperative characteristics with gallery microbiota than do the resident fungi (Fig. 5, lines 3.2–3.6). The association between naringenin-biodegrading microbiota and RTB-L. procerum symbiotic complex is quite close in the field (Supplementary Fig. S14, Tables S5 and S6), which implies the prevalence of naringenin-biodegrading microbiota that provides increased ability for this beetle-fungus complex to overcome host plant resistance (Fig. 5). Our study is the first step toward investigating possible mechanisms for reduced biotic resistance to symbiotic invasions, which could be a major contributing factor in the naturalization of an invasive pest system in the introduced ranges.

Complex interactions mediated by phytochemicals among host pine, red turpentine beetle, ophiostomatoid fungi and associated bacteria and yeasts.
1.1–1.3 (purple lines): Chinese-resident fungi acquired by RTB in China are inoculated to specifically induce naringenin in the phloem of host pines. 2 (red line): Naringenin suppresses the invasive RTB-L. procerum complex (in brown). 3.1–3.2 (blue lines): RTB gallery microbiota degrades naringenin, and naringenin stimulates the growth of gallery microbiota. 3.3–3.4 (blue lines): L. procerum is inoculated to induce retention of pinitol. 3.5–3.6 (blue lines): Pinitol enhances the growth and facilitates naringenin biodegradation of gallery microbiota. Dashed triangle (black line): A close tripartite symbiosis of RTB, L. procerum and gallery microbiota (Supplementary Fig. S14 and Table S6) enhances the invasion potential of the beetle-fungus complex in China.
Caution must be taken when making inferences on the ecological interactions in native pine forests, since these experiments were conducted with pine seedlings and beetle/symbionts bioassays in vitro. Although it was evident that naringenin concentrations were positively correlated to pine seedling stem diameters (or ages) (Fig. 2B–E), it is still unknown whether differential inductions of naringenin amongst fungi in P. tabuliformis seedlings can be extrapolated to those in mature pines; further, the potential combinatorial effects of these fungi that may synergize or antagonize naringenin induction remains unclear. In spite of good agreement between in vitro and in vivo experiments as shown in other bark beetle/fungus cases of phenolic resistances22,44,45, observations are needed on the performance of this beetle-fungus complex under elevated naringenin in mature P. tabuliformis in the future. Furthermore, this putative reduced biotic resistance should be linked to the time lag among introductions of RTB-L. procerum into new regions in China, its acquisition of resident fungal associates, its population dynamics and the compositional change of the cryptic microbiota in the field at larger temporal and spatial scales.
Microbial mediation of pine resistance may be more complicated than the results presented. Fungal infection of P. tabuliformis could reshuffle the biosynthesis of primary and secondary metabolites in the phloem. For example, in our results, the trade-off in biogenesis of pinitol (which is a cyclic polyol) and naringenin after the invasive/resident fungal infections potentially evidenced a connective pathway from cyclitols to phenolics in P. tabuliformis, which may be seen from another cyclitol, shikimic acid, that has been inferred in relation to production of many phenolics including naringenin in other systems46,47,48. The ecological role of gallery microbiota is rather intricate, especially when integrating terpenoid compounds into this work. Previous studies indicated the monoterpene 3-carene35,36 and diterpene resin acids37 appear to facilitate invasion success by the beetle-fungus complex; however, certain genera, e.g. Serratia and Rahnella in gallery microbiota of RTB were reported to metabolize monoterpenes and diterpene acids in another bark beetle system30. Compared to genera such as Burkholderia and Novosphingobium that were highly efficient in naringenin biodegradation (Supplementary Fig. S11), Serratia and Rahnella may specialize in terpenoid metabolism. Under this circumstance, it seems that gallery microbiota plays dual or multiple roles in processing pine defensive compounds, which is hard to absolutely define as “good” or “bad”. Given that terpenoids and phenolics mix in oleoresin, influence of terpenoids on microbial capacity in naringenin biodegradation needs to be evaluated in the future.
The discovery of identity and function of naringenin-biodegrading gallery microbiota went through three stages. First, utilizing RTB galleries collected from naturally infested forests, we found gallery tissues that exhibited higher naringenin-biodegrading activities after 72 h, compared to tissues from healthy phloem (Fig. 4A). Then, crude extracts from gallery tissues were filtered through 0.22 μm filters, which largely lost their capacity to degrade naringenin (Fig. 4B), revealing that naringenin degradation could be explained by microbial activities. Thus, we applied isolation method using naringenin as sole carbon source, an enrichment process for picking out naringenin-tolerant and naringenin-biodegrading species (Supplementary Table S5). Naringenin is commonly known to be antimicrobial49,50, whereas growth of the microbiota increased in response to this compound, possibly due to those microbes existing in RTB galleries (Fig. 4C). Bacteria and yeasts have been considered as novel partners in pine-bark beetle-ophiostomatoid fungi interactions25,51 and were also reported in RTB associated environments52,53,54. Many ecological functions of them have been disclosed; however, phenolic degrading microbes seem “cryptic” and their roles were seldom emphasized in bark beetle systems. For example, in this study, compared to naringenin-tolerant genera (e.g. Serratia, Pantoea and Raoultella etc.) that have often been found associated with RTB or other bark beetles, genera of microbiota capable of naringenin biodegradation (such as Burkholderia, Novosphingobium and yeast genus Rhodotorula) were previously regarded as minor ones. In another work, using community pyrosequencing, we found the frequency and abundance of Burkholderia and Novosphingobium were nearly equivalent to the more commonly seen genera (unpublished data). These “cryptic” genera may be of more importance in microbial ecology of RTB or even other bark beetles than previously thought.
Although naringenin-enrichment methods for microbial isolation helped us focus on particular microbes that potentially degrade naringenin, we did not always obtain highly effective species from the galleries that showed high biodegrading activities (Supplementary Table S6). This implied that traditional methods might overlook important microbes during the manual isolation process or that the production of high degrading activity in gallery microbiota might be more complicated than the actions of individual microbial species. The exact genes or metabolic pathways responsible for phenolics or even specifically for naringenin biodegradation that were operated by combinations of those microbes need to be studied in the future by metagenomics, an approach successfully applied in other bark beetle or insect systems to uncover the composition and functioning of their symbiotic microbiotas55,56.
The naringenin-biodegrading gallery microbiota may come partly from the RTB gut, since bacterial species of the genera Rahnella, Pseudomonas and Burkholderia as well as the yeast species Cyberlindnera americana and Kuraishia molischiana are frequently found in gut tissues from RTB and other bark beetle species26,52,53,55,56,57,58. The high naringenin-degrading Novosphingobium and Burkholderia have also been reported in oral secretions of bark beetles59, suggesting another possible source of the microbiota. Burkholderia and Pseudomonas found in the microbiota may also be vectored by RTB from associated environments, considering the presence of these genera in the pine rhizosphere60,61 and the behaviour of RTB to colonize and develop in the roots in addition to the main stem of pines34.
In general, despite the fact that there need more evidence to illustrate the mechanisms underlying reduced biotic resistance, our results presented a possible scenery that microbial symbionts functioned in such a diverse way for invasive insect pest through mediations of host plant resistance. Besides, this work involved an intricate cross-kingdom interactions (plant-insect-fungi-bacteria) showing more fascinating “warfare” between plant and herbivore than previously realized. Our findings also suggested that the full range of functions of microbes associated with invasive species should be considered in assessing the mechanisms underlying symbiotic invasion success.
Methods
Seedlings, fungal associates, and beetles
P. tabuliformis seedlings (diameter of 1-2-yr-old seedlings: Mean ± S.E = 5.87 ± 0.06 mm; diameter of 2-3-yr-old seedlings: Mean ± S.E = 7.08 ± 0.05 mm; diameter of 3-4-yr-old seedlings: Mean ± S.E = 8.09 ± 0.07 mm; diameter of 4-5-yr-old seedlings: Mean ± S.E = 9.99 ± 0.12 mm; measured at 2cm above the soil line) were cultivated in plastic pots (diameter: 12 cm) and transferred to a glasshouse (air temperature: 25 °C; relative humidity: 60%; 12 h photoperiod) for at least one month before the inoculation experiments.
The 11 fungal associates of D. valens used in this study were previously isolated from beetles or beetle galleries in infested P. tabuliformis forests in China and cultures were collected in the Forestry and Agricultural Biotechnology Institute, University of Pretoria and the Belgian Coordinated Collections of Microorganisms41,42. These fungal species (L. procerum from Chinese-invasive group; Ophiostoma floccosum, L. koreanum, O. abietinum and O. ips from shared group; O. rectangulosporium, O. minus, L. pini-densiflorae, L. sinoprocerum, L. truncatum and Hyalorhinocladiella pinicola from Chinese-resident group; Supplementary Table S1) accounted for 86.7% of total field isolations41,42,43.
Female D. valens beetles were collected from traps baited with D. valens kairomone lure (+)-(3)-carene in a naturally attacked forest at Tunlanchuan Forest Station (37°48′N, 111°44′E; average elevation 1400 m; P. tabuliformis trees: 3278 ha), Shanxi province. Second to third instar D. valens larvae were randomly collected from the beetle galleries of infested host pines at Tunlanchuan Forest Station.
Seedling inoculation and quantification of phenolic compounds
It is both ecologically dangerous and illegal (because of the National Natural Forest Protection Project in China) to operate inoculation experiments with invasive beetle and fungi or even artificial damages in uncontrolled, mature P. tabuliformis forests. Therefore, we chose to use P. tabuliformis seedlings in controlled conditions as a safe, legal alternative to large scale inoculations in natural forests. We performed single inoculation on one seedling in order to avoid excessive mechanical wound to the stem35,36,62,63. Although some studies showed ontogenetic differences in resistances between seedlings and mature plants64,65, other reports indicated that necrotic lesions from seedlings by several ophiostomatoid fungi are in agreement with those from mass-inoculated mature trees62,63. The tree size/fungal dosage ratio applied to mass-inoculated mature trees was found similar with that used in our P. tabuliformis seedlings (for example66, 20-yr-old unpruned P. sylvestris with 170 inoculations, the tree size/fungal inoculation ratio is 352, while the ratio we used as 4-5-yr-old seedling/a 5 mm diameter plug is 440).
To test the effect of fungi on pine phenolic defences and to select Chinese-resident fungi for subsequent experiments, inoculation assays were carried out using the eleven fungal associates of D. valens in China on 4-5-yr-old seedlings in May 2011. Seedlings received single inoculations with one of the fungal species or mock inoculation (n = 3 to 5 per treatment) applied by using a plug of 2% MEA alone (without fungi) on seedlings to serve as control. Infectious phloem materials (between 5 mm above and below inoculation point) were sampled at 21 d after inoculation (Experiment 1).
To test the effect of fungi on pine phenolic defences across time, in July 2011, we performed inoculation assays using Chinese-invasive fungus L. procerum and three Chinese-resident fungi (H. pinicola, L. truncatum and L. sinoprocerum) on 4-5-yr-old seedlings. The three Chinese-resident fungi accounted for 63.1% of field isolations of Chinese-resident group41,42,43. Seedlings received single inoculations with one of the four fungal species or mock inoculation (n = 7–10 per treatment in each time point). Infectious phloem materials were obtained at 3, 6, 9, 12, 18 and 24 d after inoculation (Experiment 2).
To test the effect of tree size on accumulation of naringenin, seedlings with ascending diameters across pine ages (1-2- to 4-5-yr-old seedlings) received single inoculations with one of the four fungal species (n = 8–10 per treatment in each pine age). Infectious phloem tissues were obtained at 24 d after inoculation (Experiment 3).
Before extraction, the phloem tissue was ground to powder in liquid N and washed with pentane (Sigma) to remove the resinous compounds. Phenolic compounds (two monoaryls p-coumaric acid and ferulic acid, two flavonoids taxifolin and naringenin and three stilbenes resveratrol, pinosylvin and pinosylvin monomethyl ether) were extracted from HPLC-grade methanol (JK Chemical Corporation, Beijing) and analyzed using HPLC (Agilent 1100, USA) as previously described67 with detailed procedures presented in Supplementary Methods Experiment 1. The phenolic polymer lignin was detected by spectrophotometry (Beckman Coulter, USA) and quantified by standard calibration curves with five different concentrations of lignin (TCI, Japan) according to the methods described68.
Bioassays on beetle and fungal associates under increased naringenin in vitro
In order to measure the effects of elevated levels of naringenin on beetle and fungal performances, we performed experiments on phloem media (for beetles) and MEA (for fungi) amended with naringenin to yield concentrations 0, 50, 100, 200, 500, 1000 and 2000 μg·g−1 of media dry weight, thus covering the concentration gradient induced by fungal associates from seedlings (Fig. 2).
To test tunnelling length of female D. valens under elevated levels of naringenin, phloem medium was prepared as described43 and then molten media amended with naringenin (dissolved in ethyl acetate) were transferred into a transparent glass tube (4 mm inner diameter, 50 mm in length) with enough space left at the open end for one beetle. Beetles were surface-sterilized and inserted head-down into each tube (n = 18 per treatment) then sealed with parafilm22. Tunnelling lengths were measured at half-hour intervals and longer tunnels indicate greater acceptance of substrate. To test body weight change of females, initial body weight of each beetle was recorded before insertion into tube containing naringenin amended phloem medium (n = 26–30 per treatment) and the ultimate body weight was recorded after 6 h (Experiment 4).
To test diet choice of D. valens larvae under elevated levels of naringenin, we placed one diet plug (diameter: 1.5 cm) with only solvent and the other with naringenin in a 35 mm Petri dish (two-choice bioassay). One sterilized larva was placed equidistant from the two plugs69. Petri dishes (n = 60 per treatment) were kept in 25 °C in darkness. Larvae were allowed to choose diet plugs and results were recorded after 6 h. In no-choice bioassays, to test body weight change of larvae (n = 18–20 per treatment), larvae were grouped into four blocks according to their initial weight, randomized into amended phloem media in respective blocks as one larva per Petri dish and reweighed after 6 h; to test feeding areas (n = 22 per treatment), larvae were removed from amended phloem media after 6 h and the feeding areas were photographed and measured using software Image J. It has been found that conifer phenolics have a clear antifeedant effect that acts quickly on bark beetle22, so we set experimental time of many assays as short as possible (within 6 hours); and we further tested the antifeedant effect of naringenin in longer time: to test long-term boring rate and survival (n = 45 per treatment), larval performances on phloem media amended with solvent (control) and two levels of naringenin (50 μg·g−1 and 2000 μg·g−1 of diet dry weight) were recorded every two days (Experiment 5).
To test radial growth rates of the four fungal associates under elevated levels of naringenin, we first dissolved naringenin in ethyl acetate and then mixed it into autoclaved 2% MEA to yield the appropriate naringenin concentrations. Ethyl acetate alone was used as control. Five mm fungal plugs were taken from the margin of actively growing fungal cultures and then placed on the centre of 90 mm diameter plates of MEA (n = 4–6 per treatment), ensuring that aerial mycelia were in contact with media36. Plates were incubated in the dark at 25 °C and growth was measured daily (Experiment 6).
Naringenin biodegradation
To test naringenin biodegradation by cryptic microbiotas in beetle galleries, we collected 111 D. valens galleries including 64 adult galleries, 18 egg galleries, 27 larval galleries, and 2 pupal galleries, from infested trees of a natural P. tabuliformis forest at Tunlanchuan Forest Station in July 2012. Samples were stored in sterile Eppendorf tubes at 4 °C and immediately transferred for experiments. Crude extracts (500 μl) containing putative microbiotas were obtained from one portion of the 111 gallery tissues and transferred to 1 ml of inorganic culture solution containing 1 mM naringenin as sole carbon source, methods referred to previously described70. Healthy phloem discs were also collected from 12 uninfested mature pines to see if putative innate phloem endophytes are responsible for naringenin biodegradation as well. After 6 h and 72 h of incubation, naringenin degraded by crude extracts of D. valens galleries and healthy phloem was detected and quantified by HPLC. To further determine that naringenin biodegrading activity is conferred by microbial symbionts distributed in galleries, crude extracts from 17 galleries were divided into one part with filtration through sterile 0.22 μm (pore size) filters and another part with no filtration. After 72 h of incubation, remaining naringenin was quantified by HPLC. Pinitol is a main soluble carbohydrate in the phloem of P. tabuliformis seedlings (Supplementary Fig. S9) and also in mature stems54. To test the effect of naringenin on the growth of gallery microbiota, crude extracts (40 μl) from 20 galleries were added in pinitol solutions (5 mM) with and without naringenin (1 mM). After 72 h of incubation at 30 °C, microbial solutions were transferred from glass culture tubes to 96 well flat-bottom microtiter plates to determine the final concentrations by measuring optical density at 600 nm (OD600) with a microplate reader (SpectraMax Plus, USA) and dry weights of the microbes were measured to the nearest microgram (μg) using an analytical balance (Mettler-Toledo, Switzerland) (Experiment 7).
To test the effect of the four fungi on pinitol retention in pine seedlings across time, we performed inoculation assays on 4-5-yr-old pine seedlings using the four fungal species (n = 7–10 per treatment in each time point) following procedures as described above, in August 2012. Pinitol was extracted from phloem tissues by methanol containing ribitol as internal quantitative standard, heated at 70 °C and separated by polar phase fractionation, followed by derivatization. Derivatized carbohydrates were identified and quantified by GC-MS and GC-FID (Agilent 7890A, USA) as described previously71 (Experiment 8).
To test the effect of pinitol on naringenin biodegradation of gallery microbiota, crude extracts (40 μl) from 18 galleries were added in naringenin solutions (1 mM) with and without pinitol (5 mM). After 72 h of incubation at 30 °C, final concentrations and dry weights of microbes were determined by measuring optical density at 600 nm and by using analytical balance respectively, and the remaining naringenin was quantified by HPLC (Experiment 9).
Detailed procedures for corresponding experiments mentioned above were presented in Supplementary Methods and additional information was placed into Supplementary Extended Methods.
Data analysis
Differences in two treatments were determined using t tests (Experiments 7 and 9). Differences of more than two groups of cases were analyzed by ANOVA (Experiments 1, 2, 4–8). For all t tests and ANOVA analyses, we tested the normal distribution (Kolomogorov-Simirnov test) and homogeneity (Levene’s test) of the variances of the responses for each treatment. Bonferroni post hoc tests were used for pair-wise comparisons following ANOVAs (Experiments 4 and 6). Datasets that did not conform to normality or had unequal variance were rank-transformed for one-way ANOVA (Experiment 4), one-way Brown-Forsythe’s ANOVA tested (Experiment 6), or non-parametric Kruskal-Wallis tested (Experiments 1, 2, 5, 7 and 8), with Student-Newman-Keuls post-hoc test, Dunnett’s T3 tests, or Mann-Whitney U tests using Bonferroni correction, respectively. The relationships between naringenin concentration and seedling diameter were examined by simple linear regression analysis (Experiment 3). Comparisons of regression coefficients between each of Chinese-resident fungi and the Chinese-invasive L. procerum were conducted by analysis of covariance. A significant interaction term between fungal associate inoculation and diameter indicated that the slopes of the two relationships differed significantly (Experiment 3). Preference behaviour of D. valens (Experiment 5) was tested using a chi-squared test. The survival and boring rate of larvae (Experiment 5) were calculated by Kaplan-Meier survival analysis (GraphPad Prism Software, Inc., San Diego, CA). All tests were carried out by using IBM SPSS 20 (SPSS Inc., Chicago, IL, USA) and SAS (SAS Institute Inc., Cary, NC, USA). Detailed statistics for corresponding experiments were presented in Supplementary Methods.
Additional Information
How to cite this article: Cheng, C. et al. Does cryptic microbiota mitigate pine resistance to an invasive beetle-fungus complex? Implications for invasion potential. Sci. Rep. 6, 33110; doi: 10.1038/srep33110 (2016).
References
Lyons, K. G. & Schwartz, M. W. Rare species loss alters ecosystem function–invasion resistance. Ecol. Lett. 4, 358–365 (2001).
Zavaleta, E. S. & Hulvey, K. B. Realistic species losses disproportionately reduce grassland resistance to biological invaders. Science 306, 1175–1177 (2004).
Levine, J. M., Adler, P. B. & Yelenik, S. G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 7, 975–989 (2004).
Kimbro, D. L., Cheng, B. S. & Grosholz, E. D. Biotic resistance in marine environments. Ecol. Lett. 16, 821–833 (2013).
Hokkanen, H. M. & Pimentel, D. New associations in biological control: theory and practice. Can. Entomol. 121, 829–840 (1989).
Colautti, R. I., Ricciardi, A., Grigorovich, I. A. & MacIsaac, H. J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 7, 721–733 (2004).
Himler, A. G. et al. Rapid spread of a bacterial symbiont in an invasive whitefly is driven by fitness benefits and female bias. Science 332, 254–256 (2011).
Vilcinskas, A., Stoecker, K., Schmidtberg, H., Röhrich, C. R. & Vogel, H. Invasive harlequin ladybird carries biological weapons against native competitors. Science 340, 862–863 (2013).
Parker, M. A. Mutualism as a constraint on invasion success for legumes and rhizobia. Divers. Distrib. 7, 125–136 (2001).
Nuñez, M. A., Horton, T. R. & Simberloff, D. Lack of belowground mutualisms hinders Pinaceae invasions. Ecology 90, 2352–2359 (2009).
Desurmont, G. A., Donoghue, M. J., Clement, W. L. & Agrawal, A. A. Evolutionary history predicts plant defense against an invasive pest. Proc. Natl. Acad. Sci. USA 108, 7070–7074 (2011).
Radville, L., Chaves, A. & Preisser, E. L. Variation in plant defense against invasive herbivores: evidence for a hypersensitive response in eastern hemlocks (Tsuga canadensis). J. Chem. Ecol. 37, 592–597 (2011).
Zavala, J. A., Casteel, C. L., Delucia, E. H. & Berenbaum, M. R. Anthropogenic increase in carbon dioxide compromises plant defense against invasive insects. Proc. Natl. Acad. Sci. USA 105, 5129–5133 (2008).
Franceschi, V. R., Krokene, P., Christiansen, E. & Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 167, 353–375 (2005).
Gijzen, M., Lewinsohn, E., Savage, T. J. & Croteau, R. B. Conifer monoterpenes: biochemistry and bark beetle chemical ecology in Bioactive Volatile Compounds from Plants (eds Teranishi, R., Buttery, R. G. & Sugisawa, H. ) 8–22 (American Chemical Society symposium series, 1993).
Payne, T. Olfactory basis for insect enemies of allied species in Potential for Biological Control of Dendroctonus and Ips Bark Beetles (eds Kulhavy, D. L. & Miller, M. C. ) 55–69 (Stephen F. Austin University Press, 1989).
Wood, D. L. The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annu. Rev. Entomol. 27, 411–446 (1982).
Kopper, B. J., Illman, B. L., Kersten, P. J., Klepzig, K. D. & Raffa, K. F. Effects of diterpene acids on components of a conifer bark beetle–fungal interaction: tolerance by Ips pini and sensitivity by its associate Ophiostoma ips. Environ. Entomol. 34, 486–493 (2005).
Pan, H. & Lundgren, L. N. Phenolic extractives from root bark of Picea abies. Phytochemistry 39, 1423–1428 (1995).
Pan, H. & Lundgren, L. N. Phenolics from inner bark of Pinus sylvestris. Phytochemistry 42, 1185–1189 (1996).
Viiri, H., Annila, E., Kitunen, V. & Niemelä, P. Induced responses in stilbenes and terpenes in fertilized Norway spruce after inoculation with blue-stain fungus. Ceratocystis polonica. Trees 15, 112–122 (2001).
Faccoli, M. & Schlyter, F. Conifer phenolic resistance markers are bark beetle antifeedant semiochemicals. Agric. For. Entomol. 9, 237–245 (2007).
Sallé, A., Monclus, R., Yart, A. & Lieutier, F. Effect of phenolic compounds on the in vitro growth of two fungi associated with Ips typographus. Forest Pathol. 35, 298–304 (2005).
Paine, T., Raffa, K. & Harrington, T. Interactions among scolytid bark beetles, their associated fungi, and live host conifers. Annu. Rev. Entomol. 42, 179–206 (1997).
Six, D. L. The bark beetle holobiont: why microbes matter. J. Chem. Ecol. 39, 989–1002 (2013).
Davis, T. S. The ecology of yeasts in the bark beetle holobiont: a century of research revisited. Microb. Ecol. 69, 723–732 (2015).
Hammerbacher, A. et al. A common fungal associate of the spruce bark beetle metabolizes the stilbene defenses of Norway spruce. Plant Physiol. 162, 1324–1336 (2013).
Raffa, K. F. & Smalley, E. B. Interaction of pre-attack and induced monoterpene concentrations in host conifer defense against bark beetle-fungal complexes. Oecologia 102, 285–295 (1995).
Brignolas, F. et al. Induced responses in phenolic metabolism in two Norway spruce clones after wounding and inoculations with Ophiostoma polonicum, a bark beetle-associated fungus. Plant Physiol. 109, 821–827 (1995).
Boone, C. K. et al. Bacteria associated with a tree-killing insect reduce concentrations of plant defense compounds. J. Chem. Ecol. 39, 1003–1006 (2013).
Mason, C. J. et al. Contrasting patterns of diterpene acid induction by red pine and white spruce to simulated bark beetle attack, and interspecific differences in sensitivity among fungal associates. J. Chem. Ecol. 41, 524–532 (2015).
Sun, J., Lu, M., Gillette, N. E. & Wingfield, M. J. Red turpentine beetle: innocuous native becomes invasive tree killer in China. Annu. Rev. Entomol. 58, 293–311 (2013).
Chen, H. F., Salcedo, C. & Sun, J. H. Male mate choice by chemicalcues leads to higher reproductive success in a bark beetle. Anim. Behav. 83, 421–427 (2012).
Yan, Z., Sun, J., Don, O. & Zhang, Z. The red turpentine beetle, Dendroctonus valens LeConte (Scolytidae): an exotic invasive pest of pine in China. Biodivers. Conserv. 14, 1735–1760 (2005).
Lu, M., Wingfield, M. J., Gillette, N. & Sun, J. H. Do novel genotypes drive the success of an invasive bark beetle-fungus complex? Implications for potential reinvasion. Ecology 92, 2013–2019 (2011).
Lu, M., Wingfield, M. J., Gillette, N. E., Mori, S. R. & Sun, J. H. Complex interactions among host pines and fungi vectored by an invasive bark beetle. New Phytol. 187, 859–866 (2010).
Cheng, C., Zhou, F., Lu, M. & Sun, J. Inducible pine rosin defense mediates interactions between an invasive insect-fungal complex and newly acquired sympatric fungal associates. Integr. Zool. 10, 453–464 (2015).
Adams, A. S. et al. Cellulose-degrading bacteria associated with the invasive woodwasp Sirex noctilio. ISME J. 5, 1323–1331 (2011).
Brown, A. M., Huynh, L. Y., Bolender, C. M., Nelson, K. G. & McCutcheon, J. P. Population genomics of a symbiont in the early stages of a pest invasion. Mol. Ecol. 23, 1516–1530 (2014).
Zhao, L. et al. A native fungal symbiont facilitates the prevalence and development of an invasive pathogen-native vector symbiosis. Ecology 94, 2817–2826 (2013).
Lu, M., Zhou, X., De Beer, Z., Wingfield, M. & Sun, J. Ophiostomatoid fungi associated with the invasive pine-infesting bark beetle, Dendroctonus valens, in China. Fungal Divers. 38, 133–145 (2009).
Lu, Q., Decock, C., Zhang, X. Y. & Maraite, H. Ophiostomatoid fungi (Ascomycota) associated with Pinus tabuliformis infested by Dendroctonus valens (Coleoptera) in northern China and an assessment of their pathogenicity on mature trees. Antonie Van Leeuwenhoek 96, 275–293 (2009).
Wang, B., Lu, M., Cheng, C., Salcedo, C. & Sun, J. Saccharide-mediated antagonistic effects of bark beetle funga lassociations on larvae. Biol. Lett. 9, 20120787 (2013).
Bois, E. & Lieutier, F. Resistance level in Scots pine clones and artificial introductions of Tomicus piniperda (Col., Scolytidae) and Leptographium wingfieldii (Deuteromycetes). J. Appl. Entomol. 124, 163–167 (2000).
Bois, E., Lieutier, F. & Yart, A. Bioassays on Leptographium wingfieldii, a bark beetle associated fungus, with phenolic compounds of Scots pine phloem. Eur. J. Plant Pathol. 105, 51–60 (1999).
Anderson, L. & Wolter, K. E. Cyclitols in plants: biochemistry and physiology. Annu. Rev. Plant Physiol. 17, 209–222 (1966).
Schijlen, E. G., Ric de Vos, C. H., van Tunen, A. J. & Bovy, A. G. Modification of flavonoid biosynthesis in crop plants. Phytochemistry 65, 2631–2648 (2004).
Tzin, V. & Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 3, 956–972 (2010).
Rauha, J. P. et al. Antimicrobial effects of Finnish plant extracts containing flavonoids and other phenolic compounds. Int. J. Food Microbiol. 56, 3–12 (2000).
Padmavati, M., Sakthivel, N., Thara, K. & Reddy, A. R. Differential sensitivity of rice pathogens to growth inhibition by flavonoids. Phytochemistry 46, 499–502 (1997).
Adams, A. S., Currie, C. R., Cardoza, Y., Klepzig, K. D. & Raffa, K. F. Effects of symbiotic bacteria and tree chemistry on the growth and reproduction of bark beetle fungal symbionts. Can. J. For. Res. 39, 1133–1147 (2009).
Lou, Q. Z., Lu, M. & Sun, J. H. Yeast diversity associated with invasive Dendroctonus valens killing Pinus tabuliformis in China using culturing and molecular methods. Microb. Ecol. 68, 397–415 (2014).
Xu, L., Lou, Q., Cheng, C., Lu, M. & Sun, J. Gut-associated bacteria of Dendroctonus valens and their involvement in verbenone production. Microb. Ecol. 70, 1012–1023 (2015).
Zhou, F. et al. Altered carbohydrates allocation by associated bacteria-fungi interactions in a bark beetle-microbe symbiosis. Sci. Rep. 6, 20135 (2016).
Adams, A. S. et al. Mountain pine beetles colonizing historical and naive host trees are associated with a bacterial community highly enriched in genes contributing to terpene metabolism. Appl. Environ. Microbiol. 79, 3468–3475 (2013).
Aylward, F. O. et al. Convergent bacterial microbiotas in the fungal agricultural systems of insects. MBio 5, e02077 (2014).
Adams, A. S., Adams, S. M., Currie, C. R., Gillette, N. E. & Raffa, K. F. Geographic variation in bacterial communities associated with the red turpentine beetle (Coleoptera: Curculionidae). Environ. Entomol. 39, 406–414 (2010).
Morales-Jimenez, J., Zuniga, G., Villa-Tanaca, L. & Hernandez-Rodriguez, C. Bacterial community and nitrogen fixation in the red turpentine beetle, Dendroctonus valens LeConte (Coleoptera: Curculionidae: Scolytinae). Microb. Ecol. 58, 879–891 (2009).
Cardoza, Y. J., Vasanthakumar, A., Suazo, A. & Raffa, K. F. Survey and phylogenetic analysis of culturable microbes in the oral secretions of three bark beetle species. Entomol. Exp. Appl. 131, 138–147 (2009).
Bent, E., Breuil, C., Enebak, S. & Chanway, C. P. Surface colonization of lodgepole pine (Pinus contorta var. latifolia [Dougl. Engelm.]) roots by Pseudomonas fluorescens and Paenibacillus polymyxa under gnotobiotic conditions. Plant Soil 241, 187–196 (2002).
Poole, E. J., Bending, G. D., Whipps, J. M. & Read, D. J. Bacteria associated with Pinus sylvestris-Lactarius rufus ectomycorrhizas and their effects on mycorrhiza formation in vitro. New Phytol. 151, 743–751 (2001).
Eckhardt, L., Jones, J. & Klepzig, K. D. Pathogenicity of Leptographium species associated with loblolly pine decline. Plant Dis. 88, 1174–1178 (2004).
Krokene, P. & Solheim, H. Assessing the virulence of four bark beetle-associated bluestain fungi using Norway spruce seedlings. Plant Pathol. 47, 537–540 (1998).
Barton, K. E. & Koricheva, J. The ontogeny of plant defense and herbivory: characterizing general patterns using meta-analysis. Am. Nat. 175, 481–493 (2010).
Boege, K., Dirzo, R., Siemens, D. & Brown, P. Ontogenetic switches from plant resistance to tolerance: minimizing costs with age? Ecol. Lett. 10, 177–187 (2007).
Solheim, H., Långström, B. & Hellqvist, C. Pathogenicity of the blue-stain fungi Leptographium wingfieldii and Ophiostoma minus to Scots pine: effect of tree pruning and inoculum density. Can. J. For. Res. 23, 1438–1443 (1993).
Bonello, P. & Blodgett, J. T. Pinus nigra-Sphaeropsis sapinea as a model pathosystem to investigate local and systemic effects of fungal infection of pines. Physiol. Mol. Plant Pathol. 63, 249–261 (2003).
Bonello, P. & Pearce, R. Biochemical defence responses in primary roots of Scots pine challenged in vitro with Cylindrocarpon destructans. Plant Pathol. 42, 203–211 (1993).
Cipollini, D., Stevenson, R., Enright, S., Eyles, A. & Bonello, P. Phenolic metabolites in leaves of the invasive shrub, Lonicera maackii, and their potential phytotoxic and anti-herbivore effects. J. Chem. Ecol. 34, 144–152 (2008).
Kikuchi, Y. et al. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 109, 8618–8622 (2012).
Lisec, J., Schauer, N., Kopka, J., Willmitzer, L. & Fernie, A. R. Gas chromatography mass spectrometry–based metabolite profiling in plants. Nat. Protoc. 1, 387–396 (2006).
Acknowledgements
We are especially grateful to Drs. Nancy Gillette (PSW Research Station, USDA Forest Service), Jacob Wickham (Institute of Chemistry, CAS), Kathryn Bushley (University of Minnesota), Daniel Miller (Southern Research Station, USDA Forest Service) and two anonymous reviewers for their valuable comments which improved greatly the quality of this paper. We also thank Mrs. Xiaowei Qin (Institute of Zoology, CAS) for assistance in chemical analyses. This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB11050000) and the National Natural Science Foundation of China (31110103903, 31222013 and 31170610).
Author information
Authors and Affiliations
Contributions
The study was jointly conceived by C.C., M.L. and J.S. Experiments were designed by C.C., M.L. and J.S.; C.C. prepared the manuscript; M.L., J.S., L.X., D.X. and Q.L. edited the manuscript. C.C., M.L., L.X., D.X. and Q.L. carried out the experiments. Figure 5 in this manuscript was originally drawn by the senior author, Mr. Chihang Cheng.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Cheng, C., Xu, L., Xu, D. et al. Does cryptic microbiota mitigate pine resistance to an invasive beetle-fungus complex? Implications for invasion potential. Sci Rep 6, 33110 (2016). https://doi.org/10.1038/srep33110
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep33110
This article is cited by
-
Phylogeny of Leptographium qinlingensis cytochrome P450 genes and transcription levels of six CYPs in response to different nutrition media or terpenoids
Archives of Microbiology (2022)
-
An invasive beetle–fungus complex is maintained by fungal nutritional-compensation mediated by bacterial volatiles
The ISME Journal (2020)
-
Bacterial and Fungal Midgut Community Dynamics and Transfer Between Mother and Brood in the Asian Longhorned Beetle (Anoplophora glabripennis), an Invasive Xylophage
Microbial Ecology (2019)
-
Changes in the Microbial Community of Pinus arizonica Saplings After Being Colonized by the Bark Beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae)
Microbial Ecology (2019)
-
Bacterial microbiota protect an invasive bark beetle from a pine defensive compound
Microbiome (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
