
Abstract
To test whether there are general patterns in biomass partitioning in relation to environmental variation when stand biomass is considered, we investigated biomass allocation in leaves, stems and roots in China’s forests using both the national forest inventory data (2004–2008) and our field measurements (2011–2012). Distribution patterns of leaf, stem and root biomass showed significantly different trends according to latitude, longitude and altitude and were positively and significantly correlated with stand age and mean annual precipitation. Trade-offs among leaves, stems and roots varied with forest type and origin and were mainly explained by stand biomass. Based on the constraints of stand biomass, biomass allocation was also influenced by forest type, origin, stand age, stand density, mean annual temperature, precipitation and maximum temperature in the growing season. Therefore, after stand biomass was accounted for, the residual variation in biomass allocation could be partially explained by stand characteristics and environmental factors, which may aid in quantifying carbon cycling in forest ecosystems and assessing the impacts of climate change on forest carbon dynamics in China.
Similar content being viewed by others
Introduction
The leaves of forest plants perform photosynthesis, stems (including boles and branches) provide mechanical support and a hydraulic pathway and roots absorb nutrients and water and provide anchorage1,2. The allocation of biomass among leaves, stems, roots and reproductive tissues can be influenced by plant size, growth environment and species3,4. A quantitative understanding of such patterns is of fundamental importance to understanding plant ecology and evolution and biomass patterns have implications for global climate research as well as many uses in forest practice and management2,5,6. Although large data sets within and across the broad spectrum of vascular plant species have been used to evaluate patterns of standing biomass7,8,9,10,11, theoretical or empirical assessments of these patterns remain contentious12,13,14,15,16,17,18. Thus, quantitative assessments of forest biomass allocation patterns remain central to forest biology and ecology.
Optimal partitioning theory (OPT), which is based on the assumption that there are trade-offs in resource allocation between leaf, stem and root functions, has emphasized the variability in biomass allocation observed across plant species, suggesting that plants allocate biomass to the organ that acquires the most limiting resource19,20,21. Broad-scale biomass observations in vascular plants have supported the existence of OPT22,23,24, but some findings have indicated that variation in biomass allocation may be driven by differences in plant size5,21,25. To resolve this question, allometric biomass partitioning theory (APT), was developed to predict how plants allocate biomass in leaves, stems and roots, based on the constraints of stand biomass21. Although analysis of a large collection of standing organ biomass, sampled across a broad spectrum of taxa from diverse ecological habitats, supports the relationships predicted by the model for leaf, stem and root biomass allocation, the APT has also been questioned6,13,14. Therefore, it remains unclear whether general patterns in biomass partitioning exist that are linked to environmental variation after plant size is accounted for.
In China, the country with the fifth-largest forest area worldwide, multiple combinations of complex forest characteristics (e.g., age, density, forest type and forest origin [primary, secondary and planted forest]) as well as variable environmental factors (e.g., temperature, precipitation, soil nutrients and topography)26, offer a unique opportunity to examine patterns of biomass allocation across diverse forest ecosystems. In addition, forest characteristics and environmental factors are thought to account for a large proportion of forest carbon stocks27,28,29. Obtaining a better understanding of the influence that these factors have on biomass allocation is fundamentally important for developing environmental policies and ecosystem carbon management practices to enhance the forest carbon sink. However, despite the progress made to date, large-scale patterns of biomass allocation in leaves, stems and roots based on stand biomass constraints are not well quantified in China’s forests.
In this study, we investigated the biogeographic patterns of biomass allocation in leaves, stems and roots based on a large-scale biomass survey across forest communities in China. We explored the following questions: (1) Does leaf, stem and root biomass vary with latitude, longitude and altitude gradients in forests? (2) Are trade-offs in biomass allocation mainly controlled by total tree biomass? (3) Are these patterns of biomass allocation also related to forest stand characteristics and large-scale environmental differences? Based on previous studies, we hypothesized that (1) leaf, stem and root biomass would show significant biogeographic patterns, since the ability of forest plants to utilize available energy is limited by water and temperature conditions30,31,32; (2) the relative importance of tree size (biomass) to trade-offs in biomass allocation would be greater than that of other factors5,25; and (3) after constraints imposed by tree size were accounted for, variation in patterns of biomass allocation would be modulated by forest stand characteristics and environmental factors14,25. Exploring these important questions and hypotheses is essential for quantifying carbon cycles in forest ecosystems and assessing the impacts of climate change on forest carbon dynamics in China6,10,33.
Results
Statistics and biogeographic patterns of forest biomass allocation
Tree components exhibited large variation in biomass across sampling sites (Fig. 1), ranging from 0.42 to 38.72 Mg ha−1 for leaves, 12.97 to 552.86 Mg ha−1 for stems and 1.48 to 232.53 Mg ha−1 for roots (Fig. 2), with mean values of 7.82, 121.16 and 30.20 Mg ha−1, respectively and a ratio of 6.5:100:24.9. The biomass of each component varied markedly across different forest types (Table 1). However, leaf and root biomass did not differ significantly according to forest origin (Table 1).

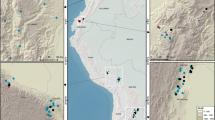
Locations of the 1,022 sampling sites across the forests of China.
(The map is made by ArcGIS 10.2 software, http://www.arcgis.com/features/).

Frequency distributions of biomass in China’s forests.
The mean and median values of the frequency distribution of biomass in (a) leaves, (b) stems and (c) roots are presented.
The tree components revealed significantly different biogeographic trends (P < 0.01; Table 2). Leaf biomass increased significantly from west to east and from south to north, while stem and root biomass showed the opposite pattern. In addition, stem and root biomass increased significantly with increasing altitude, but leaf biomass showed no significant altitudinal trend.
Factors influencing leaf, stem and root biomass in China’s forests
SMR indicated that leaf, stem and root biomass was significantly and positively correlated with AGE and MAP (Table 3), while leaf biomass was significantly and negatively correlated with TN in soil. Stem and root biomass were positively and significantly correlated with MAT and MAXT, respectively. MAP explained 13.1% of the variation in leaf biomass, while MAP and AGE combined accounted for 27.2% of this variation. However, explanation of the variation in leaf biomass was improved by only 0.9% when MAP, AGE and TN were combined. AGE explained 33.8% and 22.0% of the variation in stem and root biomass, respectively. Together, AGE and MAT accounted for 51.2% of the variation in stem biomass, while similarly, 31.3% of the variation in root biomass was explained by the combination of AGE and MAP.
Relationship of forest biomass allocation to stand characteristics and environmental factors
The PCA demonstrated that 75.5%, 16.6% and 7.9% of variation in leaf, stem and root biomass in all forests could be explained by the first, second and third principal components (PC1, PC2 and PC3), respectively. Although the explanatory power of the PCs varied with forest type and origin, more than 64.5% of the variation in leaf, stem and root biomass was accounted for by PC1 and more than 7.7% by PC2 (Table 4).
The PC axes contained the trade-offs between the biomass of each component, as reflected in the loadings (Table 5). For all forests, PC1 contained equal loadings for leaf, stem and root biomass, suggesting there was variation in total stand biomass. Compared with PC1, both PC2 and PC3 loaded positively for leaf biomass and negatively for stem and root biomass, revealing a trade-off between photosynthetic and non-photosynthetic allocation (Table 5). In addition, PC1 had similar loadings for leaf, stem and root biomass across 11 forest types and three forest origins, while loading trends in PC2 and PC3 varied with forest type and origin (Table 5).
PC1 was positively correlated with age and MAP, which demonstrated that total tree biomass increased with stand age and precipitation (Table 6). Similarly, PC2 was negatively correlated with age and positively correlated with MAP and MAXT, indicating that leaf biomass decreases with stand age and increases with precipitation and maximum temperature in the growing season, at the expense of stem and root biomass. PC3 correlated positively with density and MAT and negatively for age (Table 6), showing that leaf biomass increased with stand density and MAT at the expense of stem and root biomass. In contrast, stem and root biomass increased with age at the expense of leaf biomass. Additionally, the GLM indicated that the three PCs were closely related to forest type and origin (Table 7), which suggested that total biomass and biomass allocation were influenced by these factors.
Discussion
In this study, the distribution of leaf, stem and root biomass was examined and we revealed a large variation among forests in China (Fig. 2, Table 1). Leaf biomass showed significant positive longitudinal trends while it also was characterized by significant negative latitudinal trends. Stem and root biomass revealed significant negative longitudinal trends, as well as significant positive latitudinal trends (Table 3). These results support our hypothesis that different underlying biophysical (environmental) and biological controls shape the biogeographic patterns of biomass allocation. Climatically, the north-to-south and west-to-east gradients in China reflect shifts from cold and dry to warm and moist conditions and as might be predicted, the thermal gradient is steeper from north to south and the moisture gradient more pronounced from west to east34,35. Compared with the lack of significant altitudinal trends for leaves, both the stem and root biomass demonstrated a significant positive altitudinal trend. This phenomenon appears to be consistent with the latitudinal trends since temperature decreases as latitude and altitude increase. However, these biogeographic patterns of forest biomass allocation are also associated with geographic patterns in the structure and function of terrestrial ecosystems (e.g., stand age, forest type, biodiversity, soil development, primary production and plant ecological traits)36,37,38, which also reflect responses to climate gradations and site conditions.
Although previous studies on OPT examined trends in biomass allocation and resource availability and indicated that this theory is generally correct, others have demonstrated that plant ontogeny has a strong effect on allocation patterns39,40,41. During shifts in plant developmental stages, more investment in support tissues is expected as plants increase in size5. Therefore, to understand how plants actively alter assimilation partitioning in response to changes in the environment, plant size must be considered41. Here, trade-offs between leaves, stems and roots were mostly explained by PC1 (Table 4), which suggests that variation in biomass allocation is closely related to stand biomass, consistent with our hypothesis and APT.
Forest origin plays an important role in shaping biomass allocation and diversity–productivity relationships38,42. The differences in biomass allocation among natural, secondary and planted forests may highlight the importance of the impact of forest age. After stand biomass was constrained, most secondary forests are in early successional stages, which were reflected in relatively high stem biomass, as well as low root biomass and low total forest biomass, while primary forests in mature stages had relatively low stem biomass, high root biomass and high total forest biomass (Tables 5 and 6). Surprisingly, if stand biomass was not constrained, there was no obvious difference in trade-offs between stems and roots in primary and secondary forests (Table 1), indicating that the role of forest origin in forest biomass allocation in China may be misinterpreted if the stand biomass is not considered32. In addition, after stand biomass was accounted for, trade-offs between leaves, stems and roots differed between forest types (Table 5), similar to what was previously reported13,14,43.
Here, we also demonstrate that leaf and root biomass increases and stem biomass decreases with stand density (Tables 4, 5, 6). Zhang et al. (2012) found that stem, branch and leaf biomass varied with stand density across eight different forest types in China44. Some studies have also indicated that resource use is influenced by forest density. For example, due to constraints on stomatal conductance, the photosynthetic rate per unit leaf area was negatively correlated with forest density under hot, dry conditions, suggesting that stem biomass would increase in order to reduce the risk of cavitation45.
Our findings indicate that in all forests, leaf and root biomass increase or decrease more markedly depending on MAT or MAXT than does stem biomass (Tables 4, 5, 6). Different responses in forest biomass allocation to temperature may be partly modulated by turnover rate. Leaves and roots have higher turnover rates than stems and are more likely to contribute to biogeographic variation in biomass distribution6. Reich et al. (2014) found that forests probably allocate a smaller proportion of new biomass to foliage in cold climates since there are low turnover rates6. Although there is uncertainty in estimates of root biomass as a result of sampling methods and spatial heterogeneity in soil, root turnover, like leaf turnover, is probably slower in higher-latitude, or higher-altitude environments30,46. Furthermore, forests in high-latitude and high-altitude conditions are influenced by low temperature, low water availability and low nutrient supply47,48,49,50. Therefore, both cold temperatures and low levels of soil resources contribute to the patterns of greater root biomass associated with low MAT6.
Our data revealed that leaf biomass was significantly and negatively correlated with TN in soil (Table 3), which is not consistent with the other studies51,52. However, after the stand biomass was considered, the leaf biomass was not significantly correlated with soil nitrogen (Table 6), suggesting the soil nitrogen was not the main factor influencing leaf biomass allocation5,6. In addition, plant response to water deficit or drought may vary with the severity of water stress53. Root biomass increases little under moderate water stress54 but increases dramatically at the expense of stems when plants are subjected to severe drought. Still, biomass allocation may not respond strongly to a drought of relatively short-duration. In this study, forest root biomass increased with decreasing MAP (Tables 4, 5, 6), which is consistent with previous research. Compared with root biomass, some studies have indicated that leaf biomass changes little with increasing drought45, while others found that leaf biomass increased or decreased under different drought-stress conditions53,56. Our finding that after stand biomass is considered, relative leaf biomass decreased with increasing MAP (Tables 4, 5, 6), which is consistent with some reports53.
Moreover, there is a significant uncertainty related to patterns of forest root biomass based on the known aboveground biomass and average root/stem ratios derived from the literature in China10. A quantitative understanding of the ways in which root biomass distribution is influenced by stand characteristics and environmental factors is essential for improving the accuracy of carbon inventories and simulation modeling under global climate change57,58. As described above, our results suggest that, after stand biomass is accounted for, residual variation in biomass allocation could be partially explained by stand characteristics and environmental factors, which may aid in quantifying carbon cycling in forest ecosystems and assessing impacts of climate change on forest carbon dynamics in China7,8,9,10,11,59. In addition, these relationships will help to predict the influence of climate warming, land-use change and forest thinning on ecosystem carbon of forest vegetation. For example, our observation of higher stem biomass and lower leaf and root biomass under conditions of low stand density implies that high stand density (heavily managed) may lead to slow-growth in new stands, fast self-thinning in mature forests and low rate of wood production. The mechanisms underlying these ecological consequences, which may involve phenotypic plasticity, tissue turnover and resource supply, remain unclear2,30. In the future, we hope that the integration of regional inventory data, long-term monitoring data and controlled experiments will provide answers to these important questions.
Methods
Large-scale forest biomass data
We used forest biomass data from 1022 sites across China, including 110 sites at which we performed field measurements from 2011 to 2012 and 912 sites from the national forest inventory data set (2004–2008) (Fig. 1). In total, 1022 observations of each tree component (leaf, stem, root and whole tree) were included in the dataset. In detail, three replicate 20 × 50 m plots were established at each sampling site in our field survey and the national forest inventory. Each plot was divided into ten 10 × 10 m quadrats, with the height and diameter at breast height (DBH) of each tree and the total number of trees in each plot recorded.
Based on height and DBH, five to seven trees of different diameter classes were selected from each species for measurement of tree components biomass. For tree root biomass, coarse roots of the selected trees were dug up, washed, separated by hand into size classes based on diameter (≤2 mm and >2 mm) and weighed to obtain a measure of fresh biomass. Total coarse biomass was estimated by adding together the weights of all root sizes. Fine roots were sampled using the soil block sampling method30. Root cores were taken at three random points around each sampled tree at different depths (mostly ≥50 cm). After soil was washed from the roots, fine roots (≤2 mm diameter) were separated from coarse roots (>2 mm) and dried at 65 °C until a constant weight was obtained. Representative root samples were taken to the laboratory, dried and used to calculate the relationship between dry and fresh weight. Root biomass was then estimated using the regression models considering DBH (see Appendix Table S1 for details)8,29,60. Similarly, the leaf, bole and branch biomass of the selected trees were separated and weighed, respectively. The representative fresh samples were also dried in the laboratory to calculate the relationship between dry and fresh weight. The biomass of the leaf, bole and branch were then also estimated using the regression models that accounted for DBH and/or height8,29,60. Finally, the total biomass of the tree components per plot was computed based on the number of trees per plot.
Additionally, the ages of selected trees were measured by counting tree rings using the TSAP-Win computer program (F. Rinn Engineering Office and Distribution, Heidelberg). Stand age of planted forests (even-aged) was determined by the time since the stand was first created, while the ages of primary and secondary forests (uneven-aged) were determined by the mean age of all tree species29.
Site-related information, including site conditions (longitude, latitude, altitude), forest characteristics (stand age, stand density, forest type and forest origin [primary, secondary and planted forest]), soil chemistry (pH, total nitrogen [TN] and total phosphorus [TP]) and climate factors (e.g., mean annual temperature [MAT], mean annual precipitation [MAP]) were also documented in the dataset (see Appendix Data S1 for details).
Climatic variables, soil data and forest types
Growing season temperature and precipitation are important factors that affect large-scale patterns of biomass allocation18. Here, we used MAT (°C), MAP (mm), growing season length (GSL, days), maximum growing season temperature (MAXT, °C), mean growing season temperature (MT, °C), potential evapotranspiration (PET, mm) and aridity index (AI, PET/MAP), as indicators of climate variation. MAT, MAP and PET were extracted from a global climate dataset (0.0083° × 0.0083° resolution, approximately 1 km × 1 km) obtained from http://www.worldclim.org/. Growing season length (GSL, the number of days with diurnal mean temperature >5 °C), MAXT and MT were estimated from records (1970–1999) from 740 climate stations in China using a Kriging interpolation method31. Data were used in the analyses for each site at which the seven climatic variables and site conditions (latitude, longitude and altitude) were recorded. For records that lacked detailed altitudinal data, we used topographic maps to acquire this information.
Soil pH, TN and TP data were obtained from the second national soil survey and our field measurements. Forests in the dataset were primarily classified into eleven types: boreal/temperate Larix forest (BTLF), boreal/alpine Picea–Abies forest (BAPF), temperate Pinus tabulaeformis forest (TPTF), temperate/subtropical montane Populus–Betula deciduous forest (TSPF), subtropical montane Pinus yunnanensis and Pinus khasya forest (SPPF), subtropical Pinus massoniana forest (SPMF), subtropical montane Pinus armandii, Pinus taiwanensis and Pinus densada forest (SMPF), subtropical Cunninghamia lanceolata forest (SCLF), subtropical evergreen broadleaved forest (SEBF), tropical rainforest and monsoon forest (TRMF) and desert riverside woodland (DRW).
Statistical analysis
All data analyses were conducted with SPSS 19.0 (SPSS Inc., Chicago, IL). We used analysis of variance (ANOVA) to compare leaf, stem and root biomass of different forest types and origins to determine the effects of forest characteristics on patterns of biomass allocation. Pearson correlation analyses were performed to determine the relationships between leaf, stem and root biomass and site conditions (longitude, latitude and altitude). Principal component analysis (PCA) was completed to determine trade-offs in leaf, stem and root biomass. The PC axis with equal loading for leaf, stem and root biomass accounted for stand biomass increases and the axis with unequal loading encompassed the trade-offs in biomass between leaf, stem and root biomass21. In other words, the PC axis with equal loading was first used to constrain the influence of stand biomass on the forest biomass allocation and then the axis with unequal loading was used to determine trade-offs in leaf, stem and root biomass after stand biomass was constrained.
Stepwise multiple regressions (SMR) were used to identify the effects of forest stand characteristics (e.g., tree size, stand age, stand density) and environmental factors (e.g., climate and soil chemistry) on forest biomass and PC axes. In addition, general liner model (GLM) regressions were used to separate the variance explained by multiple factors into independent effects of individual factors and their interactive effects with the remaining factors34,35,36.
Additional Information
How to cite this article: Zhang, H. et al. Biogeographical patterns of biomass allocation in leaves, stems and roots in China's forests. Sci. Rep. 5, 15997; doi: 10.1038/srep15997 (2015).
References
Poorter, H. & Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Aust. J. Plant Physiol. 27, 595–607 (2000).
Poorter, H. et al. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytol. 193, 30–50 (2012)
Niklas, K. J. Plant allometry: The scaling of form and process (University of Chicago Press). Ch. 1, 5–12 (1994).
Reich, P. Root–shoot relations: optimality in acclimation and adaptation or the “Emperor’s new clothes”? in Plant roots: the hidden half, (eds Waisel Y., Eshel A., Kafkafi U. ) Ch. 7, 205–220 (2002).
Enquist, B. J. & Niklas, K. J. Global allocation rules for patterns of biomass partitioning in seed plants. Science 295, 1517–1520 (2002).
Reich, P. B. et al. Temperature drives global patterns in forest biomass distribution in leaves, stems and roots. Proc. Natl. Acad. Sci. USA 111, 13721–13727 (2014).
Cheng, D. & Niklas, K. Above- and below-ground biomass relationships across 1543 forested communities. Ann. Bot-London 99, 95–102 (2007).
Hui, D., Wang, J., Le, X., Shen, W. & Ren, H. Influences of biotic and abiotic factors on the relationship between tree productivity and biomass in China. For. Ecol. Manag. 264, 72–80 (2012).
Hui, D. et al. Near isometric biomass partitioning in forest ecosystems of China. PLoS ONE 9, e86550. 10.1371/journal.pone.0086550 (2014).
Luo, Y., Wang, X., Zhang, X., Booth, T. H. & Lu, F. Root:shoot ratios across China’s forests: Forest type and climatic effects. For. Ecol. Manag. 269, 19–25 (2012)
Luo, Y., Zhang, X., Wang, X. & Ren, Y. Dissecting variation in biomass conversion factors across China’s forests: implications for biomass and carbon accounting. PLoS ONE 9, e94777. 10.1371/journal.pone.0094777 (2014).
Kozlowski, J. & Konarzewski, M. Is West, Brown and Enquist’s model of allometric scaling mathematically correct and biologically relevant? Funct. Ecol. 18, 283–289 (2004).
Li, H. T., Han, X. G. & Wu, J. G. Lack of evidence of 3/4 scaling of metabolism in terrestrial plants. J. Integr. Plant Biol. 47, 1173–1183 (2005).
Li, H. T., Han, X. G. & Wu, J. G. Variant scaling relationship for mass-density across tree-dominated communities. J. Integr. Plant Biol. 48, 268–277 (2006).
Zhang, H., Wang, G. X., Liu, Z. Q., Shen, Z. X. & Zhao, X. Z. Sensitivity of response to abscisic acid affects the power of self-thinning in Arabidopsis thaliana. Bot. Bull. Acad. Sinica 46, 347–353 (2005).
Zhang, H., Wang, G. X., Shen, Z. X., Zhao, X. Z. & Qiu, M. Q. Effect of sensitivity to abscisic acid on scaling relationships for biomass production rates and body size in Arabidopsis thaliana. Acta Physiol. Plantarum 28, 373–379 (2006).
Zhang, H. et al. Plants interactions between Suaeda salsa individuals are mediated by salinity stress. Acta Physiol. Plantarum 30, 99–104 (2008).
Yang, Y. H., Fang, J. Y., Ma, W. H., Guo, D. L. & Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Global Ecol. Biogeogr. 19, 268–277 (2010).
Thornley, J. H. M. A balanced quantitative model for root: shoot ratios in vegetative plants. Ann. Bot-London 36, 431–441 (1972).
Bloom, A. J., Chapin, F. S. & Mooney, H. A. Resource limitations in plants – an economic analogy. Annu. Rev. Ecol. Syst. 16, 363–392 (1985).
Mccarthy, M. C. & Enquist, B. J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 21, 713–720 (2007).
Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities (Princeton University Press, Princeton, NJ, USA), Ch. 1, 12–18 (1988).
Bernacchi, C. J., Coleman, J. S., Bazzaz, F. A. & McConnaughay, K. D. M. Biomass allocation in old-field annual species grown γin elevated CO2 environments: no evidence for optimal partitioning. Glob. Change Biol. 6, 855–863 (2000).
Shipley, B. & Meziane, D. The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Funct. Ecol. 16, 326–331 (2002).
Niklas, K. J. & Enquist, B. J. Canonical rules for plant organ biomass partitioning and annual allocation. Am. J. Bot. 89, 812–819 (2002).
Zhao, M. & Zhou, G. S. Carbon storage of forest vegetation in China and its relationship with climate factors. Climatic Change 74, 175–189 (2006).
Fang, J. Y., Chen, A. P., Peng, C. H., Zhao, S. Q. & Ci, L. J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 292, 2320–2322 (2001).
Piao, S., Fang, J., Zhu, B. & Tan, K. Forest biomass carbon stocks in China over the past 2 decades: estimation based on integrated inventory and satellite data. J. Geophys. Res. 110, G01006 (2005).
Zhang, H. et al. Biomass and carbon storage in an age-sequence of Cyclobalanopsis glauca plantations in southwest China. Ecol. Eng. 73, 184–191 (2014).
Gill, R. A. & Jackson, R. B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 147, 13–31 (2000).
Zerihun, A., Montagu, K., Hoffmann, M. B. & Bray, S. G. Patterns of below- and aboveground biomass in Eucalyptus populnea woodland communities of northeast Australia along a rainfall gradient. Ecosystem 9, 501–515 (2006).
Cairns, M., Brown, S., Helmer, E. & Baumgardner, G. Root biomass allocation in the world’s upland forests. Oecologia 111, 1–11 (1997).
Piao, S. L. et al. The carbon balance of terrestrial ecosystems in China. Nature 458, 1009–1013 (2009).
Heikkinen, R. K., Luoto, M., Kuussaari, M. & Poyry, J. New insights into butterfly-environment relationships using partitioning methods. Proc. Royal Soc. (B), 272, 2203–2210 (2005).
Han, W. X., Fang, J. Y., Reich, P. B., Woodward, F. I. & Wang, Z. H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. lett. 14, 788–796 (2011).
Hedin, L. O. Global organization of terrestrial plant-nutrient interactions. Proc. Natl. Acad. Sci. USA 101, 10849–10850. (2004).
Wright, I. J. et al. The worldwide leaf economics spectrum. Nature 428, 821–827 (2004).
Guo Q. F. & Ren H. Productivity as related to diversity and age in planted versus natural forests. Global Ecol. Biogeogr. 23, 1461–1471 (2014).
Evans, G. C. The Quantitative Analysis of Plant Growth (Oxford, UK: Blackwell Scientific Publications), Ch. 1, 3–12 (1972).
Poorter, H. & Pothmann, P. Growth and carbon economy of a fast-growing and a slow-growing species as dependent on ontogeny. New Phytol. 120, 159–166 (1992).
Coleman, J. S., McConnaughay, K. D. M. & Ackerly, D. D. Interpreting phenotypic variation in plants. Trends Ecol. Evol. 9, 187–191 (1994).
Wang, X., Fang, J. & Zhu, B. Forest biomass and root-shoot allocation in northeast China. For. Ecol. Manag. 255, 4007–4020 (2008).
Mokany, K., Raison, R. J. & Prokushkin A. S. Critical analysis of root: shoot ratios in terrestrial biomes. Global Change Biol. 12, 84–96 (2006).
Zhang, W. P., Jia, X., Morris, E. C., Bai, Y. Y. & Wang, G. X. Stem, branch and leaf biomass-density relationships in forest communities. Ecol. Res. 27, 819–825 (2012).
Santiago, L. S. et al. Leaf photosynthetic traits scale with hydraulic conductivity. Oecologia 140, 543–550 (2004).
Finér, L., Ohashi, M., Noguchi, K. & Hirano, Y. Factors causing variation in fine root biomass in forest ecosystems. For. Ecol. Manag. 261, 265–277 (2011).
Kirschbaum, M. U. F. The temperature dependence of soil organic matter decomposition and the effect of global warming on soil organic C storage. Soil Biol. Biochem. 27, 753–760 (1995).
Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems (Springer, Berlin), Ch. 1, 1–13 (1999).
Aerts, R. & Chapin, F. S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 30, 1–67 (2000).
Hobbie, S. E., Schimel, J. P., Trumbore, S. E. & Randerson, J. R. Controls over carbon storage and turnover in high-latitude soils. Glob. Change Biol. 6, 196–210 (2000).
Rubilar, R. A. et al. Influences of silvicultural manipulations on above- and belowground biomass accumulations and leaf area in young Pinus radiata plantations, at three contrasting sites in Chile. Forestry 86, 27–38 (2013).
Cunningham, S. A., Summerhayes, B. & Westoby, M. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecol. Monogr. 69, 569–588 (1999).
Poorter, H. & Sack, L. Pitfalls and possibilities in the analysis of biomass allocation patterns in plants. Front. Plant Sci. 3, 259 (2012).
Padilla, F. M., Miranda, J. D., Jorquera, M. J. & Pugnaire, F. I. Variability in amount and frequency of water supply affects roots but not growth of arid shrubs. Plant Ecol. 204, 261–270 (2009).
Poorter, H., Niinemets, Ü., Poorter, L., Wright, I. J. & Villar, R. Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol. 182, 565–588 (2009).
Deng, J. M. et al. Plant mass–density relationship along a moisture gradient in north-west China. J. Ecol. 94, 953–958 (2006).
Sitch, S. et al. Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models (DGVMs). Glob. Change Biol. 14, 2015–2039 (2008).
Piao, S. L. et al. Interannual variations of monthly and seasonal normalized difference vegetation index (NDVI) in China from 1982 to 1999. J. Geophys. Res. 108, 10.1029/2002JD002848 (2003).
Wolf, A., Field, C. B. & Berry, J. A. Allometric growth and allocation in forests: A perspective from FLUXNET. Ecol. Appl. 21, 1546–1556 (2011).
Wang, Z. C., Du, H., Song, T. Q., Peng, W. X., Zeng, F. P., Zeng, Z. X. & Zhang, H. Allometric models of major tree species and forest biomass in Guangxi. Acta Ecol. Sin. 35, 4462–4472 (2015) (In Chinese).
Acknowledgements
This research was supported by the Chinese Academy Sciences Action Plan for the Development of Western China (KZCX2-XB3-10); the Major State Basic Research Development Program of China (2015CB452703); the Strategic Priority Research Program-Climate Change: Carbon Budget and Related Issues of the Chinese Academy of Sciences (XDA05070404 and XDA05050205), the National Natural Science Foundation of China (Nos. 31370485, 31370623, 31400412 and 41471233); the “100 talents program” of the Chinese Academy of Sciences (2060299, Y251101111); the Western Light Program of Talent Cultivation of the Chinese Academy of Sciences, State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources and Guangxi Provincial Program of Distinguished Expert in China.
Author information
Authors and Affiliations
Contributions
H.Z. and F.Z. designed the study. K.W. and X.X. performed the data collection and analysis. H.Z., F.Z., K.W., T.S., X.X. and Y.X. interpreted the results and wrote the paper. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhang, H., Wang, K., Xu, X. et al. Biogeographical patterns of biomass allocation in leaves, stems and roots in China’s forests. Sci Rep 5, 15997 (2015). https://doi.org/10.1038/srep15997
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep15997
This article is cited by
-
Symbiotic mycorrhizal types affect patterns of tree aboveground and belowground C allocation in Northeast China
Ecological Processes (2023)
-
Does Long-Term Industrial Pollution Affect the Fine and Coarse Root Mass in Forests? Preliminary Investigation of Two Copper Smelter Contaminated Areas
Water, Air, & Soil Pollution (2022)
-
The global distribution and environmental drivers of aboveground versus belowground plant biomass
Nature Ecology & Evolution (2021)
-
Distributional patterns of soil nematodes in relation to environmental variables in forest ecosystems
Soil Ecology Letters (2021)
-
Can Dominant Canopy Species Leaf Litter Determine Soil Nutrient Heterogeneity? A Case Study in a Tropical Rainforest in Southwest China
Journal of Soil Science and Plant Nutrition (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
