
Abstract
Associations between the STin2 and 5-HTTLPR polymorphisms within the serotonin transporter gene, SLC6A4, and culture across societies were examined. Based on an analysis of 75 primary studies (28,726 individuals), STin2 allelic frequencies were found to vary widely across countries, ranging from 26% in Germany to 85% in Singapore. Across 53 countries, and after controlling for all major environmental influences of culture, STin2 and 5-HTTLPR were found to explain 23.6% unique variance in monumentalism but none in individualism. Our findings evidence a significant role of genetics in predicting cross-societal cultural values variation, and potentially speak to the need for and importance of incorporating both nature and nurture in theories of cultural values variation across societies.
Similar content being viewed by others


Introduction
Perhaps resulting from a revolution in contemporary understanding of genetic variability, interest in research linking genes to social and psychological phenomena, termed “sociogenetics”, has burgeoned in the last decade1. Via this paradigm, evidence indicates that there are unique and meaningful cross-societal effects of genes on complex traits such as social stratification, social trust, intelligence test performance, educational attainment, voter turnout, happiness, and cognitive achievement2,3,4,5,6. Scholars have suggested that a deeper understanding of the interplay between genes and culture is essential for a holistic understanding of psychological processes7. Yet, very little is known about the role of genetics in cross-cultural differences, defined as differences in cultural values orientations between societies1,8,9,10. To the extent that prevailing theories of cultural values focus only on environmental factors e.g.,8,9,10, a full accounting that considers the potential role of genetic differences between individuals from divergent populations is lacking. Indeed, the promise of finding genetic explanations for cultural values variation across societies is important. Insight into the genetic bases of said variation may help explain persistent differences in cultural values orientations between equally modernized and prosperous countries such as the US and Japan8,9,10. A holistic understanding involving both environmental and genetic influences of cross-cultural values variation between societies will enable scientists, students, and business and governmental decision makers to build more accurate templates for understanding social behavior and tackling social problems that arise resultant of differences between nations.
Consistent heritability estimates from twin and whole-genome studies suggest there is a reliable, albeit small, effect of genes on complex psychological traits such as personality and cultural values differences between individuals11. Comparing average population differences in genetic variability at the country or societal level has been advocated as an approach that helps address these small effects of genes at the individual level5,12. Accordingly, an emerging stream of research studying culture and genetics, via the paradigm of culture-gene co-evolution theory, has focused on associations between cultural values (e.g., individualism, tightness, power distance) and country-level variability in 5-HTTLPR12,13,14,15, a polymorphism in SLC6A4 (“Solute carrier family 6 member 4”), the gene containing information for production of the SERT protein that is responsible for transportation of serotonin from the synaptic cleft to its neuron1. However, results have been conflicting, possibly as a result of two important problems in this literature on the sociogenetics of culture.
First, a singular focus on only one polymorphism, 5-HTTLPR, has been a conceptual limitation restricting the accuracy of potential conclusions. The SLC6A4 gene is a complex system consisting of various polymorphic regions that each serve specific functions. Focusing on only one aspect of this system may lead to incorrect conclusions regarding associations between genes and culture. Crucially, there are two polymorphisms in SLC6A4 that play complementary roles in regulating serotonin receptivity, combining to either reduce or enhance serotonin uptake: 5-HTTLPR and STin216,17. Second, extant studies lack model completeness and sample representativeness. Former studies on the sociogenetics of culture have only sporadically included environmental control variables, without consideration of the full range of environmental influences18. This is problematic because failing to account for environmental covariates leaves open the question whether documented associations between 5-HTTLPR and cultural values12,13,14,15 are robust. Additionally, said associations have also mostly not included populations from the “global South”, most notably sub-Saharan Africa.
Hence, and as detailed below, we address the just-noted issues by (1) synthesizing culture-gene co-evolution theory with advances in medical science involving allelic variation in serotonin receptivity to better clarify underlying associations between genes and cultural values; (2) conducting the first study on the effects of both polymorphisms in SLC6A4, 5-HTTLPR and STin2, on cultural values; (3) including all environmental factors that have been identified as important antecedents of cultural values; (4) including good representation of countries in the global South via reference to genetic maps of Eurasian, African, and Amerindian populations. We do so by examining population level associations between the SLC6A4 polymorphisms and two cultural values for which measurement-valid and scientifically rich data are available for most countries—individualism (the extent to which the individual is prioritized over the group) and monumentalism (the extent to which personality and behavior are viewed as interdependent and static as opposed to independent and malleable19,20).
In more detail, culture-gene coevolution theory suggests that environmental pressures giving rise to genetic adaptations also give rise to corresponding sociocultural adaptations21. Natural selection works to enhance the survival and reproductive fitness of individuals possessing genes linked to behavior patterns that yield success in a given social and physical environment22. Notably, genes affecting brain function are likely to influence the adoption and formation of cultural norms; conversely, culture may also shape the expression and selection of genes21. Consequently, cross-cultural variation between societies may be a function of genetic variation, with different patterns of gene-brain interactions giving rise to cultural differences between populations7,23. These genetic variation patterns between populations are also unlikely to be a result of chance (i.e., genetic drift) because similar patterns of gene-brain interactions have been found worldwide in ecologically threatening regions that are geographically isolated, involving SLC6A47,15.
Two Variable Number of Tandem Repeat (VNTR) polymorphisms in SLC6A4, 5-HTTLPR and STin2, work together to increase SERT protein transportation from the SLC6A4 gene, combining to reduce or enhance serotonin uptake17,24,25,26. 5-HTTLPR has two allelic variants including the S (“short”) and L (“long”) alleles. Individuals with a homozygotic S-allele have lower serotonin receptivity than individuals with a homozygotic L-allele. The L-allele causes more efficient SLC6A4 transcription than the S-allele. SERT translation is decreased among homozygotic S-allele carriers, vice versa for homozygotic L-allele carriers27,28,29. STin2, responsible for SERT protein translation also30, is in intron 2 of the SLC6A4 gene and consists of 9, 10, or 12 repeat units, (STin2.9, STin2.10, and STin2.12). STin2.9 is quite rare, but almost everyone has either STin2.10 (henceforth, “10/10”) or STin2.12 (henceforth, “12/12”). Like the S-allele, 12/12 increases SLC6A4 transcription and decreases SERT translation; like the L-allele, 10/10 leads to more efficient SLC6A4 transcription and increased SERT translation17,31. Overall, both polymorphisms have been linked to similar outcomes including mental health problems, depression, suicide, and nicotine dependence29,30,32,33,34. Although underlying biological mechanisms for these findings are not fully understood yet35,36, because 12/12 and the S-allele both decrease SERT protein translation, it may generally be surmised that they should have similar effects on social outcomes and vice versa for 10/10 and the L-allele, which increase SERT translation17,31. Hence, potential associations between cultural values and STin2 may be informed by the extant research on 5-HTTLPR.
The S-allele is associated with enhancement of a wide range of cognitive functions, notably improved decision-making through better probabilistic and temporal discounting37. It promotes hypervigilance, leading to more anxiety, heightened responses to emotional stimuli, and more attention to negative information but also more environmental monitoring37,38,39. Consequently, individuals with homozygotic S-alleles are more risk-averse and more loss-averse than those with homozygotic L-alleles40,41. Thus, because national culture is closely related to aggregated personality traits5, it is reasonable to expect that societies typified by a greater prevalence of S-allele (or likewise, 12/12) carriers will also be more risk-averse, favoring cultural norms that embody more social harmony and social control13 (e.g., collectivism, monumentalism). Indeed, environmental threats that select for less serotonin uptake in the population also have been evidenced to covary with “survivalist” cultural tendencies dictating greater social cohesion such as the creation of strong hierarchies, tightly knit societies, and the prevalence of strong norms14,15,42. Therefore, we predict that SLC6A4 polymorphisms associated with reduced serotonin uptake (S-allele and 12/12) will be positively associated with monumentalism (more social control) but negatively associated with individualism (less social harmony); vice versa for 10/10, which increases serotonin uptake16,17. Going further, because culture-gene co-evolution theory maintains that associations between allelic and cultural variability are independent of environmental factors21,22,23, we predict that 5-HTTLPR and STin2 will explain unique variance in individualism and monumentalism beyond the effects of all essential environmental factors.
Summarily, we conduct the most comprehensive study to date on the confluence of cultural values and genetics, providing insight into specific patterns of genetically driven cross-cultural differences for a set of 28 countries representing 57% of the world population. First, we review the medical literature and aggregate data from 75 studies representing 28,726 individuals to determine the prevalence of the STin2 polymorphism across said countries. Ours is the first tabulation of STin2 across countries, providing a wealth of information on serotonin receptivity across populations that can be used by scholars of sociogenetics. Further, we use genetic mapping of the populations spanning the various continents to provide estimates of the STin2 polymorphism in another 27 countries that also have data available for S-allele prevalence. We thereby greatly expand the country-level database available for scholars to study SLC6A4, the serotonin transporter gene, while also ensuring that any estimates we provide are based on genetically comparable populations. Most importantly, because environmental factors exert powerful influences on cultural values and may in fact be responsible for both cultural and allelic variation42,43,44,45,46, we draw upon conceptual advancements in ecological psychology18 to statistically model 14 environmental factors including: geographical latitude, axial orientation, climatic demands, rainfall steadiness, historical/contemporary prevalence of disease, social diversity including ethnic/racial, linguistic, and religious diversity, population density, urbanization, wealth differences both between and within countries, and social conflict. This set subsumes all environmental covariates included in former studies of 5-HTTLPR12,13,14,15 and no other factors beyond these have been identified as important antecedents of cultural values8. Thus, we control for most confounds to statistically test the supposition that there is a genetic explanation for cross-cultural values variation between societies. To be clear, we do not make any inferences about a causal link between genes and culture among individuals. Rather, we model associations using a correlation and regression approach, based on overall population-wide data, focused only on predicting statistical variation in cultural values orientations.
Results
STin2 distributions across countries
Stin2 prevalence for all countries in our study are displayed in Fig. 1 (for 12/12) and 2 (for 10/10). A detailed tabulation of these countries and the primary studies they are based on is provided in Tables S1 and S2 (see "Supplementary Information"). 28 countries in our study had primary data on STin2—these are listed in the top half of Table S1. Another 27 countries were estimated based on genetic maps of the populations within these countries, via reference to the scientific literature delineating genetic overlaps between and within Eurasian, African, and Amerindian populations47,48,49,50,51 (see “Method” section for more details). Countries for which STin2 data were estimated are listed in the bottom half of Table S1.

STin2.12 (12/12) allelic prevalence worldwide. Numbers shown represent estimated population percentages. Note: Countries with primary data available are shaded in red and countries with estimated data available are shaded in green. Map generated with CorelDRAW (Ver. 2019), available at https://www.coreldraw.com/en/.
An examination of Figs. 1 and 2 reveals an interesting pattern. The five countries or regions with the highest prevalence of the 12/12 allele all have predominantly East Asian populations, including China (80%), Japan (79%), Singapore (85%), South Korea (84%), and Taiwan (80%). In contrast, the five countries with the lowest prevalence of the 12/12 allele all have mostly European populations including Australia (31%), Canada (31%), Germany (26%), Sweden (29%), and the UK (35%). Overall, Asian and African countries or regions have the highest prevalence of the 12/12 allele and European countries have the lowest. When considering 10/10 allelic prevalence, the five lowest countries/regions are East Asian, including China (1%), Japan (1%), South Korea (1%), and Taiwan (2%), and Southeast Asian (Vietnam; 2%); the five highest countries mostly also have predominantly European populations, including Australia (19%), Brazil (23%), Croatia (16%), France (18%), and Russia (20%). The 12/12 allele is much more prevalent in the population than the 10/10 allele.

STin2.10 (10/10) allelic prevalence worldwide. Numbers shown represent estimated population percentages. Note: Countries with primary data available are shaded in blue and countries with estimated data available are shaded in green. Map generated with CorelDRAW (Ver. 2019), available at https://www.coreldraw.com/en/.
Correlation analyses
Correlation analyses were conducted in SPSS (Ver. 28). A detailed tabulation of the means, standard deviations, and intercorrelations between all study variables is provided in Table S3 (see "Supplementary Information"). As expected, the two STin2 alleles, 12/12 and 10/10, are very strongly and negatively related to each other (r = − 0.75, p < 0.001). As expected also, given that they are in different regions of SLC6A4, neither of the STin2 alleles are strongly correlated with S-allele prevalence (for 12/12: r = 0.26, n. s.; for 10/10: r = − 0.05, n. s.). To the extent that all correlations between the genetic variables are in logically expected directions and are consistent with prior research, we may have some confidence that the currently presented genetic data have measurement validity.
As shown in Table S3, directions of correlations between the SLC6A4 polymorphisms and the two cultural values orientations are generally in theoretically expected directions. S-allele and 12/12 allelic prevalence are positively correlated with monumentalism (a cultural value associated with more social control); this association is significant only for the S-allele (for 12/12: r = 0.07, n. s.; for S-allele: r = 0.58, p < 0.01). However, only 12/12 is, as expected, negatively correlated with individualism (a cultural value associated with less social harmony); S-allele prevalence is positively correlated with individualism, although this latter correlation is not statistically significant (for 12/12: r = − 0.52, p < 0.01; for S-allele: r = 0.22, n. s.). On the other hand, the 10/10 allele is, as expected, positively correlated with individualism and negatively correlated with monumentalism; the correlation is significant for individualism but not monumentalism (for individualism: r = 0.26, p = 0.05; for monumentalism: r = − 0.08, n. s.). Overall, findings provide some support for the notion that 12/12 and the S-allele are positively associated with cultural values that promote social harmony or social control (monumentalism) and negatively with those that do not (individualism), vice versa for 10/10.
Regression analyses
Hierarchical regression analyses were conducted in SPSS (Ver. 28) to test the effects of the SLC6A4 polymorphisms on cultural values after accounting for the effects of environmental factors, with country as the unit of analysis. This analysis helps shed light on concerns surrounding the validity of previously evidenced correlations between SLC6A4 polymorphisms and cultural values orientations12,13,14,15. The regression analysis was conducted in three steps. All control variables were included in Step 1—this step indicates the amount of cultural values variance explained by factors other than the SLC6A4 polymorphisms and serves as a baseline to compare the unique contributions of the polymorphisms in explaining values variation between societies. The S-allele percentage was included in Step 2—this step indicates the unique contribution of the S-allele to values variation between societies after accounting for the control variables. Finally, 12/12 and 10/10 allelic percentages were included in Step 3—this step indicates the unique contribution of STin2 to cultural values variation between societies after accounting for the control variables and S-allele prevalence. Japan and New Zealand were excluded from the regression analyses because they lacked data on axial orientation. Findings are thus based on the remaining 53 countries.
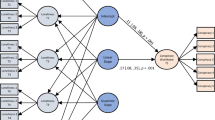
A summary snapshot displaying the main findings of interest is displayed in Fig. 3. Detailed results for the hierarchical regression analyses are provided in Table 1. The table lists the effects of the individual predictors (standardized regression coefficients) on individualism and monumentalism, the amount of variance explained in each step on individualism and monumentalism (Adjusted R2), the incremental variance explained by the SLC6A4 polymorphisms on individualism and monumentalism in each of Steps 2 and 3 (Δ R2), and a test statistic (F) indicating whether said variance explained for individualism or monumentalism is statistically significant. Statistically significant effects are bolded.

Percentage variance explained in cultural values by the SLC6A4 polymorphisms and other factors.
As shown in Table 1, the effects of pathogen prevalence and GDP PPP on individualism are statistically significant. The environmental control variables collectively explain 79.2% of the cultural values variation in individualism. None of the SLC6A4 polymorphisms (S-allele, 12/12, and 10/10) are significantly associated with individualism after controlling for the environmental factors in Step 1. These polymorphisms do not explain additional variation in individualism beyond the effects of the environmental factors, and collectively account for close to 0% of the variance in this cultural values dimension. Turning to monumentalism, the effects of latitude, rainfall steadiness, and the Gini coefficient are statistically significant. The environmental control variables collectively explain 58.5% of the cultural values variation in monumentalism. Effects of the S-allele and the 12/12 allele on monumentalism are statistically significant. The S-allele explains an additional 17.4% of the variation in monumentalism beyond the effects of all environmental factors, and the 12/12 allele explains an additional 6.2% of the variance beyond this. Collectively, the SLC6A4 polymorphisms explain 23.6% additional variation in monumentalism beyond the effects of the environmental factors, indicating a strong and potentially important cultural antecedent.
Robustness tests
There are several limitations with our statistical model. Four countries including France, Italy, Singapore, and Sweden, and one region, Taiwan, had only small sample sizes for STin2 frequency estimates in the population, raising the possibility that small sample sizes for population genetic estimates may have confounded study results. Four countries including Burkina Faso, El Salvador, Tanzania, and Zambia relied on estimated data on both genetic variables and cultural dimensions, thereby raising the possibility that data dependencies may have confounded study results. Twenty-seven countries (all countries listed in the bottom half of Table S1) did not include primary data on STin2, raising the possibility that study results may have been confounded by our use of genetic maps to “guess” STin2 estimates on genetically similar countries where no such primary data were in fact available. We conducted robustness tests to address all these potential limitations with our statistical model. Results are provided in Tables S4–S6 (see "Supplementary Information").
As shown in Table S4, study findings did not change after excluding countries with small sample sizes for STin2 frequency estimates in the population. In line with results in Table 1, neither S-allele, 12/12, nor 10/10 prevalence significantly predicted individualism after controlling for the environmental factors. As before, both S-allele and 12/12 prevalence significantly predicted monumentalism beyond the effects of environmental factors.
As shown in Table S5, study findings changed slightly after excluding countries with estimated data on both genetic variables and cultural dimensions. After controlling for environmental factors, the effect of S-allele prevalence on individualism was marginally significant; as before, neither 12/12 nor 10/10 were significant. As before too, both S-allele and 12/12 prevalence significantly predicted monumentalism beyond the effects of environmental factors.
As shown in Table S6, study findings became even stronger after excluding countries with estimated data on STin2. In line with results in Table 1, neither S-allele, 12/12, nor 10/10 prevalence significantly predicted individualism after controlling for the environmental factors. However, in addition to the S-allele and 12/12, 10/10 also had a statistically significant effect on monumentalism; taken together, the effect of the SLC6A4 polymorphisms on monumentalism dwarfed the effects of all environmental factors, accounting for almost half of the variation (48.9%) in this cultural value.
Overall, results of the robustness tests converge toward results reported in Table 1, ruling out the possibility that small sample sizes, data dependencies, or using genetic maps to estimate data for countries without primary data available for STin2 may have confounded the study results. These results demonstrate that there is a robust and practically meaningful genetic explanation for cultural values variance between societies as a function of naturally evolving differences in serotonin uptake across geographically distinct human populations.
Multicollinearity analyses
The small variable-observation ratio may have led to biased regression coefficients resulting from multicollinearity between study variables. Therefore, multicollinearity analyses were conducted to rule out this potential confound. Across all models (Table 1, Tables S4–S6), analyses indicated no multicollinearity between study variables, with all variance inflation factors below 10, ruling out the small variable-observation ratio as a study confound.
Discussion
Synthesizing culture-gene co-evolution theory with advances in medical science involving allelic variation in serotonin receptivity, we conducted the most comprehensive study to date on cross-societal cultural values variation and genetics. We posited and found some support for the notion that there is a genetic explanation for a portion of the cultural values variation across societies. Notably, 12/12 and S-allele prevalence, both responsible for decreased serotonin uptake, were found to explain almost a quarter of the variation in monumentalism but no variance in individualism. These results held even after controlling for all key environmental factors and subjecting the data to stringent robustness tests.
The fact that the SLC6A4 polymorphisms did not predict individualism converges with previous research indicating that S-allele prevalence does not predict individualism-collectivism after other influences are accounted for12,52. Our results also replicate previous research indicating S-allele prevalence to be an important predictor of flexibility-monumentalism12. However, we go further and show that in addition to the S-allele, 12/12 is also perhaps an equally important predictor. Taken together, our findings suggest that the SLC6A4 polymorphisms may be very important influences of cultural values that concern social control, such as flexibility-monumentalism, but of no consequence for cultural values concerning social harmony such as individualism-collectivism. Indeed, the fact that former studies have found robust associations between S-allele prevalence and other cultural values related to social control such as social hierarchy and social constraint14,15 buttresses this notion. Future research is needed to deeper investigate potential associations between serotonin receptivity and different types of cultural values e.g.,8,9,10.
Coupled with evidence that there are vast differences across countries on frequencies of the STin2 alleles, ranging from 26% in Germany to 85% in Singapore for 12/12, our findings speak to the potential importance of incorporating both nature and nurture in the study of culture. Given evidence associating values and genetic variation, theories on antecedents of cultural values e.g.,8,9 may now be able to add another focal predictor: population allelic variation in SLC6A4, the serotonin transporter gene. Scholars studying cross-societal cultural values differences across countries may easily include this factor into their predictive models by referring to the STin2 data we provide; countries not represented in our data may be estimated using the genetic mapping method we have introduced.
A genetic component to cultural variability across societies helps to explain the persistent cross-cultural differences that have been found between equally modernized and wealthy countries such the US and Japan and suggests a potential roadblock toward global cultural convergence. Illustratively, cross-cultural scholars have found evidence for global convergence toward individualism and related values across the world’s countries, and speculate that at the present rate of change, cross-societal cultural values differences may altogether disappear by 205053. Our findings, however, argue for the opposite position, indicating that the world’s societies will yet be quite divergent in their cultural values orientations given stubborn genetic differences between populations. Future research on the potential role of genetic variation in cultural values change across societies is needed.
Interestingly also, although the broad pattern suggests continental differences in the genetic divide, with East/Southeast Asia anchoring the high-end of 12/12 prevalence and Europe the low end, there are differences even within continents. Focusing on the continent with the most country-specific data available, Europe, there are large differences even between neighboring countries, with 12/12 allelic percentages ranging from 25% in Germany to 35% in neighboring France and 40% in the Netherlands and Belgium. Indeed, a 10–15% difference is quite substantial given how geographically close and historically intertwined these countries are; by way of comparison, the East Asian countries are all within ≈ 5% of each other. Could these genetic differences help explain persistent cultural values variation that has been found across Europe?9,10 Future research examining associations between cultural values variation and serotonin receptivity is needed to best answer this question.
Limitations
Because we used population-wide metrics for environmental antecedents, polymorphisms, and cultural values, our study is strictly correlational only—a causal link between serotonin receptivity and values cannot be presently inferred. Ideally, individual-level data on the studied polymorphisms, coupled with population-wide metrics on environmental antecedents and cultural values, ought to be paired in a hierarchical linear model to make inferences regarding said causal link. We are unable to do this because the polymorphic data do not exist at the individual- but only at the sample-level (see Table S2). Hence, we maintain only that the SLC6A4 gene is an important factor in predicting cultural values variance (i.e., a statistical, not causal, explanation). On that note, we caution the reader to not derive a deterministic understanding of culture based upon a reading of our paper. We take the position that both nature and nurture are important antecedents of culture, and we hope that more research on this issue will spur forward.
Likewise, lacking individual-level data on specific polymorphisms, we cannot rule out the possibility that sociocultural differences between neighboring countries such as France and Germany may simply be resultant of differential ancestral origins, as opposed to genetic variation in serotonin receptivity. For instance, Bell Beaker culture was predominant in Germany but not so much in France54, leaving open the possibility that these countries evolved different cultural values simply because they were populated by people with different cultural practices to begin with. Future research disentangling the effects of ancestral origin and allelic variation in serotonin receptivity is needed to resolve this nature-nurture question.
Finally, we acknowledge that assigning a certain percentage in allelic variation for a given country is an imperfect method of estimating the actual allelic variation in countries with highly diverse populations such as Singapore, New Zealand, the UK, and the USA. Unfortunately, the primary studies from the medical science literature (see Table S2) by and large did not include detailed sample demographic breakdowns by race/ethnicity, precluding us from creating more refined estimates of cross-societal allelic variation in STin2. Nevertheless, the genetic maps we referenced47,48,49,50,51, particularly for Eurasian populations, suggest that many ethnic groups share common genetic origins (e.g., Finns, Russians, and Turks fall in the same genetic group49), thereby somewhat alleviating this concern. Moreover, it is unlikely that estimation sensitivity could rule out the effects we found considering the sheer size of said effects, whereby the SLC6A4 polymorphisms accounted for almost a quarter of the variation in monumentalism but close to zero in individualism (see Table 1). The robustness of our results hence lends some measure of confidence to our conclusions.
Method
STin2 literature review
PubMed was searched using the term “STin2” in February and March 2020; the search was then updated in March 2021. One hundred and fifty-three potential studies were identified for inclusion. To be included, studies must have been published in English and provided relevant sample statistics, including (a) the number of participants with STin2.12/STin2.12 (“12/12”), (b) the number of participants with STin2.10/STin2.10 (“10/10”), and (c) the total number of participants. Seventy-five studies met our inclusion criteria. The 75 studies are listed in Table S2 (see "Supplementary Information"). Information on country of origin, sample type, total sample size, and the total number of sample participants carrying both 12/12 and 10/10 STin2 alleles are also listed. As shown, primary STin2 data were available for 28 countries (28,726 individuals), representing parametric estimates for ≈ 57% of the world population.
Calculation of country-specific percentages of the STin2 alleles
Across all countries, some samples were clinical (defined as samples whose members have specific ailments or comorbidities such as depression or cancer), whereas others included only healthy adults. To confirm genetic equivalencies between clinical and non-clinical samples, we compared the number of individuals with 12/12 and 10/10 alleles present in each type of sample. Independent-samples t-tests indicated no statistically significant differences between clinical and non-clinical samples on either the 12/12 (t (115) = − 1.146, p = 0.254) or 10/10 alleles (t (115) = 0.367, p = 0.715). These non-significant results are consistent with the extant medical literature indicating similar frequencies of the STin2 alleles between clinical and non-clinical samples27,55. Therefore, both clinical and non-clinical samples were summed to calculate STin2 percentages across specific countries.
For each country, the number of participants with 12/12 and/or 10/10 STin2 alleles was divided by the total number of sample participants to calculate a country-specific percentage. If a country had data from more than one study, all the participants were taken into consideration, with percentages calculated using the total participants carrying each allele, respectively, and divided by the total number of participants included in that country. For the United Kingdom, data were available separately for both England and Scotland. Given large discrepancies in population between these two parts of the UK, 12/12 and 10/10 percentages were multiplied by 0.90 for England, and 0.10 for Scotland, and then summed. The final calculated percentages for all 28 countries with primary data available are listed in the top half of Table S1 (see "Supplementary Information").
STin2 percentage estimates via genetic mapping
Following prior research12, we estimated an additional 27 countries based on the available primary data. To ensure that all our estimates are valid, we utilized population genetic mapping of the continents to build genetically accurate estimates of STin2 allelic prevalence in each country. Because hypotheses involve comparing the relative effects of 5-HTTLPR and STin2, we limit estimates to 27 additional countries that also have information on S-allele prevalence12. STin2 (12/12 and 10/10) prevalence estimation procedures for these 27 countries are detailed below, by continent. Estimates are provided in the bottom half of Table S1 (see "Supplementary Information").
Africa
A genetic map of Africa is provided by Tishkoff et al.47. Using these data, estimates for Burkina Faso, Ghana, Nigeria, Zambia, and Zimbabwe are made based on South Africa; estimates for Rwanda and Tanzania are made based on Uganda.
Americas
A genetic map of the Americas is provided by Salas et al.48. Using these data, estimates for Chile, Argentina, and Uruguay are based on Brazil; El Salvador and Guatemala are based on Colombia; Peru is estimated based on the average of Brazil and Colombia.
Asia
Genetic clusters for the various Eurasian populations are provided by the GenomeAsia100K Consortium49. Using these data, Azerbaijan, Israel, and Turkey are estimated based on the average of Iran and Russia.
Europe
A genetic map of Europe is available at: https://www.eupedia.com/europe/european_y-dna_haplogroups_by_region.shtml. The map is based on converging genetic data on the Y-chromosome prevalence from fourteen peer-reviewed, scientific sources e.g.,50,51. Based on these data, and coupled also with genetic cluster data from the GenomeAsia100K Consortium49, STin2 estimates for Europe are as follows: Austria is estimated based on the average of Hungary and the Czech Republic; Estonia, Finland, and Ukraine are estimated based on Russia; Denmark and Norway are estimated based on Sweden; Poland is estimated based on the average of Czech Republic, Hungary, and Russia; Slovenia is estimated based on Croatia; Switzerland is estimated based on the average of Germany and France; Ireland is estimated based on the UK.
Exceptions
One exception to the use of these genetic maps is New Zealand. The related genetic map49 detailed the DNA analysis of the local Maori population. However, the native Maori are a minority in New Zealand; 70% of the country’s population is of European (mostly British) descent. Therefore, we estimated New Zealand based on the UK.
S-allele percentages
S-allele percentages for most countries are provided by Minkov et al.12, excepting Croatia and Portugal. The S-allele percentage for Croatia was calculated by averaging data from available studies56,57,58. Data for Portugal are provided by Ferreira et al.59.
Environmental control variables
Conceptual advancements in ecological psychology18 have introduced a typology of four spatial and temporal explanations that are theoretically linked to variation in human behavior across countries. First, geographical explanations based on human biogeographical theory suggest that peoples’ habits are dependent on their habitat locations60. Second, ecological explanations based on climatic theories of human behavior e.g.,61 suggest that climatic harshness shapes peoples’ behaviors as a function of the number of resources they may have to use in coping with these threats coupled with threat severity. Third, evolutionary explanations drawing upon life-history theory e.g.,62 suggest that dangerous and unpredictable environments promote fast life strategies (e.g., risk aversion), whereas stable and safe environments promote more self-control. Fourth, contemporary explanations draw upon compensatory-control theories of human behavior e.g.,63, suggesting that threat adaptation is a function of the amount of material resources that people have. Thus, we control for variables subsuming all four sets of environmental factors.
Geographical controls
First, we control for two geographical factors including latitude and axial orientation. Latitudinal position is defined as the geographical position of a country on a bipolar axis ranging from the South Pole through the equator to the North Pole; scores are available for all countries from van de Vliert and van Lange18. Latitude has been found to robustly predict cultural values such as societal individualism, dwarfing associations between values and the concomitant longitudinal position18. Axial orientation, defined as the extent to which countries spread out in an east–west vs. a north–south fashion, drives cultural diversity because flora and fauna are more easily domesticated across longitudes than latitudes, allowing for better cultural spread64. It has been found to be strongly associated with cultural diversity, even after considering other environmental factors; scores for all non-archipelago countries are provided by Laitin et al.44. Data were unavailable on this index for two countries in our study, Japan and New Zealand, because they are archipelagos.
Ecological controls
Second, we control for two ecological factors including climatic demands and rainfall steadiness. Climatic demands are defined as a country’s absolute deviation from ambient temperature (22 °C) in both heat and cold in both the hottest and coldest months. It represents the bipolarity of a climate, with larger seasonal variations having greater impacts on human behavior18. Climatic demands have been found to robustly predict cultural values such as societal collectivism45,61. Rainfall steadiness is defined as the minimum monthly precipitation divided by the maximum monthly precipitation18. Scores for all countries are available from van de Vliert and van Lange18. It represents year-round water accessibility, a vital resource for human populations, with more steady rainfall leading to reduced threats of both deluge and drought and is theorized to be an effective proxy for climate-induced calamities, such as food scarcity, flooding, soil erosion, and crop ruination18.
Evolutionary controls
Third, we control for four evolutionary factors including pathogen prevalence (historical and contemporary), social diversity (including racial/ethnic, linguistic, and religious diversity), modernization (including urbanization and population density), and social conflict.
Pathogen prevalence is calculated as the average rate of nine pathogens that capture all biological detriments to human fitness resulting in disease including leishmanias, trypanosomes, malaria, schistosomes, filariae, leprosy, dengue, typhus, and tuberculosis. Both historical and contemporary epidemiological indicators of these variables were calculated because they have both been found to be strongly associated with cultural values such as societal individualism42. Scores for all countries are available from Fincher et al.42. Data on pathogen prevalence were not available for a few countries in our analysis and were thus estimated based on neighboring countries: Azerbaijan was estimated based on Iran; Rwanda and Uganda were estimated based on Tanzania; Ukraine was estimated based on Russia. Because both historical and contemporary pathogen prevalence were very strongly and positively correlated in our data (r = 0.81), an “average pathogen prevalence” measure was calculated for inclusion in analyses.
Social differences in language, race, and religion have a long history of being associated with culture e.g.,65,66,67. We thus control for social diversity, defined as the extent to which a country is fractionalized along ethnic/linguistic/religious lines, and using country scores provided for all countries from Luiz68. Because these three types of diversity tend to coincide (e.g., more ethnicities imply more languages spoken), we calculated an averaged “social diversity” score based on the three indices for inclusion in all analyses (α = 0.70).
Likewise, modernization theory has long attributed cultural differences to the shift from agricultural and relatively isolated forms of living to urbanized and crowded forms of living69,70. We thus control for this factor using World Bank data on the urbanization rate within countries, defined as the percentage of people living in cities71, and population density, defined as the logged number of inhabitants per square kilometer in each country72. Both these variables have been found to predict cultural values such as collectivism and tightness43,45,73.
Finally, social conflict has long been identified as a source of threat mandating the formation of socially cohesive societies8,74. More specifically, a country’s history of territorial threat is defined as the number of conflict incidences that have occurred within its territorial borders8. Illustratively, the bombing of Pearl Harbor and the Russian Revolution were conflict incidences occurring within US and Russian territorial borders, respectively, whereas the Iraq wars and the invasion of Afghanistan were extra-territorial conflict incidences involving these countries. Data on historical conflict incidences from 1918 to 2016 are available for all countries from the International Crisis Behavior Database75,76.
Contemporary controls
Fourth, we control for two contemporary factors, including material resources available for threat adaptation both between- and within-countries (i.e., wealth). Wealth differences have been found to strongly predict cultural values such as individualism-collectivism45,46,61. Wealth differences between countries is defined as the gross domestic product per capita (GDP PPP); data were available for all countries in our study from the World Bank77. Wealth differences within countries is defined as the Gini coefficient; data were available for all countries in our study from the World Bank78.
Dimensions of societal culture
Individualism and monumentalism country scores were available for 50 out of 55 countries in our study20. The remaining five countries were estimated based on cultural values clusters across world regions10, as follows: Burkina Faso and Zambia were based on South Africa; El Salvador was based on Colombia; Tanzania and Uganda were based on Rwanda.
Data availability
The data used in this study are available from the following sources: STin2: Please see Table S2 below for the 75 primary studies that these data were extracted from. S-allele (Croatia): Hranilovic et al.56. Noskova et al.57. Božina et al.58. S-allele (Portugal): Ferreira et al.59. S-allele (All Other Countries): Minkov et al.12. Latitudinal position, climatic demands, rainfall steadiness, historical pathogen prevalence, and contemporary pathogen prevalence: Van de Vliert and van Lange18. Axial orientation: Laitin et al.44. Social diversity including racial/ethnic, religious, and linguistic diversity: Luiz68. Urbanization rate: World Bank71. Population density: World Bank72. GDP PPP: World Bank77. Gini coefficient: World Bank78. Number of territorial conflicts: Brecher and Wilkenfeld75. Brecher et al.76. Cultural dimensions: Minkov and Kaasa20.
References
Gelfand, M. J., Aycan, Z., Erez, M. & Leung, K. Cross-cultural industrial organizational psychology and organizational behavior: A hundred-year journey. J. Appl. Psychol. 102, 514–529. https://doi.org/10.1037/apl0000186 (2017).
Abdellaoui, A. et al. Genetic correlates of social stratification in Great Britain. Nat. Hum. Behav. 3, 1332–1342. https://doi.org/10.1038/s41562-019-0757-5 (2019).
Aaroe, L. et al. Genetic predictors of educational attainment and intelligence test performance predict voter turnout. Nat. Hum. Behav. 5, 281–291. https://doi.org/10.1038/s41562-020-00952-2 (2021).
Kong, D. T. A gene-environment interaction model of social trust: The 5-HTTLPR S-allele prevalence as a moderator for the democracy-trust linkage. Pers. Individ. Differ. 87, 278–281. https://doi.org/10.1016/j.paid.2015.08.028 (2018).
Minkov, M. & Bond, M. H. A genetic component to national differences in happiness. J. Happiness Stud. 18, 321–340. https://doi.org/10.1007/s10902-015-9712-y (2017).
Minkov, M., Welzel, C. & Bond, M. H. The impact of genes, geography, and educational opportunities on national cognitive achievement. Learn. Individ. Differ. 47, 236–243. https://doi.org/10.1016/j.lindif.2016.03.004 (2016).
Kwon, H. & Sasaki, J. Y. Gene-Culture Interactions: Toward an Exploratory Framework (Cambridge University Press, 2019).
Gelfand, M. J. Rule Makers, Rule Breakers: How Culture Wires Our Minds, Shapes Our Nations, And Drives Our Differences (Robinson, 2018).
House, R. J. et al. (eds) Culture, Leadership, and Organizations: The GLOBE study of 62 Societies (Sage, 2004).
Hofstede, G., Hofstede, G. J. & Minkov, M. Cultures and Organizations: Software of the Mind 3rd edn. (McGraw-Hill, Uk, 2010).
Sallis, H., Smith, G. D. & Munafo, M. R. Genetics of biologically based psychological differences. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 373, 20170162. https://doi.org/10.1098/rstb.2017.0162 (2018).
Minkov, M., Blagoev, V. & Bond, M. H. Improving research in the emerging field of cross-cultural sociogenetics: The case of serotonin. J. Cross. Cult. Psychol. 46, 336–354. https://doi.org/10.1177/0022022114563612 (2015).
Chiao, J. Y. & Blizinsky, K. D. Culture-gene coevolution of individualism-collectivism and the serotonin transporter gene. Proc. Biol. Sci. 277, 529–537. https://doi.org/10.1098/rspb.2009.1650 (2010).
Fischer, R. Gene-environment interactions are associated with endorsement of social hierarchy values and beliefs across cultures. J. Cross. Cult. Psychol. 44, 1107–1112. https://doi.org/10.1177/0022022112471896 (2013).
Mrazek, A. J., Chiao, J. Y., Blizinsky, K. D., Lun, J. & Gelfand, M. J. The role of culture-gene coevolution in morality judgment: Examining the interplay between tightness-looseness and allelic variation of the serotonin transporter gene. Cult. Brain. 1, 100–117. https://doi.org/10.1007/s40167-013-0009-x (2013).
Blazevic, S. et al. Epigenetic adaptation of the placental serotonin transporter gene (SLC6A4) to gestational diabetes mellitus. PLoS ONE 12, e0179934. https://doi.org/10.1371/journal.pone.0179934 (2017).
Hemmings, S. et al. Appetitive and reactive aggression are differentially associated with the STin2 genetic variant in the serotonin transporter gene. Sci. Rep. 8(6714), 2–3. https://doi.org/10.1038/s41598-018-25066-8 (2018).
Van de Vliert, E. & van Lange, P. A. M. Latitudinal psychology: An ecological perspective on creativity, happiness, aggression, and beyond. Perspect. Psychol. Sci. 14, 860–884. https://doi.org/10.1177/1745691619858067 (2019).
Minkov, M. A revision of Hofstede’s model of national culture: Old evidence and new data from 56 countries. Cross Cult. Strateg. Manag. 25, 231–256. https://doi.org/10.1108/CCSM-03-2017-0033 (2018).
Minkov, M. & Kaasa, A. Do dimensions of culture exist objectively? A validation of the revised Minkov-Hofstede model of culture with World Values Survey items and scores from 102 countries. J. Int. Manag. 28, 100971. https://doi.org/10.1016/j.intman.2022.100971 (2022).
Walsh, A. & Yun, I. Evoked culture and evoked nature: The promise of gene-culture co-evolution theory for sociology. Front. Sociol. 1, 8. https://doi.org/10.3389/fsoc.2016.00008 (2016).
Putnam, S. P. & Gartstein, M. A. Aggregate temperament scores from multiple countries: Associations with aggregate personality traits, cultural dimensions, and allelic frequency. J. Pers. 67, 157–170. https://doi.org/10.1016/j.jrp.2016.07.009 (2017).
Chiao, J. Y. & Ambady, N. Cultural neuroscience: Parsing universality and diversity across levels of analysis. In Handbook of Cultural Psychology (eds Kitayama, S. & Cohen, D.) (The Guilford Press, 2007).
Ali, F. R. et al. Combinatorial interaction between two human serotonin transporter gene variable number tandem repeats and their regulation by CTCF. J. Neurochem. 112, 296–306. https://doi.org/10.1111/j.1471-4159.2009.06453.x (2010).
Kalungi, A. et al. Association between serotonin transporter gene polymorphisms and increased suicidal risk among HIV positive patients in Uganda. BMC Genet. 18(71), 2–7. https://doi.org/10.1186/s12863-017-0538-y (2017).
Vijayan, N. N. et al. Evidence of association of serotonin transporter gene polymorphisms with schizophrenia in a South Indian population. J. Hum. Genet. 54, 538–542. https://doi.org/10.1038/jhg.2009.76 (2009).
Joshi, G., Pradhan, S. & Mittal, B. No direct association of serotonin transporter (STin2 VNTR) and receptor (HT 102T>C) gene variants in genetic susceptibility to migraine. Dis. Markers. 29, 223–229. https://doi.org/10.3233/DMA-2010-0752 (2010).
Kohen, R. et al. The serotonin transporter polymorphism rs25531 is associated with irritable bowel syndrome. Dig. Dis. Sci. 54, 2663–2670. https://doi.org/10.1007/s10620-008-0666-3 (2009).
Koks, G. et al. Genetic interaction between two VNTRs in the SLC6A4 gene regulates nicotine dependence in Vietnamese men. Front. Pharmacol. 9(1398), 1–8. https://doi.org/10.3389/fphar.2018.01398 (2018).
Kohen, R. et al. Association of serotonin transporter gene polymorphisms with poststroke depression. Arch. Gen. Psychiatry. 65, 1296–1302. https://doi.org/10.1001/archpsyc.65.11.1296 (2008).
Saiz, P. A. et al. Association between the A-1438G polymorphism of the serotonin 2A receptor gene and nonimpulsive suicide attempts. Psychiatr. Genet. 18, 213–218. https://doi.org/10.1097/YPG.0b013e3283050ada (2008).
Li, D. & He, L. Meta-analysis supports association between serotonin transporter (5-HTT) and suicidal behavior. Mol. Psychiatry 12, 47–54. https://doi.org/10.1038/sj.mp.4001890 (2007).
Bah, J. et al. Serotonin transporter gene polymorphisms: Effect on serotonin transporter availability in the brain of suicide attempters. Psychiatry Res. 162, 221–229. https://doi.org/10.1016/j.pscychresns.2007.07.004 (2008).
Mackenzie, A. & Quinn, J. A serotonin transporter gene intron 2 polymorphic region, correlated with affective disorders, has allele-dependent differential enhancer-like properties in the mouse embryo. Proc. Natl. Acad. Sci. 96, 15251–15255. https://doi.org/10.1073/pnas.96.26.15251 (1999).
Mitchell, C. et al. Role of mother’s genes and environment in postpartum depression. Proc. Natl. Acad. Sci. 108, 8189–8193. https://doi.org/10.1073/pnas.1014129108 (2011).
Saiz, P. A. et al. Association study between obsessive-compulsive disorder and serotonergic candidate genes. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 765–770. https://doi.org/10.1016/j.pnpbp.2007.12.005 (2008).
Jolmsberg, J. R. & Lesch, K.-P. Looking on the bright side of serotonin transporter gene variation. Biol. Psychiatry 69, 513–519. https://doi.org/10.1016/j.biopsych.2010.09.024 (2011).
Munafo, M. R., Durrant, C., Lewis, G. & Flint, J. Gene x environment interactions at the serotonin transporter locus. Biol. Psychiatry 65, 211–219. https://doi.org/10.1016/j.biopsych.2008.06.009 (2009).
Lee, P. H., Bakermans-Kranenburg, M. J., van-Ijzendoom, M. H. & Yair, B.-H. Variations in the promoter region of the serotonin transporter gene and biased attention for emotional information: A meta-analysis. Biol. Psychiatry. 71, 373–379. https://doi.org/10.1016/j.biopsych.2011.10.030 (2012).
He, Q. et al. Serotonin transporter gene-linked polymorphic region (5-HTTLPR) influences decision making under ambiguity and risk in a large Chinese sample. Neuropharmacology 59, 6518–6526. https://doi.org/10.1016/j.neuropharm.2010.07.008 (2010).
Heitland, I. et al. Genetic polymorphisms of the dopamine and serotonin systems modulate the neurophysiological response to feedback and risk taking. Cogn. Affect. Behav. Neurosci. 12, 678–691. https://doi.org/10.3758/s13415-012-0108-8 (2012).
Fincher, C. L., Thornhill, R., Murray, D. R. & Schaller, M. Pathogen prevalence predicts human cross-cultural variability in individualism-collectivism. Proc. R. Soc. Lond. B. Biol. Sci. 275, 1279–1285. https://doi.org/10.1098/rspb.2008.0094 (2008).
Harrington, J. R. & Gelfand, M. J. Tightness-looseness across the 50 United States. Proc. Natl. Acad. Sci. 111, 7990–7995. https://doi.org/10.1073/pnas.1317937111 (2014).
Laitin, D. D., Moortgat, J. & Robinson, A. L. Geographic axes and the persistence of cultural diversity. Proc. Natl. Acad. Sci. 109, 10263–10268. https://doi.org/10.1073/pnas.1205338109 (2012).
Marcus, J., Kahraman, F., Su, S. & Fritzsche, B. A. Capturing intranational cultural variation in international business research: Microsocietal differences in collectivism across Turkey. J. World Bus. 54, 101020. https://doi.org/10.1016/j.jwb.2019.101020 (2019).
Santos, H. C., Varnum, M. E. W. & Grossman, I. Global increases in individualism. Psychol. Sci. 28, 1228–1239. https://doi.org/10.1177/0956797617700622 (2017).
Tishkoff, S. A. et al. The genetic structure and history of Africans and African Americans. Science 324, 1035–1044. https://doi.org/10.1126/science.1172257 (2009).
Salas, A. et al. Mitochondrial echoes of first settlement and genetic continuity in El Salvador. PLoS ONE 4, e6882. https://doi.org/10.1371/journal.pone.0006882 (2009).
GenomeAsia100K Consortium. The GenomeAsia100K Project enables genetic discoveries across Asia. Nature 576, 106–111. https://doi.org/10.1038/s41586-019-1793-z (2019).
King, R. J. et al. The coming of the Greeks to Provence and Corsica: Y-chromosome models of archaic Greek colonization of the western Mediterranean. BMC Evol. Biol. 11(69), 1–8. https://doi.org/10.1186/1471-2148-11-69 (2011).
Underhill, P. A. et al. Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a. Eur. J. Hum. Genet. 18, 479–484. https://doi.org/10.1038/ejhg.2009.194 (2010).
Eisenberg, D. T. & Hayes, M. G. Testing the null hypothesis: Comments on “Culture-gene coevolution of individualism-collectivism and the serotonin transporter gene”. Proc. R. Soc. Lond. B. Biol. Sci. 278, 329–332. https://doi.org/10.1098/rspb.2010.0714 (2011).
Taras, V., Steel, P. & Kirkman, B. L. Improving national cultural indices using a longitudinal meta-analysis of Hofstede’s dimensions. J. World Bus. 47, 329–341. https://doi.org/10.1016/j.jwb.2011.05.001 (2012).
Olalde, I. et al. The Beaker phenomenon and the genomic transformation of Northwest Europe. Nature 555, 190–196. https://doi.org/10.1038/nature25738 (2018).
Wendland, J. R., Kruse, M. R., Cromer, K. R. & Murphy, D. L. A large case-control study of common functional SLC6A4 and BDNF variants in obsessive-compulsive disorder. Neuropsychopharmacology 32, 2543–2551. https://doi.org/10.1038/sj.npp.1301394 (2007).
Hranilovic, D. et al. Serotonin transporter gene promoter (5-HTTLPR) and intron 2 (VNTR) polymorphisms in Croatian suicide victims. Biol. Psychiatry 54, 884–889. https://doi.org/10.1016/s0006-3223(03)00179-3 (2003).
Noskova, T. et al. Ethnic differences in the serotonin transporter polymorphism (5-HTTLPR) in several European populations. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 1735–1739. https://doi.org/10.1016/j.pnpbp.2008.07.012 (2008).
Božina, N. et al. Suicide ideators and attempters with schizophrenia—the role of 5-HTTLPR, rs25531, and 5-HTT VNTR Intron 2 variants. J. Psychiatr. Res. 46, 767–773. https://doi.org/10.1016/j.jpsychires.2012.03.008 (2012).
Ferreira, J. T., Levy, P. Q., Marinho, C. R., Bicho, M. P., & Mascarenhas, M. R. G. Association of serotonin transporter gene polymorphism 5HTTVNTR with osteoporosis. Acta Reumatol. Port. 36, 14–19. https://pubmed.ncbi.nlm.nih.gov/21483275/ (2011).
Harcourt, A. H. Humankind: How Biology and Geography Shape Human Diversity (Pegasus, 2015).
Fischer, R. & van de Vliert, E. Does climate undermine subjective well-being? A 58-nation study. Pers. Soc. Psychol. Bull. 37, 1031–1041. https://doi.org/10.1177/0146167211407075 (2011).
Kaplan, H. S., & Gangestad, S. W. Life history theory and evolutionary psychology. In Handbook of Eevolutionary Psychology (ed. Buss, D. M.) 68–95 (Wiley, 2005).
Landau, M. J., Kay, A. C. & Whitson, J. A. Compensatory control and the appeal of a structured world. Psychol. Bull. 141, 694–722. https://doi.org/10.1037/a0038703 (2015).
Diamond, J. Guns, Germs, and Steel: The Fates of Human Societies (W. W. Norton and Company, 1997).
Chomsky, N. Language and Mind (Harcourt, 1968).
Greenfield, P. M. Sociodemographic differences within countries produce variable cultural values. J. Cross. Cult. Psychol. 45, 37–41. https://doi.org/10.1177/0022022113513402 (2014).
Huntington, S. P. The Clash of Civilizations and the Remaking of World Order (Simon & Schuster, 1997).
Luiz, J. M. The impact of ethno-linguistic fractionalization on cultural measures: Dynamics, endogeneity, and modernization. J. Int. Bus. 46, 1080–1098. https://doi.org/10.1057/jibs.2015.6 (2015).
Harari, Y. N. Sapiens: A Brief History of Humankind. (Harvill Secker, 2011/2014).
Tonnies, F. Community and Society. (Michigan State University Press, 1887/1957).
World Bank. Urbanization rate. https://data.worldbank.org/indicator/SP.URB.TOTL.IN.ZS (2018a).
World Bank. Population density. https://data.worldbank.org/indicator/EN.POP.DNST (2018b).
Gelfand, M. J. et al. Differences between tight and loose cultures: A 33-nation study. Science 332, 1100–1104. https://doi.org/10.1126/science.1197754 (2011).
Sumner, W. G. Folkways: A Study of the Sociological Importance of Usages, Manners, Customs, Mores, and Morals (Ginn and Company, 1906).
Brecher, M. & Wilkenfeld, J. A Study of Crisis (University of Michigan Press, 2000).
Brecher, M., Wilkenfeld, J., Beardsley, K., James, P., & Quinn, D. International crisis behavior data codebook, Version 13. http://sites.duke.edu/icbdata/data-collections/ (2020).
World Bank. GDP, PPP. https://data.worldbank.org/indicator/NY.GDP.MKTP.PP.CD (2018c).
World Bank. GINI index. https://data.worldbank.org/indicator/SI.POV.GINI (2018d).
Acknowledgements
We would like to thank the five anonymous reviewers for their highly constructive, detailed, and insightful feedback on previous iterations of our manuscript. Their collective comments greatly elevated manuscript quality and potential study contribution.
Author information
Authors and Affiliations
Contributions
Both authors contributed equally to the manuscript and were equally involved in all stages of the research.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Marcus, J., Cetin, E. Genetic predictors of cultural values variation between societies. Sci Rep 13, 7986 (2023). https://doi.org/10.1038/s41598-023-34845-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-34845-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
