
Abstract
The Turin Shroud is traditionally considered to be the burial cloth in which the body of Jesus Christ was wrapped after his death approximately 2000 years ago. Here, we report the main findings from the analysis of genomic DNA extracted from dust particles vacuumed from parts of the body image and the lateral edge used for radiocarbon dating. Several plant taxa native to the Mediterranean area were identified as well as species with a primary center of origin in Asia, the Middle East or the Americas but introduced in a historical interval later than the Medieval period. Regarding human mitogenome lineages, our analyses detected sequences from multiple subjects of different ethnic origins, which clustered into a number of Western Eurasian haplogroups, including some known to be typical of Western Europe, the Near East, the Arabian Peninsula and the Indian sub-continent. Such diversity does not exclude a Medieval origin in Europe but it would be also compatible with the historic path followed by the Turin Shroud during its presumed journey from the Near East. Furthermore, the results raise the possibility of an Indian manufacture of the linen cloth.
Similar content being viewed by others

Introduction
The Turin Shroud (TS) is a linen cloth, 4.4 m long and 1.1 m wide, bearing the double image of a man who suffered physical trauma in a manner consistent with crucifixion after being beaten, scourged and crowned with thorns1,2. TS is the most important relic of Christianity because the Catholic tradition identifies this burial cloth as that in which the body of Jesus Christ was wrapped before being placed in a Palestine tomb approximately 2000 years ago. Such a scenario is supported by numerous scholars who believe that the journey of TS began in Jerusalem in the year 30 or 33 AD3. After concealment for years, TS would have been first moved to Edessa (now Şanliurfa in Turkey) and then to Constantinople (now Istanbul in Turkey) in 944 AD. A burial cloth, which some historians consider the Shroud, was owned by the Byzantine emperors but disappeared during the Sack of Constantinople in 12044. After this event, TS would have been taken by the crusaders and transferred to Athens (Greece), where it remained until 1225. Official documents attest that it was in France at Lirey around the years 1353 to 1357 and then was kept at Chambéry from 1502 to 1578, where passed into the hands of the Dukes of Savoy3,4,5. From 1578, apart from some brief displacements in an effort to hide it during war periods, TS was kept in Turin (Italy) and later placed in the royal chapel of the city Cathedral inside a specially designed shrine where it has been permanently conserved from 1694 to the present.
The TS shows many marks caused by human blood, fire, water and folding of the cloth that partially obscure the double, front and back, body image that is not yet reproducible1,2,6,7. In 1988, the age of the TS linen cloth was assessed by accelerator mass spectrometry. Results of radiocarbon measurements from distinct and independent laboratories yielded a calendar age range of 1260–1390 AD, with 95% confidence8, thus providing robust evidence for a Medieval recent origin of TS. However, two papers have highlighted some concerns about this determination9,10 and a Medieval age does not appear to be compatible with the production technology of the linen nor with the chemistry of fibers obtained directly from the main part of the cloth in 19781,11.
In 1978 and 1988, dust particles were vacuumed from the interspace between the Shroud and the Holland Cloth sewn to it as reinforcement12. The composition of the particles was later studied in great detail by optical microscopy and specimens from different filters were retained and characterized for their contents6,13. In past decades, pollen grains were classified to the genus and species levels using microscopy14,15,16 and the geographic areas where the corresponding plants originated and now inhabit proved to be compatible with the reported historic path followed by TS during the postulated 2000-year journey from the Near East3, thus supporting the authenticity of the relic.
In this study, we performed DNA analyses to define the biological sources of the dust particles (pollen grains, cell debris and other minuscule organic specimens, such as plant-derived fibers and blood-like clots) vacuum-collected in 1978 and 1988 in distinct TS filters, corresponding to the face, hands, glutei and feet of the body image6,13 and the lateral edge, which was used for radiocarbon dating8. To identify plant taxonomic entities and human genetic lineages, universal plant DNA sequences, including nuclear rDNA intergenic transcribed spacers (ITS) and chloroplast DNA (cpDNA) barcodes and human mitochondrial DNA (mtDNA) target regions were amplified and sequenced. This allowed the identification of DNA sources from a wide range of plant species and human mitogenomes belonging to numerous haplogroups. The overall findings were then evaluated to determine whether the geographic areas of origin and distribution of detected plant cpDNA species and human mtDNA haplogroups might provide novel clues concerning the origin of the Turin Shroud.
Results
Detection of plant DNA from the Turin Shroud and identification of plant species
More than 100 PCR-derived amplicons were recovered from genic and intergenic target regions, 77 of which were successfully sequenced and attributed to a genus or species source (19 different plant taxa). Approximately half of the DNA sequences derived from the TS lateral edge samples (filter I) and allowed us to identify 16 plant species. In filters E-H, corresponding to various parts of the TS male body image, we assigned DNA sequences to one (glutei), two (feet), or three plant species (face and hands). Table 1 summarizes the plant species found on TS by querying databases of orthologous sequences from taxa of established identity. The land plant species include herbaceous weeds and crops, woody trees and shrubs; some are native to Mediterranean countries and are widespread in Central Europe, North Africa and the Middle East, whereas others have a center of origin in Eastern Asia and the Americas and hence they were not yet present into Europe during the Medieval period (Supplementary Table S1).
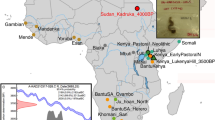
Among the taxa identified, the most abundant belongs to the genus Picea, which includes a few closely related species native to Europe (P. abies (L.) H. Karst., P. obovata Ledeb. and P. omorika Purk.) and several spruce trees widespread in temperate and boreal forest regions of the Northern Hemisphere. However, other species are present, including those native to the Mediterranean basin, such as clovers (Trifolium spp.), ryegrasses (Lolium spp.) and plantains (Plantago spp.) and Eastern Asia, such as uncommon forms of pear (Pyrus kansuensis Rehder) and plum (Pyrus spinosa Forssk, syn. Pyrus amygdaliformis Vill.) of the family Rosaceae. Among the plant species of the New World, black locust (Robinia pseudoacacia L.), a tree of the family Fabaceae native to Appalachia in the Eastern United States, is notable. In addition, we identified crop species largely grown by farmers and common in many agriculture systems of the Old World, including chicory (Cichorium intybus L.), common hop (Humulus lupulus L.), cucumber (Cucumis sativus L.) and grapevine (Vitis or Parthenocissus spp.). We also uncovered tree species commonly present in forests and woodlands, such as hornbeams (Carpinus spp.), walnuts (Juglans spp.) and willows (Salix spp.); although the centers of origin of these species are located in central Asia and Eastern Europe, their current areas of distribution are extremely broad (Fig. 1).

Plant DNA species found on the Turin Shroud.
Schematic overview of the Vavilov centers of origin of plant taxa identified in TS samples. The number of amplicons is reported for each species and across filters E, F, G, H and I (see also Table 1 for details on the distribution of species among TS filters). The world map used as background to create this schematic overview has been obtained from Wikimedia Commons, the free media repository (https://commons.wikimedia.org/wiki/seefileNo_colonies_blank_world_map.png).
Based on the overall data, we may assume that TS was likely displayed in, or in very close contact with, different types of natural and anthropological environments. The large variety of taxonomic entities is compatible with highly diverse geographic ranges, varying from farms of cultivated plains to riparian woodlands and mountain forests. Some species have a center of origin and have diversified in areas around the Mediterranean basin, including North Africa and the Middle East and most of these species were widely distributed throughout Europe before the age of Christ. However, other species identified on TS were not introduced to Europe before the XVI century, after the discovery of America by Christopher Columbus (for instance, Robinia pseudoacacia and also nightshades of the family Solanaceae), while the two species of the genera Prunus and Pyrus, rare fruit trees originating from South-East Asia and the Middle East, were likely introduced to Mediterranean territories from the XIII century and thereafter, following the travels of Marco Polo (Supplementary Table S1).
Examination of human mtDNA haplotypes from the Turin Shroud and identification of mtDNA haplogroups
Among the 93 mtDNA amplicons generated and sequenced, a large number of different human sequences corresponding to three distinct mtDNA loci (MT-DLOOP, MT-CO1, MT-ND5) were identified. This result not only indicates that human DNA was indeed unequivocally present in the dust from TS but also that the sources of human DNA could be ascribed to numerous individuals (Table 2). In fact, the mtDNA haplotypes were found to belong to different branches of the human mtDNA tree (Supplementary Table S2), even after having excluded all the mtDNA sequences that could be theoretically attributed to operator contamination (Supplementary Table S3). Moreover, not only were the observed mtDNA haplotypes numerous, but they could also be affiliated to many distinct haplogroups. Six sub-branches of haplogroup H (H1, H2, H3, H4, H13, H33) are included, as well as representatives of haplogroups U2, U5, R0a, R7, R8, L3c, M39 and M56 (Table 2).
The number and proportion of read clusters derived from the different TS samples for each of the identified haplogroups are shown in Fig. 2, together with a schematic overview of their current geographic distributions. Some haplogroups are widespread, while others are geographically and ethnically more localized (see Supplementary Table S2 for detailed information). For instance, haplogroup H1 is very common in Western Europe, with a frequency peak among Iberians (~25%) but also among the populations of Northwestern Africa, including the Berbers. Haplogroup H4 is instead present at low and rather similar frequencies in Western (Iberia ~3%) and Eastern Europe (~1%), the Caucasus (~3%) and the Near East (~1%). Haplogroup H33 is rare and mainly found thus far among the Druze, a minority population of Israel, Jordan, Lebanon and Syria. Haplogroup U2 is found largely in South Asia (~5%), but one of its subsets (U2e) is present in Europe, with a frequency of ~1%. Haplogroup U5 harbors an average frequency of 7% in modern European populations and its major sub-branches, U5a and U5b, are most common in Eastern and Western Europe, respectively. Haplogroup R0a is predominantly localized in the Arabian Peninsula and the Horn of Africa, with the highest frequency in southeast Yemen (approximately 30%), though it is also found at low frequencies all over Western Eurasia. Haplogroup L3c is extremely rare and only found in East Africa, while haplogroups M39, M56, R7 and R8 are typical of the Indian subcontinent, with the latter essentially present only in Eastern India.

Human mtDNA haplogroups found on the Turin Shroud.
Schematic overview of the current geographical distribution of human mtDNA haplogroups and sub-haplogroups identified in TS samples. The number and proportion of reads derived from samples EFGH, I and IR are shown on the left for each haplogroup. The world map used as background to create this schematic overview has been obtained from Wikimedia Commons, the free media repository (https://commons.wikimedia.org/wiki/seefileNo_colonies_blank_world_map.png).
In brief, mtDNA data indicate that numerous individuals have left traces of DNA on the TS. Moreover, their mtDNA sequences belong to haplogroups that are typical of different ethnic groups and/or different geographic regions, including not only Europe where official documents verify the presence of TS since 1353 AD but also North and East Africa, the Middle East and even India.
Collection of non-plant and non-human DNA sequences
It is worth mentioning that among all generated and sequenced cpDNA and mtDNA amplicons (~200 overall), only a few non-plant and non-human sequences were detected (Supplementary Table S1). One of these sequences, although very short being 58 bp long, partially matched and produced the best alignment with the MT-CYB gene (accession no. AY827092.1) from the southern grey shrike (Lanius meridionalis Koenigi), a medium-sized passerine bird that is reported to be resident in Southern Europe, Northern Africa and the Near East. Another of these sequences, corresponding to 694 bp of the CO1 gene, was ascribable to a marine worm (Cerebratulus longiceps Coe), rather common in the Northern Pacific Ocean, next to Canada (accession no. JQ007428-JQ007431).
Discussion
DNA extracted from dust particles that were vacuumed from the Turin Shroud shows sequence profiles that identify numerous plant species and correspond to several distinct human mtDNA haplogroups. These results not only confirm that plant fibers and pollen grains are present on TS, as previously reported by optical microscopy, but also reveal that multiple human individuals touched or otherwise left traces of their DNA on the relic linen. The detection of such a variety of DNA sources is extremely valuable in assessing whether there are possible parallelisms between the areas of origin and distribution of identified land plant species and human mtDNA haplogroups and the temporal and spatial paths associated with the two alternative scenarios that have been proposed to explain the TS origin.
The radiocarbon measurements would place the origin of the TS linen in the time frame 1260–1390 AD. This not only implies a Late Middle Age origin, but a geographical path for the TS that is essentially restricted to Western Europe. In this scenario, the DNA traces that we detected could have entered in contact with the TS only rather recently, at most in the last 800 years and these biological sources (plants and human subjects) had to be present in the geographic areas (France and Italy) where the TS was located and/or displayed. The alternative scenario implies instead a much longer journey that started in Jerusalem in the year 30 or 33 AD. In this case, the time frame for the interaction with the DNA biological sources is much longer (2000 years) and the geographic areas where the TS was located include the Near East, Anatolia, Eastern and Western Europe, with a potentially much wider range of plant and human interactions.
With regard to the land plant species identified, some are native to Mediterranean countries and widespread throughout Europe, North Africa and the Middle East and are thus compatible with both a rather recent Medieval origin in Europe and a more ancient Near Eastern origin. However, others have a center of origin in Eastern Asia and the Americas and were introduced to Europe only after the Medieval period. Clearly, the latter species cannot help in discriminating between alternative scenarios.
The quantitatively most abundant species found on TS dust is spruce: the vast majority of DNA sequences assigned to the genus Picea are likely attributable to the species Picea abies (L.) H. Karst., a forest tree that typically occupies highland areas of the Carpathians and Alps. Among the NCBI sequences most similar to those obtained from TS, one belongs to a spruce tree sampled in the Swiss Alps (accession number AF327585); this discovery is in accord with the transport of TS through the French-Italian Alps in 1578 when the relic was moved from Chambéry to Turin. Of note, our trnL-intron sequences shared the haplotype with most of those of P. abies accessions deposited in public databases, supported by specimen vouchers and annotated with a Southern European origin (e.g., Serbia) and an Eastern Asian origin (e.g., China), revealing not only common SNPs but also many private polymorphisms (accession numbers JQ007384-JQ007406 and Supplementary Figure S1).
Most of the plant species identified based on cpDNA and ITS sequences (e.g., clovers, ryegrasses, plantains and chicories) have an origin and are now widespread in regions around the Mediterranean basin, from the Iberian Peninsula to Palestine. The presence of some alien tree species introduced from Eastern USA (Robinia pseudoacacia L.) and Northern or Southern China (Salix suchowensis W.C. Cheng, Pyrus spinosa Forssk and Prunus kansuensis Rehder) is not negligible, with the former species that currently has a distribution area centered in Europe and the Mediterranean basin and the latter that are more widespread in temperate Asia, Southern Europe and Northern Africa. Overall, the various plant species and numerous taxonomic families identified on TS (Supplementary Table S1) suggest that contamination may have occurred during the past centuries and are compatible with the scenario that the linen cloth was exposed to different locations across the Mediterranean area.
With regard to the sources of human DNA, the detected haplotypes do not cluster randomly on the entire human mtDNA tree, but only on a specific subset of its branches, corresponding to numerous haplogroups from Western Eurasia and surrounding areas (Supplementary Table S2). This finding not only indicates that many individuals have left traces of their DNA on the TS, but also that they most likely belonged to different ethnic groups and were from far away geographic regions, including Europe, North and East Africa, the Middle East and India. Thus, the sources of these sequences fit well the geographic path of the postulated long journey from the Near East, even if they are also fully compatible with the scenario that among the perhaps thousands of worshippers who came into contact with the relic in France and Italy throughout the centuries, there were many coming from the far away geographic areas where these mtDNA haplogroups are common. Moreover, it should be taken into account that filters E, F, G and H correspond to the internal parts of the linen cloth that enveloped the body of the TS man; in contrast, filter I derives from the lateral edge of the cloth. Because the cloth was folded, the lateral edges were not only in contact with the external environment much more than the internal and more protected body image but are also the TS regions that were handled the most. Thus, filters E, F, G and H might provide more reliable clues than filter I with regard to the TS earliest contacts and contamination and, therefore, its hypothesized historical path. There are six haplogroups (H4, L3c, M39, R7, U2 and U5) that were detected only in the dust particles from filter I. If these are excluded, four partially overlapping geographic regions are generally outlined by the remaining haplogroups: (i) the Middle East with haplogroups H13, H33 and R0a; (ii) Southeast Europe and Turkey with haplogroups H1a, H2a and H13; (iii) Western Europe, including France and Italy, with haplogroups H1j and H3; and (iv) India with haplogroups M56 and R8.
In conclusion, our results on human mtDNA traces detected on the TS are compatible with both alternative scenarios that i) the cloth had a Medieval origin in Western Europe where people from different geographic regions and ethnic affiliations came in contact with it, possibly moved by the worship for the Christian relic; ii) the linen cloth had a Middle Eastern origin and was moved itself across the Mediterranean area, consequently coming across a wide range of local folks and devotes in a longer time span. Even in the latter case (i.e., Jerusalem in Israel until approximately 500 A.D., Şanliurfa in Turkey until 944, Constantinople in Turkey until 1204, Lirey and Chambery in France from 1353 until 1578, Turin in Italy to date)3, the detection of mtDNA haplogroups that are typically from India is somehow unexpected. One obvious possibility is that during the course of centuries, individuals of Indian ancestry came into contact with TS. Taking into account the rate of DNA degradation and PCR-biases toward undamaged DNA, the recent contamination scenario is extremely likely. However, one alternative and intriguing possibility is that the linen cloth was weaved in India, as supported perhaps by the original name of TS - Sindon - which appears to derive from Sindia or Sindien, a fabric coming from India.
Methods
Collection of TS Samples for DNA analyses
The samples used in this study for DNA investigations are sub-sets of the dust particles vacuumed from the back of the Turin Shroud (TS), which were kindly provided in 2010 by Giulio Fanti (Dept. of Industrial Engineering, University of Padua, Italy), who in turn received them personally from Giovanni Riggi di Numana in 200612,18 (see also Supplementary Document S1). In particular, four samples were originally collected on filters in 1978 from four distinct areas of the back of TS, corresponding to the hands (filter E), face (filter F), feet (filter G) and glutei (filter H) of the TS male image12. An additional sample was collected in 1988, deriving specifically from the corner area (filter I) of TS, which was used for radiocarbon dating8,12 (for details, see Supplementary Figure S2, panels A–C).
The dust samples collected on these filters were immobilized on adhesive tapes as previously described6,13 and consisted of a variety of particles. In fact, optical microscope observations of filters E-I had previously revealed the presence of pollen grains, cell debris and other minuscule organic specimens, such as plant-derived fibers and blood-like clots6,7,13.
The five pieces of adhesive tape containing TS dust particles collected on filters E, F, G, H and I were supplied as portions of the original samples (ranging in size from 5 × 10 mm to 10 × 30 mm, see the schematic representation in Supplementary Figure S2, panel D), each attached to a sterile microscope slide. Each piece of adhesive tape was manually cut using a scalpel and tweezers under a stereomicroscope into tiny sub-portions of dimensions on the order of a few square millimeters (~5 × 5 mm each); the pieces were then individually transferred into 1.5-ml sterile microtubes. A total of 2 to 12 square portioned specimens of the pieces of adhesive tape were used for independent genomic DNA extractions; the DNA samples from the individual specimens were kept separate for PCR amplifications.
PCR amplicons obtained from each of the cpDNA regions were sub-cloned and used individually for DNA sequencing, whereas PCR amplicons obtained from each of the mtDNA regions were pooled for 454 sequencing reactions. In particular, the DNA amplicons from each of the 2, 4 or 6 specimens associated with internal filters E, F, G and H were pooled together into single samples, whereas the DNA amplicons from external filter I were divided into two samples of 6 specimens each (named I and IR and taken as independent biological replications; Supplementary Figure S2). This pooling strategy was imposed by the different sizes of the pieces of adhesive tape to be analyzed for each of the five TS filters and by the fact that dust particles were demonstrated to be much more abundant on the tape representing the external filter than on those for the internal filters6,13.
All manipulation steps were performed aseptically under laminar flow hood conditions using autoclaved disposables and buffers that were filter-sterilized. In particular, we used sterile materials and all standard procedures with internal negative controls to avoid, or eventually discover, contaminations from the operators and the environment. The entire mitochondrial genomes of the three operators who came in contact with the Shroud samples were completely sequenced and all mtDNA sequences that we obtained from the TS samples and could be theoretically attributed to operator contamination (Supplementary Table S3) were excluded from our final results. As for possible environmental contaminations, none of the large varieties of plant sources detected in the Shroud (Supplementary Table S1) through the analysis of nuclear ITS, chloroplast genes or cpDNA sequences are grown or studied in our facilities.
Genomic DNA extraction
Specimens from the pieces of adhesive tape with the TS dust particles were used individually for genomic DNA extraction to avoid any loss of TS material and to perform replicate experiments for each TS filter.
All genomic DNA extractions were performed with the QIAamp®DNA Investigator commercial kit (Qiagen) by applying the protocol originally developed for laser-microdissected specimens, with some modifications. All manipulation steps were performed under laminar flow hood conditions using DNA- and DNase-free, disposable autoclaved materials and filter-sterilized buffers. Immediately after collection, sub-samples were transferred to a 1.5-ml tube containing a pre-warmed mixture composed of 40 μl of ATL buffer and 20 μl proteinase K. Each sample was then mixed by pulse-vortexing and incubated at 56 °C for 16 hours under constant agitation. After this step, 100 μl of the AL buffer, containing 2 μg of carrier RNA, was added and the solution was mixed by pulse-vortexing for 15 sec. Then, 100 μl absolute ethanol was added and the solution was mixed thoroughly by pulse-vortexing for 15 sec and then incubated for 5 min at room temperature. The next steps, which consisted of DNA immobilization to a silica membrane and wash steps to clean the isolated DNA from salts and impurities, were conducted by following the manufacturer’s suggestions. Elution of the DNA immobilized by the silica membrane was performed in two steps by using 15 μl ATE buffer at each elution step and by extending the incubation step to 10 min at room temperature.
To identify human genetic lineages and plant taxonomic entities, specific plant DNA and human mitochondrial DNA regions were designated for PCR amplification and sequencing.
PCR amplification and sequencing of plant and human DNA sequences
PCR amplifications of plant DNA sequences were performed with primer combinations designed using nuclear (rDNA intergenic transcribed spacers, ITS) and chloroplast targets (cpDNA barcodes RuBisCO or rbcL, trnH-psbA and trnL-intron) (Supplementary Table S4), following already available protocols19,20. Briefly, the reactions were performed in a total volume of 20 μl that included 2 μl of 10X reaction buffer, 1 mM MgSO4, 0.3 mM dNTPs, 0.25 U of Platinum® Pfx DNA Polymerase (Life Technologies), 0.3 μM of primer mix and 2.5 μl of eluted DNA solution. The reactions were performed in a 9700 Thermal Cycler (Applied Biosystems) using a temperature profile that consisted of an initial denaturation step of 10 min at 95 °C followed by 50 cycles of 1 min at 95 °C, 30 sec at 55 °C, 1 min at 68 °C and a final step of 10 min at 68 °C.
The same conditions were adopted for PCR amplifications of human sequences corresponding to three distinct mtDNA loci (MT-DLOOP with the hypervariable segments MT-HV1 and MT-HV2, MT-CO1 and MT-ND5) using specific primer pairs (Supplementary Table S4).
The amplicons were sub-cloned by ligation into the TOPO-blunt cloning vector (Life Technologies) and transformed into chemically competent one-shot TOP10 bacterial cells (Life Technologies). Clones were plated on LB plates (1.5% agar, 50 μg/mL ampicillin, 40 μg/ml X-Gal) and transformed colonies were selected by Colony-PCR. Amplification reactions were performed in a total volume of 20 μl including 2 μl of 10X reaction buffer, 1.5 mM MgCl2, 300 μM dNTPs, 1.5 U of BIOTaq DNA polymerase (BIOLINE), 0.2 μM of M13For (5′-GTAAAACGACGGCCAG-3′) and M13Rev (5′-CAGGAAACAGCTATGAC-3′) primers. Positive colonies were sequenced using an ABI3100 automated sequencer (Applied Biosystems).
Both nuclear ITS sequences and DNA barcodes were used to identify plant and fungal species21,22, querying the Barcode of Life Data BOLD Systems v. 3 (http://www.boldsystems.org/index.php/IDS_OpenIdEngine). Sequence similarity searches in nucleotide collections (nt) and with non-redundant protein sequences (nr) were also performed with blastn/x programs of the Basic Local Alignment Search Tool BLAST v. 2.2.30+ (http://blast.st-va.ncbi.nlm.nih.gov/Blast.cgi) using default parameters. All plant cpDNA and ITS sequences deriving from PCR amplicons were verified through replicated sequencing of both strands and deposited in GenBank with accession numbers JQ007354-JQ007431 and JQ082521-JQ082524. The human mtDNA sequences, as clusters deriving from the assembly of reads generated by 454 sequencing, were deposited in GenBank with accession numbers KP126143-KP126230.
For each plant sequence, we collected information regarding: i) the TS location source; ii) the GenBank accession number of the most similar sequence; iii) the Linnaean name of the species, including the botanical family and its common name; iv) the name and length of the target DNA sequence; v) the center of origin and geographical distribution of the species, along with information on plant type and/or common use (for details, see Supplementary Table S1). From an experimental point of view, it should be noted that some species were identified in biological replicates by using different specimens of the same filter as well as different filters but were also assessed by technical replicates according to amplicons from different genetic targets. In addition, other species were identified by performing independent experiments, meaning that the PCR analyses were conducted at different times using DNA samples isolated at different times from the same filters/dust samples. All these cases are pivotal experimental validations and demonstrate the robust assignments of plant species (or genus).
PCR amplification of human mtDNA and preparation of libraries for pyrosequencing
Amplifications of target mtDNA sequences prior to pyrosequencing were performed as described in the previous paragraph using specific primer sets (Supplementary Table S4). It is worth mentioning that the successful amplification of human mtDNA fragments ranging in size from 419 to 576 bp (as well as plant nuclear and chloroplast DNAs from 229 to 622 bp) such as the ones that we amplified, would be mostly possible from undamaged or slightly damaged DNA templates, which is unexpected when dealing with very ancient DNA specimens. However, the size of ancient DNA fragments, in addition to age, is influenced by many additional factors such as environmental conditions (e.g., temperature, moisture and pH) and mode of preservation (e.g., museum specimens vs. freshly excavated remains). In brief, DNA degradation becomes more effective only over long intervals, but is de facto unpredictable over shorter time spans such as those postulated for the DNA traces on the TS and when considering the numerous copies of extra-nuclear DNA molecules23,24,25,26.
PCR products originating from the amplification of multiple samples were pooled (Supplementary Table S4) and purified with QIAquick PCR Purification Kit (QIAGEN) by following the recommendations of the supplier. Next, the amplicons of the three different mtDNA target regions were pooled together, as reported in Supplementary Table S5 and purified with the Agencourt AMPure XP procedure (Beckman Coulter) by following the recommendations of the supplier. Quantification of the pooled amplicons was performed with a Nanodrop fluorometer ND 3300 (Thermo Scientific).
Libraries were prepared starting from 100 ng of purified PCR products using Rapid Library Preparation Method (Series GS FLX+; Roche 454 sequencing), starting from step 3.2: Fragment End Repair (thus avoiding the step 3.1, DNA Fragmentation by Nebulization). Single libraries were tagged with univocal MIDs (Supplementary Table S6). Emulsion PCR and 454 sequencing were carried out according to the manufacturer’s instructions on the Roche 454 FLX Titanium platform. Library sequencing was carried out using a Roche 454 GS Junior System.
Analysis of sequence reads, variant calling and cluster assembly
Sequence reads were de-multiplexed based on their MID sequence with the Roche 454 software SFFfile. The number of sequenced and aligned reads referring to the single libraries are reported in Supplementary Table S7.
Mapping of the sequence reads to the H. sapiens mitochondrial genome (gb|NC_012920.1) was performed with the software CLC Genomics Workbench version 5.05 with defaults parameters. Mapping of the sequence reads originating from each library was used to estimate the average coverage for each target region. Background noise in the sequences was filtered with a program developed ad hoc that analyzes the composition in words (k-mer) of the produced sequences and masks the words having a frequency lower than the value set as the cutoff. For this step, the length of the k-mers was set to 7 and the minimum frequency parameter was set to 5. After the masking of rare variants, the sequences were clustered with the software CD HIT v.4.5.427 by setting a threshold of percent identity equal to 99%. The average coverage of the mtDNA target regions for each single pool and target region is summarized in Supplementary Table S7.
Sequence clusters with a length greater than 200 bp were aligned to the H. sapiens mitochondrial genome (gb|NC_012920.1) using the program “bwa”28 with default parameters. In this step, each pool of sequences was independently aligned against the reference sequence17. The alignment files were saved as standard SAM files. Variant calling was performed on alignments of sequences representative of each cluster with the software PASS29. For each cluster, the haplotype was defined as the collection of substitutions (either transversions or transitions) that emerged in the variant calling process. Insertions and deletions were not taken into account.
A pre-filtering step for estimation of the mtDNA haplogroups represented in our cluster sequences was performed with the software HaploGrep30, which allows the automatic assignment of haplogroups according to Phylotree31. Preliminary assignments of haplogroups to sequence clusters displaying the highest sequence coverage were manually verified. The frequency of each haplogroup was estimated by the number of sequences contained in each cluster assigned to that specific haplogroup. The distribution of haplogroups in the different samples was estimated as above, but by counting the sequences according to the library information. The assembled human mtDNA sequences were deposited in GenBank with accession numbers KM655881-KM655934 (for details, see Supplementary Table S8).
Additional Information
How to cite this article: Barcaccia, G. et al. Uncovering the sources of DNA found on the Turin Shroud. Sci. Rep. 5, 14484; doi: 10.1038/srep14484 (2015).
Change history
22 March 2021
A Correction to this paper has been published: https://doi.org/10.1038/s41598-021-85973-1
References
Jumper, E. J. et al. A comprehensive examination of the various stains and images on the Shroud of Turin (In: Archaeological Chemistry III, ACS Advances in Chemistry 205, 22, American Chemical Society, Washington D.C., pp. 447–476, 1984).
Adler, A. The Orphaned Manuscript – a Gathering of Publications on the Shroud of Turin (A Shroud Spectrum Intl. Special Issue, Ed. Effatà Editrice, Turin, Italy, 2002).
Antonacci, M. The Resurrection of the Shroud (M. Evans & Co, New York, NY, 2000).
Poulle, E. Les sources de l’histoire du linceul de Turin. Revue critique. Revue d’Histoire Ecclésiastique 3–4, 747–781 (2009).
Meacham, W. The authentication of the Turin Shroud, an issue in archeological epistemology. Current Anthropology 24, 283–295 (1983).
Fanti, G. & Basso, R. The Turin Shroud, optical research in the past, present and future (Nova Science Publisher Inc., New York, 2007).
Fanti, G. Hypotheses regarding the formation of the body image on the Turin Shroud. A critical compendium. J. Imaging Sci. Technol. 55, 1–14 (2011).
Damon, P. E. et al. Radio carbon dating of the Shroud of Turin. Nature 337, 611–615 (1989).
Riani, M., Atkinson, A. C., Fanti, G. & Crosilla, F. Regression analysis with partially labelled regressors: carbon dating of the Shroud of Turin. Stat. Comput. 23, 551–561 (2013).
Rogers, R. N. Studies on the radiocarbon sample from the Shroud of Turin. Thermochimica Acta 425, 189–194 (2005).
Schwalbe, L. A. & Rogers, R. N. Physics and chemistry of the Shroud of Turin, a summary of the 1978 investigation. Anal. Chim. Acta 135, 3–49 (1982).
Riggi di Numana, G. Rapporto Sindone 1978/1987 (Ed. 3M, Milan, Italy, 1988).
Fanti, G. & Basso, R. Statistical analysis of dusts taken from different areas of the Turin Shroud (Ed. Libreria Progetto, Padua, Italy, 2009).
Frei, M. Il passato della Sindone alla luce della palinologia (In: La Sindone e la Scienza, Ed. Paoline, Turin, Italy, pp. 191–200, 1979).
Frei, M. Identificazione e classificazione dei nuovi pollini della Sindone (In: La Sindone, Scienza e Fede, Ed. CLUEB, Bologna, Italy, pp. 277–284, 1983).
Danin, A., Whanger, A. D., Baruch, U. & Whanger, M. Flora of the Shroud of Turin (Missouri Bot. Gard. Press, St. Louis, USA, pp. 1–52, 1999).
Andrews, R. M. et al. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nat. Genet. 23, 147 (1999).
Fanti, G. & Malfi, P. Sindone: primo secolo dopo Cristo (Ed. Segno, Udine, Italy, p. 403, 2014).
Nicolè, S., Barcaccia, G., Erickson, D.L., Kress, W. J. & Lucchin, M. The coding region of the UFGT gene is a source of diagnostic SNP markers that allow single-locus DNA genotyping for the assessment of cultivar identity and ancestry in grapevine (Vitis vinifera L.). BMC Res. Notes 6, 502 (2013).
Nicolè, S. et al. Biodiversity studies in Phaseolus species by DNA barcoding. Genome 54, 529–545 (2011).
Kress, W. J., Wurdack, K. J., Zimmer, E. A., Weigt, L. A. & Janzen, D. H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 102, 8369–8374 (2005).
Ratnasingham, S. & Hebert, P. D. N. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Molecular Ecology Notes 7, 355–364 (2007).
Willerslev, E. & Cooper, A. Review on ancient DNA. Proc. Royal Society B 272, 3–16 (2005).
Prüfer, K., Stenzel, U., Hofreiter, M., Pääbo, S., Kelso, J. & Green, R. E. Computational challenges in the analysis of ancient DNA. Genome Biology 11, R47 (2010).
Niemi, M. et al. Mitochondrial DNA and Y-chromosomal diversity in ancient populations of domestic sheep (Ovis aries) in Finland: comparison with contemporary sheep breeds. Genet. Sel. Evol. 45, 2 (2013).
Higgins, D., Rohrlach, A. B., Kaidonis, J., Townsend, G. & Austin, J. J. Differential nuclear and mitochondrial DNA preservation in post-mortem teeth with implications for forensic and ancient DNA studies. PLoS ONE 10 (5), e0126935 (2015)
Godzik, W. & Li, A. Cd-hit: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 22, 1658–1659 (2006).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics 25, 1754–1760 (2009).
Campagna, D. et al. PASS: a program to align short sequences. Bioinformatics 25, 967–968 (2009).
Kloss-Brandstätter, A. et al. HaploGrep: a fast and reliable algorithm for automatic classification of mitochondrial DNA haplogroups. Hum. Mutat. 32, 25–32 (2011).
van Oven, M. & Kayser, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 30, E386–E394 (2009).
Acknowledgements
The authors wish to thank Giulio Fanti, Department of Industrial Engineering, University of Padua, Italy, for providing all the samples used in this study for DNA investigations as sub-sets of the dust particles vacuumed from the back of the Turin Shroud. We also wish to thank Michela Verna, Laboratory of Plant Genetics, University of Padova, for her technical support in the laboratory analyses. This research was supported by the University of Padua: Progetto di Ateneo 2009 (CPDA099244) “Multidisciplinary analysis applied to the Shroud of Turin: A study of body image, of possible environmental pollution and of microparticles characterizing the linen fabric” (Giulio Fanti) and Progetto di Ateneo 2008 (CPDA087818) “Development of tools for the monitoring of biodiversity and the molecular identification of species and varieties in plants of agricultural and forest interest” (Margherita Lucchin and Gianni Barcaccia), by the University of Pavia: strategic theme “Towards a governance model for international migration: an interdisciplinary and diachronic perspective” (MIGRAT-IN-G) and by the Italian Ministry of Education, University and Research: Progetti Futuro in Ricerca 2012 (RBFR126B8I) (Alessandro Achilli and Anna Olivieri) and Progetti Ricerca Interesse Nazionale 2012 (Alessandro Achilli and Antonio Torroni).
Author information
Authors and Affiliations
Contributions
G.B. and G.G. conceived and designed the research. G.G., A.O., A.A., A.T. and G.B. analysed and interpreted the data. All the authors wrote and approved the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Barcaccia, G., Galla, G., Achilli, A. et al. Uncovering the sources of DNA found on the Turin Shroud. Sci Rep 5, 14484 (2015). https://doi.org/10.1038/srep14484
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep14484
This article is cited by
-
Blood stains of the Turin Shroud 2015: beyond personal hopes and limitations of techniques
Internal and Emergency Medicine (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}