
Abstract
Irrigation channels around paddy fields have been drastically improved over the past several decades in Japan. This has been accomplished exclusively by covering all the sides of channels with concrete. Although it is conventionally accepted that channel improvement has deteriorated quality of spawning and nursery sites for fish, to date, there is little evidence to support this claim. We examined the degree to which an improved irrigation channel with pools functioned as a spawning and nursery site for Silurus asotus and Cyprininae species within the Ezuragawa creek system that pours into Lake Biwa, Hikone, Shiga Prefecture, Japan. More eggs were observed for S. asotus than for Cyprininae species, but the opposite tendency was observed for the larvae and juveniles. Moreover, S. asotus juveniles were extremely rare. Therefore, our data suggest that improved irrigation channels are not suitable spawning and nursery sites for some species.
Similar content being viewed by others
Introduction
Several freshwater fish species use rice paddies as spawning and nursery sites for larvae as alternatives to temporarily flooded areas in Japan1,2,3,4,5. Despite their potential importance, rice paddies are often not accessible to fish inhabiting irrigation channels6,7 because of the considerable differences in elevation between the water surface of irrigation channels and the drainpipes from paddies. This may reduce the available spawning sites for some fish species2,3,4 even though improved irrigation channels can harbor pools, which might be utilized as spawning and nursery sites by some fish species. We examined this possibility for two taxonomic groups of fish, S. asotus and Cyprininae species (this term used for Cyprinus carpio and Carassius spp. in this article), both of which are thought to spawn and to grow up at the primary stage in rice paddies2,6,8.
Before this survey, we observed that S. asotus spawns in improved irrigation channels in which both sides and the bottom are mostly covered with concrete. Such sites, which are partly covered with aquatic weeds, are often regarded as unsuitable for spawning in comparison with other natural spawning sites such as rice paddies. However, to date, little is known about the extent to which such improved irrigation channels function as spawning sites. Suzuki et al. examined interspecific differences between the spawning sites of Cyprininae species/subspecies in Ezuragawa creek, an improved irrigation channel and identified larvae and juveniles of nine species based on morphology and random amplified polymorphic DNA (RAPD)9. Nevertheless, no studies have examined spawning of other fish species, such as S. asotus, which has been a serious concern due to the recent decline in Japanese fresh waters10.
Here, we examined the abilities of S. asotus and Cyprininae species, both of which spawn on the surface of submerged plants and release adhesive eggs11,12, to spawn in an improved irrigation channel. We selected these two groups for comparison and treated Cyprinus carpio and Carassius spp. as one group, Cyprininae species, given the considerable difficulty in distinguishing among the Cyprininae species at the egg and larval stages.
Results
Seasonal changes in environmental factors
Water temperature increased gradually during the study period (Fig. 1a). Water depth remained higher than 17 cm until mid-June, when it dropped by 5–10 cm until July, with the exception of flooding periods (Fig. 1b). The water flow velocity increased later season with water depth decrease (Fig. 1c). For conservation of fish spawning area, the water level of Lake Biwa was artificially maintained at 30 cm until 16 June 2008, after which it was dropped to −20 cm at Setagawa Weir, but we observed a gradual drop in water level from early June onwards. There was a significant positive correlation between the water level of Lake Biwa and the water depth of the study site (Spearman's rank test r = 0.62, P < 0.01).

Seasonal changes in environmental factors at sampling St. d during the study period: (a) water temperature, (b) water depth and (c) water flow velocity.
Slope and shear stress of the irrigation channel
The average slope of sections a–c was 0.0005, whereas that of sections d–h was 0.0013 (Fig. 2a). Shear stress in sections a–c was 1.75 N m−2, versus 2.26 N m−2 in sections d–h.

Slopes of the riverbed of the study area (a) and water depth at sampling St. a to h (b).
In (a),  and
and  indicates measured relative height in section a–c and section d–h, respectively.
indicates measured relative height in section a–c and section d–h, respectively.  show a point of steep drop. In (b),
show a point of steep drop. In (b),  show a station where set for regular census. Bar on st. a, c, d and h indicate water depth measured for calculation of shear stress on 30 May 2008.
show a station where set for regular census. Bar on st. a, c, d and h indicate water depth measured for calculation of shear stress on 30 May 2008.
Fish fauna and other large aquatic animals
The fish species collected during the regular censuses, the additional net samplings and trap samplings are shown in Table 1. A total of 591 individuals of 14 species, including S. asotus, Cyprinus carpio and Carassius spp. and eight families, were collected. The dominant species were C. carpio, Carassius spp., Oryzias latipes and Rhinogobius spp. In addition to fish, an alien crayfish Procambarus clarkii, shrimps, nymphs of dragonflies and damselflies, tadpoles and an invasive frog Rana catesbeiana were collected.
Silurus asotus and Cyprininae species
We collected a total of 1,210 S. asotus eggs on 30 of the 58 census days. S. asotus eggs were abundant from mid May to mid June (Fig. 3a). Similarly, a total of 308 Cyprininae eggs were collected from mid May to mid June (Fig. 3b), but were collected only on 11 of the 58 census days, indicating a significant difference in the occurrence of Cyprininae eggs in comparison with S. asotus eggs(Fisher's exact test, two-tailed, P = 0.0004).

Mean number of eggs ( ) and larvae or juveniles (
) and larvae or juveniles ( ) of S. asotus (a) and Cypirininae (b) over time.
) of S. asotus (a) and Cypirininae (b) over time.
Mean number was calculated by total number divide into the number of stations. The area size of one station is 3200 cm2.
A total of 22 individuals (larvae and juveniles) of S. asotus were collected on eight of the 58 census days. Despite collecting fewer Cyprininae eggs, a total of 136 larvae and juveniles of Cyprininae on 23 of the 58 census days, which were substantially more than those of S. asotus, were collected. There was a significant difference in the occurrence of eggs between Cyprininae and S. asotus (Fisher's exact test, two-tailed, P = 0.003). The body sizes of these larvae and juveniles were (average ± s.e.m. and range) 7.0 ± 6.3 mm and 3–21 mm and 12.9 ± 5.5 mm and 6–39 mm for S. asotus and Cyprininae, respectively. More than 95% of these fish were less than 20 mm in body length (Fig. 4). More than 70% of S. asotus and 40% of Cyprininae individuals were small fish (<10 mm in body length) (Fig. 4). Small fish of S. asotus were collected four times, versus 11 times for Cyprininae.

Size distribution of S. asotus ( : number of individuals collected,
: number of individuals collected,  : proportion of each size class) and Cyprininae (
: proportion of each size class) and Cyprininae ( and
and  , respectively).
, respectively).
Number of individuals is total number in 11 stations and the total sampling area is 35200 cm2.
Eggs were collected more frequently in section e–i, of the census areas for S. asotus, whereas more Cyprininae were found in section d–f (Fig. 5a, c). For S. asotus, a few larvae and juveniles were collected in section a–i (Fig. 5b). For Cyprininae, larvae and juveniles were collected most frequently in section a–c (Fig. 5d).

The number of eggs and of larvae and juvenile collected at each sampling station for S. asotus (a and c) and Cyprininae (b and d).
These numbers was mean number per day at each station. Error bar represent s.e.m.
Discussion
The leveling revealed that sections a–c exhibited substantially reduced shear stress compared to sections d–h. Shear stress of river bed at sections a–c (1.75 Nm−2) was about 77% of that at sections d–h (2.26 Nm−2). Many experimental results shows that grain size of uniform sand carried by a given shear stress is proportional to shear stress13. The difference in shear stress thus shows that mobility of a grain at sections a–c is smaller than that at sections d–h. Unfortunately, no such data are available for grain size and density of egg at present. The numerical analysis for mobility of the egg will be able to be possible if we can obtain the data of S. asotus and Cyprininae eggs.
Since the section exhibited reduced shear stress and pool-like environment, we predicted little egg movement in sections a–c. We collected 136 Cyprininae individuals. Suzuki et al. examined larvae and juveniles fish collected in Imagawa and reported the occurrence of three species of Cyprininae (C. carpio, C. auratus langsdorfii and C. a. grandoculis) based on RAPD analysis9. Additionally, they conducted fish sampling 10 times throughout the season and detected a trend of increasing body size over time based on 150, 316 and 118 individuals of C. a. grandoculis, C. a. langsdorfii and C. carpio, respectively. These results suggest that Imagawa provides alternative spawning and nursery sites for these Cyprininae species.
However, only one S. asotus individual was caught in 20059. We also found a substantially lower number of larvae and juveniles of S. asotus compared to Cyprininae, even though the opposite trend was seen at the egg stage. Moreover, small, recently hatched larvae (≤−10 mm) of S. asotus were collected only four times, versus 11 times for Cyprininae. These results strongly suggest that most S. asotus eggs either failed to hatch or did not survive after hatching.
Eggs of both S. asotus and Cyprininae are somewhat sticky11,12. In paddies, most eggs of Cyprininae adhere to weed culm. In contrast, eggs of S. asotus become covered with mud, camouflaging them6. Miyadi et al. reported that S. asotus spawned on the surface of floating water weeds, but that some eggs dropped onto the muddy creek bottom and hatched there14. Similarly, we found a large proportion of S. asotus eggs to be entangled with filamentous algae such as Spirogyra sp. and detected few eggs adherent to substratum, such as plant culm, due to adhesive power of the eggs. These results suggest that S. asotus eggs are less sticky than Cyprininae eggs and therefore that a substantial proportion of S. asotus eggs may flow downstream unless a suitable adherent substratum is available.
Both S. asotus and Cyprininae spawn and inhabit during early developmental period in temporal waters such as paddies, which may function as enemy free space15 for eggs and small fish. These fish, after gaining swimming ability in paddies, emigrates from paddies to permanent waters such as Lakes8. In the study area, Imagawa, it was practically impossible for fish to thrust into paddies, so it spawned in irrigation channels. Although a possibility remains that a small proportion of drifted eggs survived in the down stream, it was very unlikely to consider the low predation pressure there.
We observed numerous potential predators such as Opsariichthys platypus, Gnathopogon elongatus elongatus, Rhinogobius spp., Lepomis macrochirus, Procambarus clarkii and shrimps in the lower regions of the study area, where the water was relatively deep (Table 1). A previous study suggested that egg predation by L. macrochirus hindered the reproduction of Cyprininae16. This apparently increased predation pressure may explain why neither eggs nor newly hatched larvae survived well in the lower regions of the irrigation channels.
In contrast, other environmental conditions may be relatively suitable for fish growth in the middle and lower parts of the irrigation channels. As proof of this, many juvenile of Cyprininae were collected. In general, S. asotus and Cyprininae are regarded as similar groups of fish that use temporal bodies of water as spawning sites4,5. We consider that the large difference in survivorship between S. asotus and Cyprininae is survival rate in egg and larval stage and it may be attributable to differences in egg adherence.
The present study suggests spawning in stagnant water environments is more efficient than in currents. In addition, we confirmed the absence of S. asotus eggs even in pool-like environments where water velocity was low, suggesting substantial predation pressure in these areas. These results imply that the improved irrigation channels do not function as alternatives to paddy sites as spawning and nursery sites at least for S. asotus. In this context, it may be promising to prevent eggs from drifting in shallow water areas, providing with submerged plants as adhesive substratum for eggs. Experimental verification is required to establish effective conservation measures.
Methods
Study site
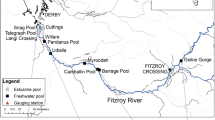
We conducted field censuses in Imagawa, a tributary of Ezuragawa, an irrigation channel running through Hassaka town, Hikone City, Shiga Prefecture, Japan (Fig. 6). Imagawa starts in Kaideima town, runs thorough paddies therein and after sharply turning southwest, pours into Ezuragawa 360 m upstream of the river mouth opening into Lake Biwa.

(a) Map of the study area near Lake Biwa, Japan.  indicate the direction of water flow. (b) Detail view of Study section.
indicate the direction of water flow. (b) Detail view of Study section.  denote the sampling stations (a to k). We created the map using Adobe Illustrator CS3 for Windows.
denote the sampling stations (a to k). We created the map using Adobe Illustrator CS3 for Windows.
Imagawa is an improved irrigation channel, with both sides covered with concrete for its whole length. Along Imagawa there was a small, 0.85-m tall weir, which prevented fish from traveling upstream at a point of 1.5 km up from the junction with Ezuragawa. The situation of Imagawa differs drastically in three parts, the lower, middle and upper parts.
The lower part, up to 300 m from the junction with Ezuragawa, consists of pools and gentle streams comprised of cobbles and pebbles 5 to 20 cm in diameter. In contrast, the middle and upper part is covered with concrete, with the exception of some scattered areas of pebble and mud.
The middle and upper parts are separated by a steep drop of approximately 0.15 m at point up to 460 m from the river junction. The lower and middle parts, below the drop, are very gentle, forming stagnant pools harboring emergent plants and submerged plants.
At Kaideima town, there are several welling points, from which spring water flows into Imagawa. Irrigation water pours into Imagawa along the paddy fields via irrigation ditches or directly from the paddies. We identified five irrigation ditches which drained stable amounts of water into Imagawa at 250 m, 690 m, 1,000 m, 1,220 m and 1430 m upstream from the junction with Ezuragawa. Fishes hardly intruded into paddies at any place due to fairly large differences in elevation between the irrigation channel and the paddies.
Leveling of the irrigation channel
To confirm that the flow within the lower part of the irrigation channel was sufficiently gentle to allow for spawning sites, we leveled the sections from St. a to h. A laser automatic level and a staff gauge were used and the relative height from the upper end of the drop at St. d was measured. Given that no small relief of the channel bed was found at the section between St. d and h, the relative height was measured at 20 m intervals in this area, whereas 5 m intervals were used in the section between St. a and c due to the small relief within this section. Further, a regression equation between distance and relative height was calculated for each of the two sections to obtain the average slopes.
Water depths measured at St. a, c, d and h on 30, May 2008 were used to determine the shear stress on the channel bed. Shear stress τ is given as

where ρ is the density of water, g is the gravitational acceleration, R is the hydraulic radius and I is the slope. Hydraulic radius R can be expressed as

where A is the cross sectional area of channel flow and S is the perimeter. Since the channel is rectangular, S can be given as

where H is water depth and W is channel width.
To calculate the shear stress, the average of the water depths obtained at St. a and c was used as the average water depth of the section between these stations. Similarly, the depths at St. d and h were averaged to represent the depth of the section between these two stations. In addition, ρ = 1000 kg m−3, g = 9.8 m s−2 and W = 3.1 m were used for the calculation.
Target species
S. asotus and Cyprininae which are target species in this study spawn in temporary waters. The areas have a risk to dry up. Therefore, these species have many and small egg and scatter their eggs to dispersion of the risk. These species, in Japan, have decreased in some decades with paddy fields had improved.
Regular census
We conducted 58 censuses over 79 days from 14 May to 31 July 2008, in the afternoon, from 15:00 to 17:00 h. Using a 40-cm wide and 30 cm depth fishing net (1 mm × 1 mm mesh) with a semicircular shape, the bottom of the irrigation channel was scraped upward at 40 cm × 40 cm three times at each station on the right and left side of each sampling site. Because the bottom of the irrigation channel was very flat, the sampling method enabled us to collect most of eggs and larvae. Size of the egg and larva are larger than mesh size, so they cannot get away through the meshes of the net. We counted the number of eggs and larvae (S. asotus or Cyprininae) within a sample spread on a plastic tray and measured the standard body length or the total body length for large larvae (≥−10 mm) and small larvae (≤−10 mm).
The censuses were conducted at 8 stations (from a to h) 40–50 m apart from 14 to 29 May and at 11 stations (from a to k) on and after 30 May (Fig. 6). Three additional stations (from i to k) were set up in the upper stream 200–280 m apart. St. a, b and c were set up in the middle part, which was gentle stream harboring emergent plants. Other 8 stations were set up in the upper part where is relatively shallow and lotic environments. The riverbed of stations is flat.
At St. d, water temperature, water depth, velocity and water flow were measured. The velocity was calculated by measuring the time at the 0.5 second rate for floating objects to move 1 m and the amount of water flow was determined by multiplying water depth, velocity and water width. The amount of water flow was used for an index to represent diurnal changes of water flow. We used data of the relative water levels of Lake Biwa measured by Kinki Regional Development Bureau Biwako Office, Japan Ministry of Land, Infrastructure, Transport and Tourism.
Additional net sampling and trap sampling
We conducted six additional censuses using fishing nets, the same type being used in the regular census, at St. a to d (24 June, 1, 8, 16 and 28 July and 14 August) to compensate sampling bias that may arise from different catch efficiency due to differences of species and of developmental stage fish of target species. To standardize the sampling effort, sampling time was set at 15 minutes divided by the number of samplers.
To collect large fish, we set up two traps (62 cm wide, 92 cm long and 54 cm tall) both being in a dome shape, consisting of 1 cm × 1 cm mesh net wall which prevented trapped fish from escaping.
These traps were set for 73 days from 21 May to 1 August 2008, during which we checked 59 times for fish.
References
Tomoda, Y. Lake Biwa And Catfishes. (Sekibun-sha, Tokyo, 1978). (In Japanese).
Saitoh, K., Katano, O. & Koizumi, A. Movement and Spawning of several freshwater fishes in temporary waters around paddy fields. Jpn. J. Ecol. 38, 35–47 (1988).
Saitoh, K. Ecological role of temporary waters for freshwater fish reproduction. In: Nagata, Y. & Hosoya, K. (eds) Circumstances In Endangered Japanese Freshwater Fishes And Their Protection. 194–204 (Midori-shobo, Tokyo, 1997). (In Japanese).
Katano, O. Fish assemblage of paddy fields and ditches. In: Ezaki, Y. & Tanaka, T. (eds) Conservation Of The Waterside Environment. 67–79 (Asakura-syoten, Tokyo, 1998). (In Japanese).
Maehata, M. Reproductive ecology of the Far Eastern catfish, Silurus asotus (Siluridae), with a comparison to its two congeners in Lake Biwa, Japan. Environ. Biol. Fish. 78, 135–146 (2007).
The Shiga Society of Naturalists. Flora And Fauna Of Rural Biotopes In Shiga. (Sunrise-syuppan, Shiga, 2001). (In Japanese).
Mori, A., Mizutani, M. & Takahashi, J. Characteristic and Deterioration of the Paddy Field Ecosystem: From the viewpoint of the Ecological Paddy Field Engineering. Trans. of J S I D R E. 254, 127–137 (2008).
Maehata, M. Ecotone around Lake Biwa –Role of rice paddy for fishes–. Mizu. 46, 26–36 (2004).
Suzuki, T., Kobayashi, T. & Ueno, K. Genetic identification of larvae and juveniles reveals the difference in the spawning site among Cyprininae fish species/subspecies in Lake Biwa. Environ. Biol. Fish. 82, 353–364 (2008).
Mizutani, S. What are the features of haruno-ogawa? In: Mizutani, S., Mori, A. (eds) Conserving Habitat Of Freshwater Fishes Inhabiting Haruno-ogawa, Irrigation/drainage Ditches, In Rice Paddies. 1–6 (Gakuho-sha, Tokyo, 2009). (In Japanese).
Kobayakawa, M. External Characteristics of the Eggs of Japanese Catfishes (Silurus). Japan J. Ichthyol. 32, 104–106 (1985).
Kawanabe, H., Mizuno, N. & Hosoya, K. Freshwater Fishes Of Japan. (Yama-kei, Tokyo, 2005). (In Japanese).
Takayam, S. River Morphology. (Kyoritsu-syuppan, Tokyo, 1974). (In Japanese).
Miyadi, D., Kawanabe, H. & Mizuno, N. Colored Illustrations Of The Freshwater Fishes Of Japan. (Hoiku-sha, Osaka, 1976). (In Japanese).
Jeffries, M. J. & Lawton, J. H. Enemy free space and the structure of ecological communities. Biol. J. Linnean. Soc. 23, 269–286 (1984).
Yamamoto, S. Impact of egg predation by bluegill Lepomis macrochirus on recruitment of carp Cyprinus carpio and crucian carp Carassius carassius sp. Report of Nagano Prefectural Fisheries Experimental Station 7, 16–20 (2005).
Acknowledgements
We appreciate S. Kanao, Lake Biwa Museum, for their generous advice and support of this survey. We also thanks D. Kado, K. Nakanishi and K. Tawa, The University of Shiga prefecture Graduate School of Environment Science, for their help in field research.
Author information
Authors and Affiliations
Contributions
T.F., T.N. and H.S. designed the study. T.F. performed the field work. T.F. and T.N. wrote the main manuscript text and prepared figures. Y.K. wrote the part of leveling method. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Funao, T., Nishida, T., Kurashige, Y. et al. Different suitability of improved irrigation channels as reproductive sites for Cyprininae and Silurus asotus. Sci Rep 4, 3588 (2014). https://doi.org/10.1038/srep03588
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03588
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
