
Abstract
The so-called man-eating wolves of Turku, a pack of three wolves, reportedly killed 22 children in South-Western Finland in 1880–1881. Enormous efforts were carried out to eradicate them. In January 1882 the last remaining wolf was killed. Since then, there has been considerable debate regarding the validity and extent of the man-eating behaviour. This study aims to clarify whether man-eating behaviour can be observed from the remains of these wolves. One of the wolves was mounted in 1882 and is on display at St. Olaf’s school in Turku, enabling us to collect hair keratin samples. Additionally, hair keratin was collected from two other suspected man-eaters. We analysed carbon (δ13C) and nitrogen (δ15N) stable isotope values to study the wolf’s diet during the last months of its life. Samples from seven temporally concurrent wolves were used to construct reference values. Our analyses indicated that δ15N values of suspected man-eaters were relatively low compared to the reference sample. We could not detect clear trends in isotope ratios associated with potential man-eating behavior. We believe that this lack of distinctive patterns can be explained by the relatively minor role that man-eating played in their overall diet.
Similar content being viewed by others

Introduction
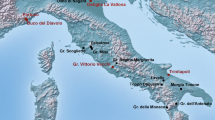
In 1880 and 1881 the so-called man-eating wolves of Turku reportedly killed 22 children1,2 in Southwestern Finland (Fig. 1). The first incident occurred on 18 January 1880, in Vellua village, 50 km northwest of Turku, the former capital of Finland. Two wolves were spotted from a neighboring house crossing a potato field and carrying something. That something turned out to be a boy named Karl Johan Hörnberg, later found dead and partly eaten in a nearby forest3.

A modern map of Finland presenting the relevant sites. Man-eating occurred in Turku region and our reference wolf sample are mainly from Kymi Valley and Hamina. Our archeological reference human samples originated from Rauma. Map: Public domain, Wikimedia Commons, modified by Tiina Väre.
During the following 22 months, the wolves were reported to have attacked 26 other children and one adult in small countryside villages north and northeast of Turku. In total, 22 children aged 2–10 years died1,2. Most of them were eaten at least partially. Large-scale wolf hunts were organized during the summers of 1880 and 1881 to eradicate this pack. Bounties placed by both the Senate and the county councils lured professional hunters to the area in the wintertime. Poisoned carcasses, trapping, and killing of the cubs during the spring thinned out the local wolf population. However, it took 2 years and even a group of lukashi—professional wolf hunters from Russia—to track and kill the alpha pair of the man-eating pack in January 18821,2,4. The cubs of the pack were killed during the summers of 1880 and 1881, with only one cub surviving until the end of October 18811,5.
The alpha female of the pack, killed on 3 January 18826, was found to be in poor condition, with worn-out teeth and a shaggy, yellow-brown fur1. The fur of the female was preserved but is believed to be currently missing. The alpha male was killed on 14 January 1882 by a son of a tenant farmer, Matti Hillberg, who sold the wolf’s hide for mounting7. The mounted wolf was later donated to a local school in Turku, where it is still preserved2. By the time of the killing, the male was reportedly tired, lame, and walking on three legs, apparently because he had been shot at the hind leg during the hunt on 3 January 18821. The exact size of the wolf pack engaging in man-eating behavior is, however, uncertain. The man-eating pack could have consisted of just the alpha pair, the alpha pair and their cub(s), or possibly also other wolves in the area.
Wolf diets and potential man-eating behavior
In Finland, European moose (Alces alces alces) is the main source of nutrition of wolves8,9. However, wolves are opportunistic if their preferred prey is scarce or unavailable10,11. In the late nineteenth century, extensive hunting had seriously reduced the moose population in southern Finland12, and it is likely that wolves of Turku hunted small prey animals and raided local farms for nutrition. For example, in 1878, 12 pigs, 2455 sheep, 472 cows, and 158 horses were lost to predators in the Turku region2. Limited prey availability is observed to be one of the main factors associated with man-eating behaviour in carnivores13. It is thus not difficult to understand the idea that in the absence of prey, the wolves would have resorted to hunting children.
Stable isotope analyses
Each tissue stores the stable isotope composition representing the diet conditions during its formation14,15. In this paper, the ratios of the stable isotopes of carbon (13C/12C) and nitrogen (15N/14N) in body hair keratin have been used to reflect the dietary habits of wolves during the period of hair growth16.
Wolves molt once a year, during the late spring (May–June), when they shed their warm winter fur. In the autumn, they grow a thick underfur for the cold winter months. Several factors are associated with the body hair growth in wolves. Hair growth rate vary among individuals and among different body parts of the same animal17. The growth rate of body hair of captive wolves was found to be 1.0 ± 0.07 mm/day in the summer and 0.63 ± 0.23 mm/day in the autumn16.
The isotopic compositions of the samples are measured in parts per thousand (‰) and are expressed with a delta (δ) notation, indicating the relative difference of isotope ratios between the sample and a standard (Vienna Peedee Belemnite for carbon and AIR for nitrogen). Earlier studies have demonstrated that the remains of man-eating carnivores can be utilized to study such behavior in detail. For example, stable isotope analyses were used with famous Tsavo lions to reveal the actual extent of their man-eating behavior18. The feasibility of determining a diet from partitioned hair samples has been studied in a controlled feeding study of captive wolves16.
In healthy, well-nourished individuals the ratio of nitrogen stable isotopes (ẟ15N) of most tissues can be expected to reflect the consumption of animal proteins14,15,19,20. Additionally, it can be used to determine the trophic position of the organism by comparing the values to local faunal data, considering trophic enrichment factors of 3–5‰21.
The ratio of carbon stable isotopes (δ13C) allows studying whether the analysed organism consumed plants using the C3 or C4 photosynthetic pathway or other organisms consuming C3- or C4-plants and to trace marine influences in the diet22,23,24,25. In brackish water environments, such as the Baltic Sea, as in freshwater environments, the carbon values likely resemble those of terrestrial C3 environments26. Thus, locally distinguishing between marine and terrestrial environments based on δ13C values is not possible. On the other hand, in addition to ẟ15N values, starvation may affect δ13C values27,28,29,30. Another, more geographically relevant factor possibly shaping the δ13C values is the canopy effect causing the ground level plants to be depleted in 13C compared to those higher in the canopies31. Also lichen-eating reindeers as a part of the trophic web can slightly elevate the δ13C values32, but reindeer are not present in the southern region of Finland, where the wolves of this study originate from.
In this study, we conducted experiments to determine whether man-eating could be observed from stable isotope values in hair samples taken from the alpha male, the St. Olaf’s school wolf, as well as two other wolves, potentially other individuals from the man-eating wolf pack. The δ13C and δ15N values in body hair keratin reflect dietary habits during the last couple of months prior to the death. Our hypothesis was that comparing the stable isotope values of the potential man-eater’s hair keratin to those measured in a concurrent reference group would allow us to further assess the man-eating behaviour. Significant consumption of human tissue should produce statistically different δ15N values in the hair keratin compared to our reference samples.
Results
The volume of human biomass consumed
The amount of consumed human tissue was approximated from the weights of the children by their age. The month starting from June 15th onwards included four confirmed human kills by the wolves. A 6-year-old boy was half-eaten (equating 8–12 kg of human tissue)33 as well as parts of an 8-year-old boy (~ 10 kg)34, a 2-year-old boy almost entirely (~ 10 kg)35 and parts of an 8-year-old girl (~ 8 kg)36.
During the month from August 15th onwards, only two kills could be confirmed. A 4-year-old boy was killed and concealed, but not eaten37 and a 9-year-old boy was partially eaten (~ 9 kg)38. The next month, starting from September 15th, included just one kill. An 8-year-old boy was almost completely eaten (~ 22 kg)39.
The month from October 15th onwards included two kills. A 7-year-old girl was killed and dragged away, but wolves were disturbed, and they could only eat parts of the girl (~ 9 kg)40. Moreover, a 5-year-old boy was killed on November 7th but only partially eaten, potentially mainly organs (~ 6 kg)41. This was the last human kill by the wolves, and there was approximately a two-month gap in human kills until the alfa pair was killed. During their last six months of existence, we estimated that the man-eating wolf pack of Turku could have been able to obtain approximately 85 kg of human biomass in their diet.
Stable isotopes
The keratin C:N atomic ratios for two wolves in the control group were outside the range (3.0–3.8) for keratin suggested by O’Connell and Hedges for human hair keratin20. However, unlike for collagen, the atomic C:N ratio of keratin is not considered as a strong indicator of preservation which is why these two samples were chosen to be included in the analyses. All other hair samples were within the suggested range for hair keratin (Table 1).
Due to the small sample sizes, the nonparametric Mann–Whitney test was selected to analyze whether the suspected man-eating wolves presented different stable isotope values than the wolves in the reference group. The δ15N value in the keratin of the St Olaf school’s wolf and Forssa wolf were the lowest in comparison to the reference group (Table 1; Fig. 2A). According to this test, the keratin values of the suspected man-eaters (n = 3) statistically deviate from those of the reference group (n = 7) (δ13C p 0.033; δ15N p 0.033). This suggests that the isotopic δ13C and δ15N composition of the diet consumed by the suspected man-eaters during the hair development was dissimilar to the reference group containing a clearly smaller portion of protein from higher trophic positions. This is not in line with the consummation of flesh of humans who typically occupy higher trophic positions.

δ13C and δ15N values of the wolves and the reference fauna. (A) The modified keratin δ13C and δ15N values of the reference wolves, suspected man-eating wolves (359 and Forssa) and the average value for the man-eating wolf from St. Olaf school from the sequentially cut hairs. The faunal background bone/dentin collagen isotopic data is from archaeological (Middle Ages to Early modernity) or ecological (pre-1950) animal remains from Finland49,55,56,57,58) The human dentin collagen isotopic data is from the early nineteenth century population of Rauma located near Turku44. (B) The δ13C and δ15N isotope values in the sequential hair samples of the St Olof school’s wolf. The variation is 0.2‰ for δ13C and 0.3‰ for δ15N while the analytical error for both is 0.1‰.
Both the δ13C and δ15N values of the St Olaf school’s wolf showed very little variation across the hair shaft (Table 2; Fig. 2B). The increase in nitrogen levels during the entire period was only 0.3‰, and the carbon values among sequences varied even less, with the variation remaining at 0.2‰ which could be nearly completely explained by analytical imprecision.
In addition to hair samples, isotopic data was attempted to obtain from a bone of the taxidermized St. Olaf’s school wolf. The atomic C:N ratio (4.1) of the bone collagen sample fell outside the accepted range of 2.9–3.6 suggested by DeNiro42, indicating dissatisfactory collagen quality. Thus, the obtained values (δ13C − 22.8‰; δ15N 8.2‰) were not included in the analysis. This is unfortunate as they would have reflected the long-term average diet of the wolf and could have been used as the baseline with which to compare the keratin values.
Discussion
The relatively low keratin δ15N values of the St. Olaf’s school’s wolf are not well aligned with the suspected man-eating behaviour, which should have elevated the trophic level of the wolf. The lack of variation in the values across the hair shaft implies a steady diet during the period of the hair growth. The δ15N values of the two other suspected, but less well documented man-eaters, wolf 359 and Forssa wolf, were somewhat lower in relation to the reference group.
An adult wolf with a body weight of 35 kg is estimated to require 3.25 kg of food per day. There is a significant variation in observed daily food consumption levels; depending on food abundance, the consumed amount could be even more than 10 kg/day43. The wolf pack under study consisted of an adult alfa pair, to which an offspring was added during the summer of 1881. Based on the daily food requirements, it is obvious that an estimated human biomass of 85 kg can explain just part of the diet of these wolves. It could comprise roughly 10–15% of their diet during the months of most frequent kills as the total food requirement for the pack should have been at least 200 kg/month.
Our results cannot conclusively support or rule out the man-eating behavior of the studied wolves. While the hair keratin 15N values were depleted relative to the human dentine values suggest that human flesh likely did not play a significant role in their diets during their last months, small portions would not necessarily have elevated the δ15N values, particularly if the diet primarily consisted of low 15N content foods, like ungulates (Fig. 2A; Appendix 1). If man-eating behaviour would have played a key role in diet, we should expect values elevated in comparison to the local level of human values. The average δ15N value of 11 samples of archaeological dentin collected from cementoenamel junction of permanent second premolars of individuals aged around 7–8 years from early nineteenth century Rauma was 11.0‰44 (Fig. 1; Appendix 1). Dentine develops during childhood45 and its isotope composition reflects the dietary conditions of the growing period14,15. On the other hand, food shortages of predators are not only connected to man-eating behaviour18,46 but may urge wolves to eat more plant-based nutrition47. Even normally, the occurrence of berries in wolfs diet in Finland varies between 1–3%9. This could have significantly dragged down the δ15N values.
Much like the Tsavo man-eating lion case18, the last months of the St. Olaf’s school wolf are well documented, and we have relatively accurate records of the number and age of the children killed and eaten. However, the proportion of the human tissue in the diet of Tsavo lions was up to 30%, clearly more than the 10–15% estimated for the man-eating wolves. Such a large portion is much more likely to show up in the stable isotope composition of the hair keratin and additionally, in the study by Yeakel and colleagues18, the possibility to compare the results with values of the bone collagen provided more potential for interpretation.
Ungulates and moose in particular are the main source of nutrition for wolves8,9, but in the nineteenth-century, European moose (Alces alces alces) population in Finland was markedly reduced12. However, we could not detect isotopic signs of starvation, which in inert tissues may be observed as δ15N value elevation and sometimes as simultaneous depletion of 13C27,28,29,30. Rather, the δ13C and δ15N values of both the suspected man-eaters, as well as those of the references group are well in line with a diet possibly containing a wide variety of herbivorous and omnivorous animal species from terrestrial C3 plant-based community10,11 (Fig. 2A; Appendix 1). As mentioned earlier, in year 1878 alone, 12 pigs, 2455 sheep, 472 cows, and 158 horses were reportedly lost for predators, probably mostly wolves. The δ15N and δ13C values of the suspected man-eating wolves are well in line with consuming domesticated species (Fig. 2A; Appendix 1). The transition to killing and eating children may have been easy for wolves familiar to proximity of humans via their domestic animals.
The isotopic ratios of nitrogen also suggests inclusion of some aquatic species from brackish or freshwater environments—such as fish-eating waterfowl—as a part of the diets of the wolves particularly in the control group48. The δ15N values are at a high level in comparison to modern wolf values although even much higher levels have been obtained, e.g. up to 12.9‰ in Alaskan wolves relying much on Pacific salmon48. In this geographic region, the δ13C values were not helpful in reliably distinguishing between marine and terrestrial/freshwater origin26,49 but typically, the tissues of species at high levels of the long and complex aquatic food-webs are enriched in 15N24,50.
As a main limitation of this study, we can only estimate the amount of human biomass consumed by the St. Olaf’s school wolf. It likely covered less than a tenth of what was nutritionally required by the pack. It is also unlikely that all the human tissue would have been consumed by an individual wolf. As the wolf pack of Turku consisted of an adult alfa pair and their offspring, there were two or three wolves sharing the kill. We can only assume how they distributed their prey. The bone collagen stable isotope results of the St Olaf’s wolf could have provided information on the baseline diet of that individual against which to reflect the keratin analysis results, but unfortunately the analyses were unsuccessful.
The man-eating behavior of carnivores is often connected to problems with their usual diet. This may have been the case also with the man-eating wolves of Turku. The significant reduction of the moose population in the late nineteenth century, the main prey of wolves, may have led some wolves to rely on atypical nutrition. The δ13C and δ15N values of the suspected man-eaters, align well with a diet that contained a variety of herbivorous and omnivorous animal species from terrestrial C3 plant-based community. These likely included domestic animals, but also the man-eating behavior remained as a possibility as it would have formed just a minor part of the total diet and thus could not significantly affect the isotopic values.
Methods
Human biomass
Man-eating wolves killed 22 children aged 2–10 years in 1880 and 1881. Although these killings are well documented, assessing the potential amount of human tissue or biomass consumed by the alpha male presents several challenges. Firstly, most of the prey was only partially consumed by these wolves as they were often disturbed while eating, resulting in the incomplete consumption of the body1. Secondly, while the age and sex of the victims are known, there may be a significant variation in body size among individuals of the same age. Additionally, the body size of children from the 1880s is not directly comparable to modern-day values. In our estimations, we opted to use modern-day children’s growth charts from Finland, and instead of using the normal growth rate values directly, we subtracted them by two standard deviations.
Stable isotope analyses
The stable isotope ratios of carbon and nitrogen were analyzed from the keratin of body hair collected from the taxidermized hide of the suspected man-eating alpha male from St. Olaf’s school. Taxidermy has previously been found not to affect the stable isotope values51. The hair was analyzed in c. 1 cm sequences to scrutinize any possible temporal change in diet during the growth period of the hair. The minimum length of this hair was 6 cm. An additional seventh sample from the remaining hair tips of varying lengths was analysed, representing the furthest time from the animal’s death. Based on the deduced growth rate of 0.63 mm/day16, the isotopic composition of these hairs can be assumed to reflect the dietary conditions of roughly the last 4 months of the wolf’s life, approximately the period beginning from September.
Additionally, an attempt was made to collect a bone sample from the mandible of the taxidermized St. Olaf’s school wolf. The bone collagen extraction was performed using suction filtering according to a modified Longin method introduced by Brown and colleagues52 and analysed in the same patch with the hair samples.
In addition, we analysed two wolves possibly involved in man-eating but with much less documented history than the St. Olaf’s school wolf. These samples were a taxidermized wolf 359 from Turku-region (late nineteenth century; the collections of Lammi Biological station, University of Helsinki) and a wolf hide originally from Turku region (late nineteenth century; the collections of the Forssa Nature Museum). The exact dates of the hunting of these individuals are missing, but wolf 359 is marked as “man-eater” in the collection, and the hide from the Forssa Nature Museum is also suspected to have potentially originated from a wolf belonging to a man-eating pack. The hair sample of wolf 359 was obtained from the mid-shafts of the hair while the Forssa sample was complete hairs due to their shortness, and thus they represent the diet some months prior to the death.
Nevertheless, rather than merely the most recent diet, stable isotope signals also convey the composition of the organism’s former amino acid pool, the primary source for protein synthesis. This buffering effect mitigate immediate dietary changes from fully manifesting in tissues, although some nitrogen atoms from the new diet will gradually begin to incorporate among the old ones, altering the composition of growing tissues20. Therefore, even the earlier human casualties may be relevant in this case.
The reference group of concurrent wolves consists of samples taken from six wolf hides belonging to the collections of The Hunting Museum of Finland. The provenance of these subjects is uncertain but they all date to the nineteenth century and at least four of them were shot by Anders Edvard Baskin presumably in 1860–1880, in Kymi Valley. One taxidermized wolf 400 (coll. 29.9.1845, Pornainen) was from the collections of Lammi Biological station, University of Helsinki. The reference samples from the control group were obtained from the mid-shafts of the hair, and they represent the diet some months prior to the death.
The wolf hair samples were prepared for the carbon and nitrogen stable isotope analyses at the Archaeological Research Laboratory, Stockholm University, Stockholm, Sweden, with the exception of the Forssa Nature Museum sample, which was prepared in the Nuclear Research Department, Center for Physical Sciences and Technology, Vilnius, Lithuania. To remove lipids, they were ultrasonicated for 3 × 15 min in a chloroform–methanol solution (2:1) following the modified protocol of Britton and colleagues53. Subsequently, the hair was rinsed of any residue by ultrasonically cleaning them for 1 × 15 min in ultrapure water and left to dry for 24 h, before being cut and weighed into tin cups in samples ranging from 0.7 to 1.1 mg.
The isotope analyses were conducted at the Nuclear Research Department, Center for Physical Sciences and Technology, Vilnius, Lithuania. Samples were transferred to the tin capsules and analysed using Flash EA 1112 series Elemental analyzer that was connected to the Delta V Advantage Isotope Ratio Mass Spectrometer (IRMS) via ConFlo III interface (all from Thermo, Bremen, Germany). The Elemental analyzer consists of oxidation and reduction furnaces (operating at 1020 °C and 650 °C, respectively), a chromatographic column (PoraPlot Q), a water absorption column, and a TCD detector. During the analysis, N% and C% were determined using the Elemental analyzer, and later gases were passed to the IRMS for the stable isotope ratio measurements. Ratios of 15N/14N and 13C/12C were expressed relative to the international standards, atmospheric air (N) and Pee Dee Belemnite ©, and presented in a δ-notation as parts per thousand difference from the standard. Caffeine IAEA-600 was used as a secondary reference material (δ15N = + 1‰, δ13C = − 27.771‰); the analytical precision was 0.1‰ for both δ15N and δ13C.
Statistical analyses, including testing for normal distribution of the δ15N and δ13C values and comparing the St. Olaf’s school wolf and two other potential man-eaters with the reference material with Mann–Whitney tests, were conducted by using SPSS (IBM, Armonk, NY, USA) version 25, 64-bit edition. P-values were considered statistically significant if they were smaller than 0.05.
We also compared the hair keratin stable isotope values of the studied wolves to bone or dentine collagen stable isotope values of wild and domestic herbivores, domestic omnivores, and birds from the dIANA database (Appendix 1). Additionally, we compared them to the early nineteenth century human population of the town of Rauma located near Turku44 (Fig. 1). To make this comparison, keratin values were modified to correspond to the more frequently used collagen values, following the suggestions of O’Connell and Hedges20 by adding + 0.5‰ to the δ13C values and 1‰ to the δ15N values. Additionally, the δ13C values were adjusted for the SUESS effect according to the table presented by McCarroll and Loader54.
Data availability
All data is available upon reasonable request from the corresponding author.
References
Godenhjelm, U. Minnen från vargåren i Åbo län, J (Simeliuksen kirjapaino, 1891).
Tikkanen, J. Lauma—1880-luvun lastensurmat ja susiviha Suomessa (Kustannusosakeyhtiö Otava, 2019).
KA (National Archives of Finland) TPLLH, Eae:1178 kd 3/238.
Teperi J. 1977. Sudet Suomen rintamaiden ihmisten uhkana 1800-luvulla. SHS, Helsinki.
KA (National Archives of Finland) TPLLH Eae:1191 kd 10/333.
KA (National Archives of Finland ) MTK (Maataloustoimituskunta) Ea:116 kd 24/459.
KA (National Archives of Finland) TPLLH, lGac:592, 3787.
Kojola, I. Suden, hirven ja metsästyksen vuorovaikutussuhteet. Suomen Riista 46, 76–81 (2000).
Gade-Jorgensen, I. & Stagegaard, R. Diet composition of wolves Canis lupus in East-Central Finland. Acta Theriol. 45(4), 537–547 (2000).
Zlatanova, D., Ahmed, A., Vlasseva, A. & Genov, P. Adaptive diet strategy of the wolf (Canis lupus L.) in Europe: A review. Acta Zool. Bulg. 66, 439–452 (2014).
Newsome, T. M. et al. Food habits of the world’s grey wolves. Mamm. Rev. 46, 255–269 (2016).
Nygrén, T. The history of moose in Finland. Swed. Wildl. Res. Suppl. 1, 49–54 (1987).
DeSantis, L. & Patterson, B. Dietary behaviour of man-eating lions as revealed by dental microwear textures. Sci. Rep. 7, 904. https://doi.org/10.1038/s41598-017-00948-5 (2017).
DeNiro, M. J. & Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 42, 495–506 (1978).
DeNiro, M. J. & Epstein, S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 45, 341–351 (1981).
McLaren, A. A. D., Crawshaw, G. J. & Patterson, B. R. Carbon and nitrogen discrimination factors of wolves and accuracy of diet inferences using stable isotope analysis. Wildl. Soc. Bull. 39, 788–796 (2015).
Gunaratnam, P. & Wilkinson, G. T. A study of normal hair growth in the dog. J. Small Anim. Pract. 24, 445–453 (1983).
Yeakel, J. et al. Cooperation and individuality among man-eating lions. PNAS 106, 19040–19043 (2009).
O’Connell, T. C. & Hedges, R. E. M. Isotopic comparison of hair and bone: Archaeological analyses. J. Archaeol. Sci. 26, 661–665 (1999).
O’Connell, T. C. & Hedges, R. E. Investigations into the effect of diet on modern human hair isotopic values. Am. J. Phys. Anthropol. 108(4), 409–425 (1999).
Hedges, R. & Reynard, L. Nitrogen isotopes and the trophic level of humans in archaeology. J. Archaeol. Sci. 34, 1240–1251 (2007).
Minagawa, M. & Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 48, 1135–1140 (1984).
Schoeninger, M. J. & DeNiro, M. J. Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim. Cosmochim. Acta 48, 625–639 (1984).
Ambrose, S. H. Isotopic analysis of paleodiets: Methodological and interpretive considerations. In Investigations of Ancient Human Tissues. Chemical Analyses in Anthropology (ed. Sandford, M. K.) 59–130 (University of North Carolina of Greensboro, 1993).
Kohn, M. J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc. Natl. Acad. Sci. USA 107(46), 19691–19695 (2010).
Katzenberg, M. A. Stable isotope analysis of archaeological faunal remains from southern Ontario. J. Archaeol. Sci. 16, 319–329 (1989).
Lee-Thorp, J. A., Sealy, J. C. & van der Merwe, N. J. Stable carbon isotope ratio differences between bone collagen and bone apatite, and their relationship to diet. J. Archaeol. Sci. 16, 585–599 (1989).
Mekota, A. M., Grupe, G., Ufer, S. & Cuntz, U. Serial analysis of stable nitrogen and carbon isotopes in hair: monitoring starvation and recovery phases of patients suffering from anorexia nervosa. Rapid Commun. Mass Spectrom. 20, 1604–1610 (2006).
Mekota, A., Grupe, G., Ufer, S. & Cuntz, U. Identifying starvation episodes using stable isotopes in hair. Rechtsmedizin 19, 431–440 (2009).
Doi, H., Akamatsu, F. & González, A. L. Starvation effects on nitrogen and carbon stable isotopes of animals: An insight from meta-analysis of fasting experiments. R Soc. Open Sci. 4, 170633 (2017).
Drucker, D. G., Bridault, A., Hobson, K. A., Szuma, E. & Bocherens, H. Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 266(1–2), 69–82 (2008).
Drucker, D., Bocherens, H., Pike-Tay, A. & Mariotti, A. Isotopic tracking of seasonal dietary change in dentine collagen: Preliminary data from modern caribou. Comptes Rendus Acad. Sci. Ser. IIA Earth Planet. Sci. 333(5), 303–309 (2001).
KA (National Archives of Finland) TPLLH (Turun ja Porin läänin lääninhallitus) Eae:1192 kd 118/336.
KA (National Archives of Finland) TPLLH Eae:1192 kd 123/360.
KA (National Archives of Finland) TPLLH Eae:1192 kd 132/361.
KA (National Archives of Finland) TPLLH Eae:1191 kd 6/333.
KA (National Archives of Finland) TPLLH Eae:1191 kd 15/338.
KA (National Archives of Finland) TPLLH Eae:1191 kd 18/339.
KA (National Archives of Finland) TPLLH Eae:1192 kd 145/365.
KA (National Archives of Finland) TPLLH, Eae 1191 kd 10/333.
KA (National Archives of Finland) TPLLH Eae:1192 kd 193/367.
DeNiro, M. J. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809 (1985).
Stahler, D., Smith, D. & Guernsey, D. Foraging and feeding ecology of the gray wolf (Canis lupus): Lessons from Yellowstone National Park, Wyoming, USA. J. Nutr. 136, 1923S-1926S (2006).
Väre, T., Fjellström, M., Arppe, L., Núñez, M. & Lidén, K. Diets in three late medieval to early modern coastal populations in Finland according to the δ13C and δ15N values of bone and dentin collagen. Fennoscandia Archaeologica XXXIX, 96–117 (2022).
AlQahtani, S. J., Hector, M. P. & Liversidge, H. M. Brief communication: The London atlas of human tooth development and eruption. Am. J. Phys. Anthropol. 142(3), 481–490 (2010).
Packer, C., Ikanda, D., Kissui, B. & Kushnir, H. Conservation biology: Lion attacks on humans in Tanzania. Nature 436(7053), 927–928 (2005).
Gable, T. D., Windels, S. K. & Bruggink, J. G. Estimating biomass of berries consumed by gray wolves. Wildl. Soc. Bull. 41, 129–131 (2017).
Szepanski, M. M., Ben-David, M. & Van Ballenberghe, V. Assessment of anadromous salmon resources in the diet of the Alexander Archipelago wolf using stable isotope analysis. Oecologia 120(3), 327–335 (1999).
Etu-Sihvola, H. et al. The dIANA database—Resource for isotopic paleodietary research in the Baltic Sea area. J. Archaeol. Sci. Rep. 24, 1003–1013 (2019).
O’Brien, D. M. Stable isotope ratios as biomarkers of diet for health research. Annu. Rev. Nutr. 35, 565–594 (2015).
Koehler, G. & Hobson, K. A. Effects of tanning on the stable isotopic compositions of hair. Forensic Sci. Int. 292, 78–82 (2018).
Brown, T., Nelson, D., Vogel, J. & Southon, J. Improved collagen extraction by modified Longin method. Radiocarbon 30(2), 171–177 (1988).
Britton, K. et al. Maritime adaptations and dietary variation in prehistoric Western Alaska: Stable isotope analysis of permafrost-preserved human hair. Am. J. Phys. Anthropol. 151, 448–461 (2013).
McCarroll, D. & Loader, N. Stable isotopes in tree rings. Quatern. Sci. Rev. 23, 771–801 (2004).
Bläuer, A. et al. Inferring prehistorical and historical feeding practices from δ15N and δ13C isotope analysis on Finnish archaeological domesticated ruminant bones and teeth. Fennoscandia Archaeologica 2016, 171–188 (2016).
Lahtinen, M. & Salmi, A.-K. Mixed livelihood society in Iin Hamina: A case study of medieval diet in the Northern Ostrobothnia, Finland. Environ. Archaeol. 24(1), 1–14 (2019).
Salmi, A. K. & Heino, M. T. Tangled worlds: The Swedish, the Sámi, and the Reindeer. Int. J. Histor. Archaeol. 23, 260–282 (2019).
Oinonen, M. et al. Buried in water, burdened by nature—Resilience carried the Iron Age people through Fimbulvinter. PLoS One 15(4), e0231787 (2020).
Funding
This study was funded by Academy of Finland (No. 323428), Eino Jutikkala Foundation and Kone Foundation.
Author information
Authors and Affiliations
Contributions
J.A.J., J.T., T.K.S., M.N. and J.A. designed the study. J.A.J. and I.K. collected the samples. T.V. prepared the samples, J.A.J., T.V., J.H. and J.H. analyzed the data. J.A.J., T.V., M.H., M.N., J.H., L.K., J.H. and J.A. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Junno, JA., Väre, T., Tikkanen, J. et al. Stable isotope analyses of carbon and nitrogen in hair keratin of suspected man-eating wolves from 1880s. Sci Rep 14, 4946 (2024). https://doi.org/10.1038/s41598-024-55521-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-55521-8
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
