
Abstract
To analyse the pathogenic genes and mutations of a family with hereditary deafness. We recruited a three-generation family with NSHL. A detailed medical history inquiry and related examinations were performed. Next-generation sequencing (NGS) was used to confirm the gene mutation in the proband, and Sanger sequencing was used for verification. The effect of the WFS1 mutation on the function and structure of the wolframin protein was predicted by multiple computational software. From the Gene Expression Omnibus (GEO) database, we obtained GSE40585 dataset and performed enrichment analyses. The family clinically manifested as autosomal dominant NSHL. A novel WFS1 c.2421C>G (p.Ser807Arg) mutation was identified in four affected individuals in the pedigree . The p.Ser807Arg mutation is a highly conserved residue and causes an increase in protein stability. It had an important influence on not only amino acid size, charge and hydrophobicity but also protein intermolecular hydrogen bonding and spatial structure. There were differentially expressed genes (DEGs) in GSE40585 dataset. Enrichment analysis revealed that DEGs mainly functioned in amino acid metabolism, signal transduction and dephosphorylation. We reported a novel mutation c.2421C>G (p.Ser807Arg in WFS1. This study expands the mutation spectrum of WFS1.
Similar content being viewed by others
Introduction
As one of the most common sensorineural defects, hereditary hearing loss is divided into syndromic hearing loss (SHL) and nonsyndromic hearing loss (NSHL), accounting for approximately 30% and 70% of HHL, respectively1,2. It is estimated that approximately 80% of NSHL cases are autosomal recessive, 15% are autosomal dominant, and less than 5% are X-linked/mitochondrial3. hereditary hearing loss has an extremely high degree of heterogeneity in terms of causative gene mutations and inheritance modes. To date, 51 causative genes and over 70 loci responsible for autosomal dominant nonsyndromic hearing loss (ADNSHL) have been confirmed (http://hereditaryhearingloss.org/; The Hereditary Hearing Loss Homepage). Nevertheless, there are still many mutations and genes involved in the onset and development of hearing loss unidentified. With the development of next-generation sequencing (NGS)4, characterized by high throughput and low cost, it is possible to screen all deafness-associated genes in a high-throughput manner. In our study, a Chinese ADNSHL family was recruited, and targeted deafness multigene sequencing and computational tools were used to explore the molecular basis of the mutation.
Materials and methods
Subjects
Proband with ADNSHL was recruited from the Second Hospital of Hebei Medical University. In addition to the proband, six additional members (I:2, II:2, II:3, II:5, II:7, III:3) of the family were recruited from the hospital, including two affected and three unaffected. The Ethics Committee of the Second Hospital of Hebei Medical University approved the study. We confirmed that all experiments were performed in accordance with relevant guidelines and regulations. We obtained informed consent from the subjects.
Clinical information and examination
We obtained the medical histories of the subjects through a questionnaire on the following aspects: onset age, progression, tinnitus, noise exposure, use of ototoxic drugs, pregnancy/labour process, medication, use of hearing aids, and other relevant clinical manifestations. A detailed physical examination was performed in all patients and ruled out the possibility of syndromic hearing loss. The grades of hearing impairment were measured by pure tone audiometry (PTA). The average hearing threshold refers to the average values at 500, 1000, 2000, and 4000 Hz. According to the world report on hearing 2021 (https://www.who.int/publications/i/item/world-report-on-hearing), hearing levels were classified as normal (< 20 dB), mild (20 to < 35 dB), moderate (35 to < 50 dB), moderately severe (50 to < 65 dB), severe (65 to < 80 dB), profound (80 to < 95 dB), and complete (≥ 95 dB)5.
Targeted sequencing and mutation analysis
For mutation analysis, peripheral blood (approximately 5–10 ml) was obtained from the subjects, and genomic DNA was extracted with a DNA extraction Kit (Qiagen, Shanghai, China) and stored at − 20 °C until use. We used the Nanodrop 2000 (Thermal Fisher Scientific, USA) to quantify the DNA and the GenCap® deafness gene system capture kit (MyGenostics Inc, Beijing, China) to capture the amplified DNA. The gene panel for hearing loss disease is summarized in Table S1. The capture experiment was conducted according to the manufacturer’s protocol. Targeted NGS of 415 known deafness genes was performed using the MyGenotics gene enrichment system and the HiSeq2000 sequencer (Illumina, USA). The parameter BWA of Sentieon software (https://www.sentieon.com/) can map the clean reads to the UCSC hg19 human reference genome. ANNOVAR software (http://annovar.openbioinformatics.org/en/latest/) annotated variants further and was associated with multiple databases, such as 1000 Genomes (http://www.1000genomes.org/), ESP6500, dbSNP, EXAC, Inhouse (MyGenostics), and HGMD. Computational programs, such as SIFT (http://sift.jcvi.org/), PolyPhen_2 (http://genetics.bwh.harvard.edu/pph2/), Mutation Taster (http://www.mutationtaster.org/), and GERP+, were used to predict the pathogenicity of the candidate variants. We used the guidelines of the American College of Medical Genetics and Genomics (ACMG) to interpret the data. Beijing MyGenostics Company performed the sequencing analysis.
Mutation detection by Sanger sequencing
Validation primers were designed for the detected WFS1 gene mutation sites, and Sanger sequencing was performed on subjects whose samples had been collected. Forward (5′-ACATCAAGAAGTTCGACCGC-3′) and reverse (5′- GCAATCTACACATGGTCGCA-3′) primers were designed using NCBI Primer-BLAST and synthesized by MyGenostics Company (Beijing, China). Sanger sequencing was used to verify the cosegregation of the pathogenic mutations and the hearing phenotype within the family.
Mutation analysis by computational servers
MUpro (http://mupro.proteomics.ics.uci.edu), I-Mutant2.0 (https://folding.biofold.org/i-mutant/i-mutant2.0.html) and iStable (http://predictor.nchu.edu.tw/istable/indexSeq.php) were used to predict protein stability changes upon single point mutation. HOPE (http://www.cmbi.umcn.nl/hope) can analyse the impact of a given mutation on the protein structure. TMHMM Server 2.0 (https://services.healthtech.dtu.dk/service.php?TMHMM-2.0) is an online tool to predict transmembrane structures in proteins. Accessible Surface Area and Accessibility Calculation for Protein (ver. 1.2) online server can calculate solvent-accessible surface areas (SASA) of 890 amino acids of wolframin and S807 mutant protein. We built a three-dimensional (3D) model of the wolframin protein by Alphafold2 and presented the wild-type and mutant protein structures by PyMOL programs.
Screening key genes and signalling pathways by bioinformatics analysis
GSE40585 gene expression profile microarray data were downloaded from GEO database (http://www.ncbi.nlm.nih.gov/geo/). The dataset was uploaded by Sulev Kõks in 2012 and included two types of HEK transfected cells: wild type and WFS1 silenced type. Quantitative RT-PCR and western blot confirmed clear silencing of WFS1 gene expression after 48 h. Therefore, we selected 4 samples transfected with blank plasmid for 48 h as the control group and 8 samples transfected with WFS1 siRNA for 48 h as the experimental group. Using R software, we identified genes which were differentially expressed genes (DEGs) between groups. STRING (https://string-db.org/) was used to predict the protein–protein interaction (PPI) network of DEGs. Cytoscape software (v3.8.0) is used to visualize the network diagram. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were conducted on differential genes to investigate their potential biological functions. The Gene Set Enrichment Analysis (GSEA) (v.4.2.3) was performed for all genes in study samples.
Results
Clinical features
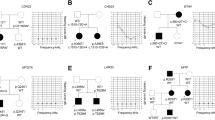
There was a three-generation family with ADNSHL in China, including five affected patients and seven unaffected individuals. Seven people in the family participated in the research (4 affected and 3 unaffected). The affected member III:1 did not participate in the study due to personal reasons. The autosomal dominant inheritance pattern was shown in the family (Fig. 1a). None of the subjects in the study showed clinical signs in any other organs except hearing impairment. There was no vestibular or visual involvement. None of the members had ever used cochlear implants or hearing aids. Audiograms of the subjects of the family showed that their hearing impairments were bilateral, symmetric, sensorineural, postlingual, early onset and progressive (Table 1).

Combined figure. (a) Pedigreed diagrams of the family with autosomal dominant hearing loss (symbols with line, deceased). The affected subjects are denoted in black. The proband is indicated by an arrow. (b–g) Audiograms of the family subjects (red: right ear; blue: left ear). (b–e) Audiogram showed bilateral sensorineural hearing impairment of affected subjects. (f–h) tonal audiometric curves of unaffected family members.
A 24-year-old man is the proband, whose history of sensorineural hearing loss is 21 years. His disease is characterized by low frequency hearing loss, which gradually increased with age (Fig. 1b).The subject I:2 had an onset age in the third decade of life, which was the latest in this family, and her hearing impairment had slowly progressed to profound deafness with advancing age (Fig. 1c). The onset age of the subject II:2 was in the first decade of life, and his audiogram showed moderate hearing loss at all frequency (Fig. 1d). We could not get the exact age of onset of the subjects, except the proband, because of the negligence of their hearing loss and the lack of audiologic data at early ages. The subject II:5 had similar audiograms with the proband, but his left ear had a higher hearing threshold (Fig. 1e). The audiogram of the subjects II:3, II:7, III:3 were normal (Fig. 1f,g,h).
Mutation detection and analysis
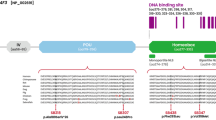
A novel WFS1 mutation in the Chinese ADNSHL family was identified in the study. The average sequencing depth for the targeted regions was 917.17-fold. There was 98.85% and 98.36% coverage of the targeted exons for 10X and 20X, respectively. In targeted sequencing, the c.2421C>G (p.Ser807Arg) mutation in the WFS1 gene in exon 8 was identified in the proband (Fig. 2a). At amino acid 807, serine was replaced by arginine in the mutation. All patients carried this heterozygous mutation, but the normal family members did not carry the mutation (Fig. 2b). We verified the mutation by Sanger sequencing. This mutation cosegregated with hearing loss in four affected patients in pedigree (I:2; II:2; II:5; III:2) (Table 1). The prediction results using REVEL, SIFT, PolyPhen_2, Mutation Taster and GERP were shown in Table 2. All of the tools predicted that WFS1 c.2421C > G (p.Ser807Arg) was a damaging mutation. According to the ACMG guidelines 6, c.2421C>G (p.Ser807Arg) is classified as a likely pathogenic variant (PS1 + PM1 + PM2_Supporting + PP3). The mutation is classified as PS1, meaning that mutations of the same amino acid changes have been reported in database, but the nucleotide changes are different (Table 3). The p.Ser807Arg mutation localized in the intracytoplasmic hydrophilic C-terminal domain of the wolframin protein. The alignment of WFS1 from various species is shown in Fig. 2c. It was shown that the serine residue at position 807 was highly conserved among different species, meaning it may play an important role in proper protein function.

Mutation detection and conservation analysis. (a) Sequencing results show that the heterozygote c.2421C > G mutation was found in the proband (III:2). (b) unaffected family member (II:7). (c) Protein alignment shows conservation of the S807 residue of WFS1 across 9 species.
Computational analysis of the mutation effect
We further explored the changes in the secondary or tertiary structure of the protein that may be caused by the p.Ser807Arg mutation. According to the results of the ConSurf analysis, p.Ser807Arg had a conservation score of 9, meaning that it was highly conserved in evolution (Table 4). The effect of the p.Ser807Arg mutation on protein stability was predicted by MUpro, I-Mutant 2.0 and iStable software. It was shown that the p.Ser807Arg mutation resulted in increased protein stability. Compared with the wild type, the p.Ser807Arg mutation increased the size of the amino acid, changed the charge, and decreased the hydrophobicity. These changes may change the intramolecular interactions to influence the function of the wolframin protein.
We used TMHMM to investigate the effect of the p.Ser807Arg mutation on the transmembrane region of the wolframin protein. As shown in Fig. 3a, it was suggested that the p.Ser807Arg mutation resulted in no changes in the structure of the wolframin transmembrane region. According to SASA analysis, the residual fluctuations between the wild-type and mutant p.Ser807Arg proteins were similar (Fig. 3b). Compared with the wild type, the proportion of mutant p.Ser807Arg protein core amino acids was decreased, which suggested a detrimental influence on protein function (Fig. 3c).

Combined figure. (a) The nine transmembrane regions in mutant S807R protein are shown by TMHMM server. (b) The bottom panels describe per-residue SASA of wolframin protein and mutant S807R protein. (c) The distribution about the SASA of all 890 amino acids in wild and mutant S807R protein.
The three-dimensional structures of wolframin and mutant p.Ser807Arg protein were predicted by the Robetta server. To compare the three-dimensional structure before and after mutation, the p.Ser807Arg mutant protein was superimposed on the wild-type protein by PyMOL (Fig. 4a). The substitution regions were highlighted in the protein model (Fig. 4b). Comparing the qualitative electrostatic representation between the wild-type and mutant p.Ser807Arg proteins, the p.Ser807Arg mutation changed the charge of the amino acid at this site from neutral to positive (Fig. 4c,d). In the wild type, Ser807 had a hydrogen bond with Glu809 and Phe810 (Fig. 4e). In the mutant, the original hydrogen bond distances between Arg807 and Phe810 became shorter, while the hydrogen bond between Arg807 and Glu809 disappeared (Fig. 4f). In addition, a hydrogen bond between Arg807 and Asp339 was added. The stability and intramolecular interactions of the wolframin protein may be affected by the above changes in hydrogen bonds, resulting in diseases.

Mutation-induced structural changes in WFS1. (a) It shows superimposed view of wolframin protein in wild and mutant state. (b) Predicted structures depict the changes of mutant wolframin protein with the amino acid change S807R. Red and brown structures indicate differences between wild and mutant type. (c) A qualitative electrostatic representation of wolframin protein generated by PyMOL. Protein contact potentials can be represented by displaying virtual (false) red/blue charged smooth surfaces on wolframin protein. (d) A qualitative electrostatic representation of mutant S807R protein generated by PyMOL. The black circle indicates the position of amino acid 807. (e–f) The change of hydrogen bond between amino acids before and after mutation. The proteins are shown as cartoon. Amino acids at the mutated site 807 are highlighted in yellow and the interacting amino acids are highlighted in blue (red dotted line: the hydrogen bond).
Hub genes screening and enrichment analysis
For the purpose of exploring the potential mechanism of deafness caused by WFS1 gene mutation, we have screened key genes and signaling pathways by integrated bioinformatics analysis. A heatmap of DEGs is shown in Fig. 5a, while a PPI network is shown in Fig. 5b. Jun proto-oncogene (JUN), KIT proto-oncogene (KIT), cyclin dependent kinase inhibitor 1B(CDKN1B), calreticulin (CALR), interferon beta 1(IFNB1), sapiens secreted phosphoprotein 1(SPP1) and Toll Like Receptor 2 (TLR2) were indicated as hub genes. According to GO analysis, the changes of biological process (BP) of DESs were mainly concentrated in detection of chemical stimulus involved in sensory perception and dephosphorylation (Fig. 6a–c). KEGG pathway analysis showed that olfactory transduction, complement and coagulation cascades and other types of o-glycan biosynthesis were statistically significant (Fig. 6d). Comparing gene expression differences between groups is the purpose of GO and KEGG enrichment analysis. GSEA analyses the whole gene dataset, so genes with little differential expression but a great deal of significance will not be excluded from GSEA analysis (Fig. 6e). JAK-STAT signalling pathway, amino acid metabolism, and taste transduction are the most statistically significant biological processes.

Heat map and PPI network. (a) Heat map of the 12 samples. Each column represents a sample, and each row represents one of the differentially expressed genes. (b) PPI network of GSE40585.

Enrichment analysis between wild type and WFS1 silenced type. (a) Biological process of DEGs. (b) Cellular component of DEGs. (c) Molecular function of DEGs. (d) KEGG pathway of DEGs. (e) Significant KEGG enrichment plot from GSEA analysis.
Discussion
The WFS1 gene, also named Wolfram syndrome type 1, encodes the wolframin protein. It is a membrane glycoprotein localized in the endoplasmic reticulum (ER). Wolframin has been shown to be highly expressed in the pancreas, brain, heart, and muscle but expressed differentially in inner ear cells7. It plays a critical role not only in the regulation of insulin production and secretion from pancreatic β-cells but also in the regulation of ER stress response and cellular calcium homeostasis8. Wolframin can negatively regulate the ER stress signalling network to avoid the accumulation of misfolded and unfolded proteins9. Wolframin can modulate the filling state of the ER Ca2+ store to participate in the regulation of cellular Ca2+ homeostasis10. Once a variant occurs in WFS1, ER stress is strongly induced, and endolymphatic ion composition and homeostasis are disrupted, which leads to deafness.
There are more than 490 reported mutations of the WFS1 gene in the Human Gene Mutation Database. Mutations in the WFS1 gene are predominantly responsible for nonsyndromic hearing loss (DFNA6), Wolfram syndrome and Wolfram-like syndrome 11,12. In recent years, some studies have also shown that WFS1 is linked to type 2 diabetes, autosomal dominant optic atrophy (ADOA), and psychiatric problems13,14,15. WFS1-associated Wolfram syndrome type 1 (WS1) is defined as a rare autosomal neurodegenerative recessive progressive disease and is also known as DIDMOAD, characterized by diabetes insipidus (DI), childhood onset diabetes mellitus (DM), optic atrophy (OA), and deafness (D)16. Wolfram-like syndrome is an uncommon autosomal dominant disease that contains variable clinical manifestations, including sensorineural hearing loss, optic atrophy and/or diabetes mellitus17. The WFS1-associated nonsyndromic hearing loss has two types: autosomal dominant hearing loss, characterized by low-frequency decline, and autosomal recessive hearing loss, characterized by congenital profound sensorineural deafness.
Low-frequency sensorineural hearing loss (LFSNHL) is a rare form of hearing loss that affects only 2000 Hz and below frequencies and generally deteriorates over time without progressing to severe deafness18. Only two associated genes have been reported to date: DIAPH1 and WFS119,20. DIAPH1 is linked to the DFNA1 locus on chromosome 5q3121. It is characterized by rapid progression, eventually developing into profound deafness, from low to involve all higher frequencies. DFNA54 is the third locus reported to be associated with LFSNHL. It was mapped in a large Swiss family harbouring an unknown gene that results in low frequency hearing impairment when affected22. In 2001, mutations in the WFS1 gene were confirmed to be one of the causes of LFSNHL, which was the causative gene at the DFNA6 and DFNA14 loci23. In the same year, Young et al. came to the same conclusion through linkage analysis of a 6-generation LFSNHL family in Canada, thus suggesting that the causative genes of DFNA6, DFNA14 and DFNA38 are the same—the WFS1 gene24. Generally, ADNSHL patients with WFS1 mutations mostly have progressive, bilateral sensorineural hearing loss. In this study, the grandmother and uncle of the proband (I:2 and II:2) besides low frequency sensorineural hearing impairment, also show impairment at higher frequencies. It may be related to ageing in the grandmother and to working in a noisy environment for a long time in the uncle.
WFS1-accociated hearing loss can be caused by in-frame insertions or deletions, multiple protein terminating mutations, missense mutations and so on25,26,27. We summarized all types of mutations in the WFS1 gene that have been identified thus far in NSHL from the Human Gene Mutation Database (Tables S1–S2)28,29,30,31. In fact, the most common mutations associated with LFNSHL are missense mutations located in exon 8. In this study, we found that the WFS1 gene c.2421C>G (p.Ser807Arg) heterozygous mutation, located in exon 8, is the gene that causes deafness in this Chinese family. There was no report about the novel mutation in public databases. However, a mutation at the same residue (p.Ser807Arg) causing LFSNHL was first found in an English family32. Then, it was also reported in a Korean family33. The c.2419A>C (p.Ser807Arg) mutation was found in the Korean family, and the affected members had symmetrical bilateral hearing loss, which was mild to moderate in severity. The c.2421C>G (p.Ser807Arg) mutation in a Chinese family was first reported and identified in this study. The analysis of p.Ser807Arg mutations in the WFS1 gene is shown in Table 3. Missense mutations normally partially destroy the function of wolframin protein7. However, its exact physiological role in the cochlea remains unknown.
Generally, the majority of NSHL patients with mutations in WFS1 experienced hearing loss change from low frequency to full frequency, with no or slow progression except for presbycusis. Its onset time varies greatly from prelingual to the early 40 s. Most of them are heterozygous missense mutations, which belong to autosomal dominant inheritance. However, the homozygous variant c.972C>G of WFS1 in an Egyptian family was reported. The affected members presented with congenital bilateral severe to severe hearing loss, but they did not have other manifestations of Wolfram syndrome, presenting autosomal recessive inheritance 34. The most common hearing loss is the low frequency type, but some studies have proven that WFS1 mutation can also lead to middle, high or all frequency hearing loss; for example, Chinese researchers have reported that the mutation c.2389G>A (p. Asp797Asn) causes NSHL at all frequencies35. Some studies have shown that WFS1 mutations also play an important role in age-related hearing loss (ARHL). For example, a pathogenic mutation 1582T>C (p. Tyr528His) in WFS1 was identified in an ARHL Finnish family36. It is suggested that the wolframin protein could participate in the signalling pathway of the unfolded protein response (UPR)37, which is related to the pathogenesis of ARHL38.
To date, it has been reported that at least 71 different missense mutations in WFS1 are associated with NSHL, comprising 65 mutations in exon 8, four in exon 5, and one in exon 4 (Table S1). The mutations associated with NSHL were mainly concentrated in exon 8 and mostly clustered in the C-terminal domain of the wolframin protein (amino acids 652–890), suggesting that the region was very important for proper protein function of the inner ear. The mutations associated with DIDMOAD mostly do not cluster in any particular region of the protein and tend to be inactivating, which means that the cause of Wolfram syndrome is probably a loss of function of the wolframin protein. However, the causative mutations were found to be non-inactivating in NSHL, which suggests that NSHL results from specific mutations that partially destroy the function of intact proteins. The C-terminal region of wolframin is located on the cytoplasmic side of the ER membrane, adopting a folded confirmation. It can interact with the C-terminal domain of the ER-localized Na+/K+ ATPase beta1 subunit, which is important for subunit maturation39. To our knowledge, Na+/K+ ATPase deficiency is responsible for apoptosis and neural degenerative disease. If there are similar associations in the inner ear, amino acid substitutions may result in hearing loss in this way. It is also predicted that amino acid substitutions could change the secondary structure by disrupting helical chains, interfering with the ability of the C-terminal region to interact with other proteins in the ER lumen40. In conclusion, missense mutations in WFS1 may act in a dominant-negative way, causing hearing loss by disrupting protein‒protein interactions or protein phosphorylation.
Several signalling pathways may be upregulated or downregulated when the WFS1 gene is knocked down. The PPI network and hub genes analysis indicated that the knockdown of WFS1 had significant influence on carcinogenesis and development of cancer, calcium regulation, signal transduction, dephosphorylation. The JAK/STAT pathway is involved in lots of biological processes, such as immune regulation, cell division and apoptosis, and also plays a critical role on the embryonic development of the inner ear41. There are a number of amino acids that can act as neurotransmitters or neuromodulators, including alanine, aspartate, glutamate, and histidine42. WFS1 mutation may cause significant interference in amino acid metabolism pathway involved in neurotransmission. In addition, histidine may be related to metabolic syndrome and affect ion absorption43. All the analysis provided an indication for further study of the deafness mechanism caused by WFS1 mutation.
In conclusion, our study showed that the c.2421C>G p.Ser807Arg) mutation is a novel mutation of the WFS1 gene in the Chinese population. We recommended that WFS1 gene testing should be incorporated into routine examination for the genetic diagnosis of deafness, especially low-frequency hearing loss. Low-frequency hearing loss usually cannot be identified by routine methods of newborn hearing screening44, so it is necessary for LFNSHL families to conduct genetic counselling and genetic testing.
Data availability
The data supporting the results reported in the article can be found in the Supplementary Information files. The datasets generated and/or analysed during the current study are available in the NCBI database with the accession number SUB12158042 (https://www.ncbi.nlm.nih.gov/).
References
Kremer, H. Hereditary hearing loss; About the known and the unknown. Hear. Res. 376, 58–68. https://doi.org/10.1016/j.heares.2019.01.003 (2019).
Koffler, T., Ushakov, K. & Avraham, K. B. Genetics of hearing loss: Syndromic. Otolaryngol. Clin. N. Am. 48, 1041–1061. https://doi.org/10.1016/j.otc.2015.07.007 (2015).
Petersen, M. B. Non-syndromic autosomal-dominant deafness. Clin. Genet. 62, 1–13. https://doi.org/10.1034/j.1399-0004.2002.620101.x (2002).
Metzker, M. L. Sequencing technologies—The next generation. Nat. Rev. Genet. 11, 31–46. https://doi.org/10.1038/nrg2626 (2010).
Chadha, S., Kamenov, K. & Cieza, A. The world report on hearing, 2021. Bull. World Health Organ. 99, 242-242A. https://doi.org/10.2471/BLT.21.285643 (2021).
Oza, A. M. et al. Expert specification of the ACMG/AMP variant interpretation guidelines for genetic hearing loss. Hum. Mutat. 39, 1593–1613. https://doi.org/10.1002/humu.23630 (2018).
Cryns, K. et al. The WFS1 gene, responsible for low frequency sensorineural hearing loss and Wolfram syndrome, is expressed in a variety of inner ear cells. Histochem. Cell Biol. 119, 247–256. https://doi.org/10.1007/s00418-003-0495-6 (2003).
Lemaire, K. & Schuit, F. Integrating insulin secretion and ER stress in pancreatic beta-cells. Nat. Cell Biol. 14, 979–981. https://doi.org/10.1038/ncb2594 (2012).
Fonseca, S. G. et al. Wolfram syndrome 1 gene negatively regulates ER stress signaling in rodent and human cells. J. Clin. Investig. 120, 744–755. https://doi.org/10.1172/JCI39678 (2010).
Takei, D. et al. WFS1 protein modulates the free Ca(2+) concentration in the endoplasmic reticulum. FEBS Lett. 580, 5635–5640. https://doi.org/10.1016/j.febslet.2006.09.007 (2006).
Cryns, K. et al. Mutational spectrum of the WFS1 gene in Wolfram syndrome, nonsyndromic hearing impairment, diabetes mellitus, and psychiatric disease. Hum. Mutat. 22, 275–287. https://doi.org/10.1002/humu.10258 (2003).
Khanim, F., Kirk, J., Latif, F. & Barrett, T. G. WFS1/wolframin mutations, Wolfram syndrome, and associated diseases. Hum. Mutat. 17, 357–367. https://doi.org/10.1002/humu.1110 (2001).
Middle, F. et al. Bipolar disorder and variation at a common polymorphism (A1832G) within exon 8 of the Wolfram gene. Am. J. Med. Genet. 96, 154–157. https://doi.org/10.1002/(sici)1096-8628(20000403)96:2%3c154::aid-ajmg5%3e3.0.co;2-f (2000).
Hogewind, B. F. et al. Autosomal dominant optic neuropathy and sensorineual hearing loss associated with a novel mutation of WFS1. Mol. Vis. 16, 26–35 (2010).
Chistiakov, D. A. et al. A WFS1 haplotype consisting of the minor alleles of rs752854, rs10010131, and rs734312 shows a protective role against type 2 diabetes in Russian patients. Rev. Diabetic Stud. RDS 7, 285–292. https://doi.org/10.1900/RDS.2010.7.285 (2010).
Rigoli, L., Caruso, V., Salzano, G. & Lombardo, F. Wolfram syndrome 1: From genetics to therapy. Int. J. Environ. Res. Public Health 19, 3225. https://doi.org/10.3390/ijerph19063225 (2022).
Deng, H., Zhang, J., Zhu, F., Deng, X. & Yuan, L. Identification of the rare variant p.Val803Met of WFS1 gene as a cause of Wolfram-like syndrome in a Chinese family. Acta Diabetol. 57, 1399–1404. https://doi.org/10.1007/s00592-020-01572-y (2020).
Jung, A. R., Kim, M. G., Kim, S. S., Kim, S. H. & Yeo, S. G. Clinical characteristics and prognosis of low frequency sensorineural hearing loss without vertigo. Acta Otolaryngol. 136, 159–163. https://doi.org/10.3109/00016489.2015.1094824 (2016).
Pennings, R. J. et al. Progression of low-frequency sensorineural hearing loss (DFNA6/14-WFS1). Arch. Otolaryngol. Head Neck Surg. 129, 421–426. https://doi.org/10.1001/archotol.129.4.421 (2003).
Neuhaus, C. et al. Extension of the clinical and molecular phenotype of DIAPH1-associated autosomal dominant hearing loss (DFNA1). Clin. Genet. 91, 892–901. https://doi.org/10.1111/cge.12915 (2017).
Lynch, E. D. et al. Nonsyndromic deafness DFNA1 associated with mutation of a human homolog of the Drosophila gene diaphanous. Science 278, 1315–1318 (1997).
Gurtler, N. et al. DFNA54, a third locus for low-frequency hearing loss. J. Mol. Med. 82, 775–780. https://doi.org/10.1007/s00109-004-0597-1 (2004).
Bespalova, I. N. et al. Mutations in the Wolfram syndrome 1 gene (WFS1) are a common cause of low frequency sensorineural hearing loss. Hum. Mol. Genet. 10, 2501–2508. https://doi.org/10.1093/hmg/10.22.2501 (2001).
Young, T. L. et al. Non-syndromic progressive hearing loss DFNA38 is caused by heterozygous missense mutation in the Wolfram syndrome gene WFS1. Hum. Mol. Genet. 10, 2509–2514. https://doi.org/10.1093/hmg/10.22.2509 (2001).
Garcia-Garcia, G. et al. Improving the management of patients with hearing loss by the implementation of an NGS panel in clinical practice. Genes 11, 1467. https://doi.org/10.3390/genes11121467 (2020).
Gurtler, N., Rothlisberger, B., Ludin, K., Schlegel, C. & Lalwani, A. K. The application of next-generation sequencing for mutation detection in autosomal-dominant hereditary hearing impairment. Otol. Neurotol. 38, 900–903. https://doi.org/10.1097/MAO.0000000000001432 (2017).
Shearer, A. E. et al. Advancing genetic testing for deafness with genomic technology. J. Med. Genet. 50, 627–634. https://doi.org/10.1136/jmedgenet-2013-101749 (2013).
Walls, W. D. et al. A comparative analysis of genetic hearing loss phenotypes in European/American and Japanese populations. Hum. Genet. 139, 1315–1323. https://doi.org/10.1007/s00439-020-02174-y (2020).
Kobayashi, M. et al. WFS1 mutation screening in a large series of Japanese hearing loss patients: Massively parallel DNA sequencing-based analysis. PLoS ONE 13, e0193359. https://doi.org/10.1371/journal.pone.0193359 (2018).
Fukuoka, H., Kanda, Y., Ohta, S. & Usami, S. I. Mutations in the WFS1 gene are a frequent cause of autosomal dominant nonsyndromic low-frequency hearing loss in Japanese. J. Hum. Genet. 52, 510–515. https://doi.org/10.1007/s10038-007-0144-3 (2007).
Li, J. et al. Missense variant of endoplasmic reticulum region of WFS1 gene causes autosomal dominant hearing loss without syndromic phenotype. Biomed. Res. Int. 2021, 6624744. https://doi.org/10.1155/2021/6624744 (2021).
Cryns, K. et al. Mutations in the WFS1 gene that cause low-frequency sensorineural hearing loss are small non-inactivating mutations. Hum. Genet. 110, 389–394. https://doi.org/10.1007/s00439-002-0719-1 (2002).
Choi, H. J. et al. Whole-exome sequencing identified a missense mutation in WFS1 causing low-frequency hearing loss: A case report. BMC Med. Genet. 18, 151. https://doi.org/10.1186/s12881-017-0511-7 (2017).
Budde, B. S. et al. Comprehensive molecular analysis of 61 Egyptian families with hereditary nonsyndromic hearing loss. Clin. Genet. 98, 32–42. https://doi.org/10.1111/cge.13754 (2020).
Bai, X. et al. Identification of a novel missense mutation in the WFS1 gene as a cause of autosomal dominant nonsyndromic sensorineural hearing loss in all-frequencies. Am. J. Med. Genet. A 164A, 3052–3060. https://doi.org/10.1002/ajmg.a.36760 (2014).
Kytovuori, L., Hannula, S., Maki-Torkko, E., Sorri, M. & Majamaa, K. A nonsynonymous mutation in the WFS1 gene in a Finnish family with age-related hearing impairment. Hear. Res. 355, 97–101. https://doi.org/10.1016/j.heares.2017.09.013 (2017).
Fonseca, S. G. et al. WFS1 is a novel component of the unfolded protein response and maintains homeostasis of the endoplasmic reticulum in pancreatic beta-cells. J. Biol. Chem. 280, 39609–39615. https://doi.org/10.1074/jbc.M507426200 (2005).
Wang, W. et al. Impaired unfolded protein response in the degeneration of cochlea cells in a mouse model of age-related hearing loss. Exp. Gerontol. 70, 61–70. https://doi.org/10.1016/j.exger.2015.07.003 (2015).
Zatyka, M. et al. Sodium-potassium ATPase 1 subunit is a molecular partner of Wolframin, an endoplasmic reticulum protein involved in ER stress. Hum. Mol. Genet. 17, 190–200. https://doi.org/10.1093/hmg/ddm296 (2008).
Gurtler, N. et al. Two families with nonsyndromic low-frequency hearing loss harbor novel mutations in Wolfram syndrome gene 1. J. Mol. Med. 83, 553–560. https://doi.org/10.1007/s00109-005-0665-1 (2005).
Chen, Y. et al. Hedgehog signaling promotes the proliferation and subsequent hair cell formation of progenitor cells in the neonatal mouse cochlea. Front. Mol. Neurosci. 10, 426. https://doi.org/10.3389/fnmol.2017.00426 (2017).
Engskog, M. K. et al. β-N-Methylamino-l-alanine (BMAA) perturbs alanine, aspartate and glutamate metabolism pathways in human neuroblastoma cells as determined by metabolic profiling. Amino Acids 49, 905–919. https://doi.org/10.1007/s00726-017-2391-8 (2017).
Moro, J., Tome, D., Schmidely, P., Demersay, T. C. & Azzout-Marniche, D. Histidine: A systematic review on metabolism and physiological effects in human and different animal species. Nutrients 12, 1414. https://doi.org/10.3390/nu12051414 (2020).
Morton, C. C. & Nance, W. E. Newborn hearing screening—A silent revolution. N. Engl. J. Med. 354, 2151–2164. https://doi.org/10.1056/NEJMra050700 (2006).
Acknowledgements
We sincerely thank all the family members for their participation and cooperation in this study.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: Q.Z. Performed the experiments: J.Z. Data collection: S.Z. and Y.J. Data analysis: Y.L. and J.W.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhao, J., Zhang, S., Jiang, Y. et al. Mutation analysis of the WFS1 gene in a Chinese family with autosomal-dominant non-syndrome deafness. Sci Rep 12, 22180 (2022). https://doi.org/10.1038/s41598-022-26850-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-26850-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
