
Abstract
In the Mediterranean Sea, the Strait of Messina (MS) is a very peculiar area, connecting highly different regions and representing a privileged observatory for an early comprehension and assessment of ecosystems shifts. It is hypothesized that the outbreaks observed near the coast of many sites in the Mediterranean Sea may be the result of transport of permanent populations of P. noctiluca in pelagic waters to the coast, caused by specific hydrodynamic conditions. By both visual observations and numerical experiments our objective is twofold: (A) to help clarify whether the basin of the Aeolian Islands Archipelago (AIA), in the Southern Tyrrhenian Sea (STS), may be the site from which large populations of P. noctiluca are transported to the MS, and (B) to evaluate whether the upwelling turbulent system of the MS can be an energetic opportunity for this species. It should offer a rich habitat without jeopardizing the overall survival of the population, that is subject to stranding due to strong currents. Although very different, the two involved ecosystems (AIA and MS ) are complementary for the success of Pelagia noctiluca life cycle. Outputs obtained by coupling the 3D hydrodynamic model (SHYFEM) with a Lagrangian particle tracking model support the hypothesis of a connectivity between these two ecosystems, particularly in the first half of the year, indicating the coastal areas around the AIA as potential optimal source location for Pelagia larval stages. We support the very attractive hypothesis that two connected systems exist, the former one favours Pelagia's reproduction and acts as a nursery and the latter favours its growth due to higher productivity. We speculate that the reproductive population of the AIA is not permanent, but is renewed every year by individuals who have fed and quickly grown in the MS and who are passively transported by downwelling along canyon "corridors".
Similar content being viewed by others
Introduction
Gelatinous zooplankton are found throughout the world's oceans, but their species composition and abundance vary at local rather than global scales. The biomass of gelatinous zooplankton (Cnidaria, Ctenophora, Thaliacea) is greatest in productive coastal regions and is mainly related to sea surface temperature, dissolved oxygen, distance from shore, and primary production1. However, the occurrence of jellyfish in a wide range of environmental conditions suggests that they can adapt to different ecological niches. Despite the lack of scientific consensus on global long-term jellyfish trends2, their multiple negative impacts on human enterprises and ecosystem services3 are of concern4. Pelagic stages of Scyphozoa (scyphomedusae), particularly P. noctiluca, have been most commonly associated with negative impacts on coastal tourism, fisheries and aquaculture in the Mediterranean Sea5,6.
Unlike most scyphozoans, which have a metagenic life cycle with perennial polyps that require available substrate for attachment, Pelagia noctiluca is holoplanktonic. It inhabits both coastal waters and open oceans and is widespread in warm and temperate regions of the Atlantic and Pacific oceans7,8.
The success of their population in the Mediterranean Sea is reflected in their widespread distribution and increasingly frequent reports of blooms (outbreaks) in some coastal areas9. Marambio et al.10 consider P. noctiluca as the most common scyphozoan species in the recent review of jellyfish blooms in the western and central Mediterranean basins. Outbreaks have also been reported in the eastern Mediterranean along the coasts of Syria11, the Aegean Sea12 and Tunisia13. In contrast to the 1970s, 1980s and early 2000s, when Pelagia blooms occurred in the northern Adriatic Sea14, outbreaks in the last decade have been restricted to the central and southern Adriatic15.
Currently, Pelagia blooms in the western and central Mediterranean appear to be more common than observed until 199012,16,17,18,19,20,21,22. Climatic factors such as North Atlantic Oscillation23,24,25,26 and especially the influx of Atlantic Surface Water (AW) in the Mediterranean Sea27 influence the temporal fluctuations of P. noctiluca in this area.
The hypothesis is that nearshore blooms in the Mediterranean Sea could arise from permanent P. noctiluca populations drifting from offshore waters to the coast due to specific hydrodynamic conditions28,29. Suitable winds and currents could favour the accumulation of specimens and thus the development of blooms, especially in semi-enclosed areas such as bays and harbours13,21. Models have been used to infer connectivity between different Mediterranean Sea subregions and forecast Pelagia transport from the pelagic zone to the coasts with subsequent stranding9,22,30,31.
Population connectivity is central to their viability32. Ecological connectivity at the relevant time scale, conferring stability to the overall demographic system, may be crucial for short-lived pelagic species. Spatio-temporal dynamics and reconstruction of putative source-sink locations are more straightforward for meroplanktonic scyphomedusae when the source location, i.e., benthic polyp habitats, are known33. Otherwise, assumptions about preferred habitat characteristics34 and potential refugia must be made, especially for holoplanktonic species.
Submarine canyons, geomorphic features of continental margins, are seascapes that provide specific pelagic habitats. They are characterized by diverse and complex hydrodynamics and have important ecological roles35. Although still poorly studied, the inventory of canyons in the Mediterranean Sea has been compiled36. Recently, it has been suggested37 that submarine canyons may be critical refugia for the Mediterranean P. noctiluca. Field observations of blooms in the proximity of canyons along the Catalan coast9,38 support this hypothesis. "Slope index", the distance from the nearest marine canyon, was included as a predictor in habitat models10. Slope index was a significant predictor of potential P. noctiluca blooming areas for the northern Catalan coast, the Tunisian coast near Bizerte, the Ligurian Sea, the Strait of Sicily and the Southern Tyrrhenian Sea (STS) near the Aeolian Islands Archipelago (AIA)10.
This work focuses on the connectivity of populations in the STS (AIA) and the Strait of Messina (MS), combining data from field observations of P. noctiluca spatial distribution, stranding events, and numerical modelling. The modelling experiments were built on demographically relevant time scales consistent with previous life expectancy and reproduction findings. The average lifespan of holoplanktonic P. noctiluca was estimated to be nine months39. In the Mediterranean Sea, P. noctiluca spawns throughout the year40, with two seasonal peaks in spring and autumn41. The AIA and the MS provide complementary habitat qualities for P. noctiluca. The AIA has already been reported as a preferred area for the reproduction of P. noctiluca and formation of outbreaks9. The particular geomorphological and hydrological features of the MS have a remarkable influence on plankton production42,43 and create a specific habitat for P. noctiluca growth44.
This article aims to reveal the seasonal variability of P. noctiluca and its stranding events and unveil the connectivity between two western-central Mediterranean areas using high-resolution hydrodynamic modelling and Lagrangian particle tracking. We hypothesize that the STS and the MS act as a coupled system that enables the successful long-term maintenance of P. noctiluca populations. The latter is more productive, favours growth, and is the source of mature individuals who come back to the former that in turn provides better conditions for spawning and early development. Such a concept of interconnected systems may have broader applications also in other geographic areas.
Material and methods
Study sites
Aeolian Islands archipelago
AIA is located in the SE Tyrrhenian Sea. The STS is characterized by a surface west–east flow of modified Atlantic Water (AW). AW enters the area near the northern Sicilian coast, forming a broad cyclonic eddy that carries water towards the northern part of the sub-basin and moves geostrophically along the Italian coast45,46,47,48. According to recent findings on the variability of the surface circulation of the STS49, the AW current, while moving cyclonically northward, forms broad meanders around several anticyclonic cells located along the Italian coast during winter and spring (Fig. 1). The first anticyclonic cell is off the Sicilian coast, the second in the Gulf of Gioia Tauro, and the others are roughly off Naples and Rome. On a more local scale, a cyclonic subcell50,51 drags the AW out of the broad main eddy to enter the AIA basin from the northwest and flow southeastward toward the coasts of Sicily and MS. The estimated mean flows may be of the order of 5–10 cm s-1. As the summer season approaches, the aforementioned anticyclonic cells of the coastal recirculation along the Italian peninsula appear to shift offshore, towards the west. The winter flow pattern of AW collapses due to the weakening of the cyclonic wind stress curl, which reverses in summer and becomes anticyclonic in the region49.

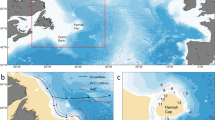
The eastern sector of the Southern Tyrrhenian Sea: boxes indicate the following sub-regions (a) Aeolian Island Archipelago; (b) Calabrian coast; (c) Sicilian coast; (d) Messina Strait. The main paths of surface circulation are depicted: the stream of AW flowing geostrophically and the anticyclonic recirculation cells. Map was generated with QGIS v. 3.22.6-Białowieża (https://qgis.org/it/site/) under GNU General Public License. Coordinate Reference System: EPSG:4326 (WGS84).
Strait of Messina
The MS separates Sicily from the Italian peninsula and connects the Ionian Sea in the south with the Tyrrhenian Sea in the north (Fig. 2). Detailed hydrographic information can be found in Vetrano et al.48 and Cucco et al.52 and references therein. From a hydrological point of view, the MS exhibits very relevant tidal currents driven by both barotropic and baroclinic processes, which depend on strong bathymetric forcings exerted mainly by the presence of a transverse sill (about 70 m deep) and a steep coastal morphology53. It has been reported that the maximum current intensity often exceeds 2 m s−1 in spring54. During the ebb phase ("scendente"), the Tyrrhenian surface water is directed into the MS and moves southward, while during the flood phase ("montante"), the currents are directed northward. The interaction with the topography and bathymetry of the channel leads to the formation of advective eddies and strong horizontal current shears, generally located in the lee of the prominent capes (Cape Peloro, Sicily and P.ta Pezzo, Calabria)54. The residual tidal flow (generally around 0.2–0.3 m s−1) shows the formation of an anticyclonic recirculation area just north of the sill with residual currents towards the Tyrrhenian Sea along the Sicilian coast (from Ganzirri to Cape Peloro) and towards the Ionian Sea along the Calabrian coast (Fig. 2).

The Strait of Messina: the main morphological and bathymetrical features are indicated. The residual current pattern (redrawn from52) is superimposed (black arrows): (i) the advective eddy at the entrance of the Strait, in the lee side of Cape Peloro (ii) the anticyclonic recirculation pattern just north of the sill and (iii) the cyclonic residual circulation south of the sill can be recognized. The black star indicates the position of the ENERMAR-Kobold platform that provided water temperature near the seabed and current velocity data; the black triangle indicates the CTD station (T/S profiles). The 1-km-long stretches of coast monthly surveyed for jellyfish strandings are highlighted in black (six along the Sicilian coast; three along the Calabrian coast). Map was generated by SHYFEM v. 7.5 with the embedded visualization tool “shyplot” (https://github.com/SHYFEM-model/shyfem).
The particular geomorphological characteristics of the MS lead to intense upwelling phenomena of Intermediate Ionian Waters in a specific area just south of the sill55. In contrast to other parts of the Mediterranean Sea, a stable thermocline can't develop in the MS, due to strong tidal currents and upwelling56. These conditions have a remarkable impact on the abundance and structure of planktonic communities43,56,57,58. Indeed, the total primary production rate has been estimated to range from 0.22 to 1.56 mg C m−2 h−142. This stimulating conditions lead to great biological richness and biodiversity in this ecosystem, as evidenced by the phytoplankton, zooplankton and fish communities43,57.
Habitat features in the Strait of Messina
To characterise the physical structure of the water column in this dynamic region, repeated vertical CTD casts were made down to 100 m depth (Fig. 2). Profiles were collected near the sill (38° 14′ 36ʺ N 15° 36′ E) every two weeks for 15 months from February 2003 to May 2004. A SeaBird 911 equipped with an altimeter was used. Overall twenty-nine available vertical profiles were analysed59,60.
In addition, automatic measurements from the Kobold ENERMAR platform anchored off the village of Ganzirri (38° 15′ 31ʺ N, 15° 37′ 40ʺ E) were used to characterize the water temperature at the seafloor (22 m depth) and the current regime near the Sicilian coast (Fig. 2). Water temperature measurements were collected during 2009–2010 (every 15 min from October 2009 to June 2010, every 1–6 h from June to October). Currents were measured every 15 min using a 3D ADCP (Nortek Aquadopp). In total, approximately 20,000 valid values of current intensities were acquired from October 2008 to May 200961.
Visual observations in coastal and open waters of the study areas
The Pelagia blooms, and spawning events in the Aeolian Archipelago have been documented and filmed over the past 18 years by diving expert Dario Lopes (pers. comm.). To observe the jellyfish larval stages (planulae and ephyrae) during blooms, samples were collected using a 2-L Niskin bottle, while juvenile and adult specimens were collected using a hand net. Collected individuals were observed and measured within 1 h using a stereomicroscope.
Monitoring of jellyfish presence and abundance was carried out almost daily from the beginning of March 2015 to January 2016 on board the Tuccoli T25 Sports Fishing "Suerte", a 7.06 and 2.47 m long and wide boat used for swordfish fishing in the Strait of Messina and adjacent areas (Fig. 3). Each survey started from the Tyrrhenian Sicilian coast (38° 16.516ʹ N, 15° 35.998ʹ E) early in the morning (6 am) and ended around 7 pm on the same day. The repeated outward and return trips lasted 4–6 h each and included alternately the MS, the Tyrrhenian coastal stretches of Sicily (from Messina to Milazzo) and Calabria (from Scilla to Gioia Tauro), and the region around AIA on days with good weather conditions (smooth sea, with waves, generally 0.2 m, light or weak breeze). The boat travelled at an average speed of 2 knots, which allowed good observations of the sea surface down to a depth of 5 m. Observations were made from the deck of the boat at a height of about 2 m above the sea surface, using a spotlight after sunset. An observation area of about 40 square metres was marked out around the boat. Currents, wind and sea state were noted during the cruise.

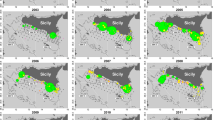
Visual observation effort in coastal and open waters sub-regions of the study area during March 2015–January 2016: (a) Aeolian Island Archipelago; (b) Calabrian coast; (c) Sicilian coast; (d) Messina Strait. Reported observations of Pelagia noctiluca in January–June (black dots) and July-December (empty triangles) are indicated. Maps were generated with QGIS v. 3.22.6-Białowieża (https://qgis.org/it/site/) under GNU General Public License. Coordinate Reference System: EPSG:4326 (WGS84).
A total of 181 surveys were conducted on different dates in the patrolled areas (Fig. 3). The 412 reported jellyfish occurrences were estimated by eye count and reported as number categories from 0 to 3: 0 no individuals observed, 1 for 1–10 individuals, 2 for 10–100 individuals and 3 for more than 100 individuals. Only in two cases were specific events with several hundred individuals were observed and marked as 'large aggregation'. These classes of abundance were considered sufficient to describe the distribution of P. noctiluca in the study area and are consistent with the earlier approach62.
To integrate available information within the Strait of Messina, data on the presence of jellyfish published by Rosa et al.21 and the Jellywatch CIESM monitoring from 2014 to 2015 (http://www.ciesm.org/marine/programs/jellywatch.htm) were included. Other validated information obtained through the Citizen Science program Meteomedusa, reports from well-known naturalists (youtube, video clips), beach guards, fishers or news from the local press (e.g. TempoStretto: info@tempostretto.it) were summarised.
Strandings of Pelagia in the Strait of Messina
Stranding events of epi-, meso- and bathy-pelagic organisms along the Sicilian and Calabrian coasts of the MS are a very common phenomenon observed for a long time63,64,65,66. This allowed us to identify the oceanographic, meteorological and geomorphological causes and consequently to select the most suitable beaches for the collection of stranded P. noctiluca.
Specifically, a survey of P. noctiluca abundance was conducted along the coast of the MS from March 2015 to February 2016, based on two surveys per week at each site. The observations were carried out along the following selected beaches (from north to south, Fig. 2) (a) Sicilian coast: Faro, Ganzirri, Pace, S. Francesco, and S. Raineri (b) Calabrian coast: Scilla, Cannitello and P.ta Pezzo. The survey took place in the morning and afternoon and essentially involved a careful inspection for Pelagia occurrences along the shoreline. Information on wind and current directions and weather conditions were recorded on the sampling days. In addition, the presence of Pelagia was also recorded within Messina Harbour, previously identified as a preferred location for large accumulations of floating debris.
In particular, about two stranding berms could be distinguished, of which the one farthest from the shore was characterized by the presence of pumice of the AIA, on the surface of which are barnacles, tubular annelids, and exceptionally, in the tiny cavities, small lamellibranchs. Pelagia was more commonly found along a berm closer to the shoreline, characterized by the presence of various pelagic organisms, and formed mainly in calm or slightly rough seas. For these reasons, beach sections approximately 1-km-long and 1-m-wide were surveyed by walking in both directions along identified stranding paths when there was more than one.
To determine the size/weight distribution of the stranded population, we calculated the average weight of jellyfish following Rosa et al.21. For specimens that were too damaged or dehydrated (especially in summer), we assumed that the stranded population was the same size as a reference specimen collected in the sea near the shoreline.
The numerical model
A high-resolution 3D hydrodynamic model (SHYFEM, https://github.com/SHYFEM-model/shyfem) was used to test some of our hypotheses about the transport and stranding of P. noctiluca. The model resolves the system of shallow water equations integrated over each layer in their formulations with water levels and transport67. It is based on the finite element method and has been successfully used in several applications and case studies at Mediterranean Sea68,69,70,71,72. The unstructured mesh resolves the model equations at a different spatial resolution, which is particularly useful when studying water circulation in both open ocean and coastal environments.
In particular, SHYFEM was used in a previous study52 to reproduce in detail the 3D water circulation caused by tides, heat and salt fluxes, and meteorological forcing in the MS, the STS, and the northern part of the Ionian Sea. In this study, the same model setup was used52, including the model mesh (Fig. 4) and boundary conditions, to reproduce the currents for the 2014–2015 period. This period was chosen since it includes most observations and measurements and is representative of recent years in terms of oceanographic and meteorological conditions52. The hourly model results (currents and winds) were used for the offline particle tracking simulations. The Particle Tracking Model (PTM) solves the advection and diffusion equations in a Lagrangian reference frame and allows the simulation of the path followed by active or passive numerical particles within the hydrodynamic model domain73,74,75,76.

The model computational domain as represented by the finite element mesh with bathymetric details. Red line delimits the seeding area for Experiment 1, yellow lines enclose the MS area where numerical particles were seeded during Experiment 2. Map was generated by SHYFEM v. 7.5 with the embedded visualization tool ”shyplot” (https://github.com/SHYFEM-model/shyfem).
The PTM uses the horizontal currents fields computed at a depth of 5 m to simulate the transport of the numerical particles and therefore neglects any vertical motion.
The contribution of wind to particle motion is very small and is assumed to be 4‰ of wind speed, as suggested by several modeling studies applying SHYFEM to reproduce the path followed by Lagrangian current- meters published in Western Mediterranean Sea70,72,73. In the Supplementary Note on model implementation, a detailed description of the model equation system and simulation setup is reported.
Three different modeling experiments were conducted:
EXP1 The first experiment was focused on investigating the dispersion of the early life stages of jellyfish from the spawning regions around the AIA toward the MS. The approximation of a purely horizontal displacement is assumed to hold for P. noctiluca larval stages (mostly ephyrae) which preferentially remain in the surface layer77 and therefore their passive transport can be approximated as mainly driven by the surface currents. In this simulation, the coastal and shelf waters of the AIA and Northern Sicily in the STS (west of the red line in Fig. 4) were assumed as possible spawning areas and sources of numerical particle seeding. About 10,000 passive Lagrangian particles were homogeneously released in the selected area with semi-diurnal frequency and their transport by surface currents and winds was computed by PTM.
Our observations show that reproduction in Aeolian waters is most intense from mid-winter to late spring (February–June), with minor reproduction also occurring in late autumn21,78. Accordingly, numerical particle seeding in our model was performed from late January to early June. Hourly positions of each released particle were used in the analysis. Relative probability of entering the MS as well as travel time were calculated for each release position. Probability was calculated using the Normalized Source Index (NSI), a dimensionless index obtained by normalizing and averaging the ratio of the number of numeric particles that entered the MS to the total number of particles released for each element of the computational domain. The NSI describes the relative degree of connectivity between each element of the seeding regions with the MS.
EXP2 The second simulation was conceived to evaluate the role of MS water circulation in promoting the retention and growth of Pelagia individuals. Horizontal passive transport of young/adult specimens by very strong barotropic currents dominates in the MS area so that their active vertical movements can be neglected to evaluate the trapping efficiency of the MS. Approximately 10,000 numerical particles were released uniformly with semi-diurnal frequency within the MS (between the yellow lines in Fig. 4). The trajectory of each particle was calculated and the relative abundance of particles within MS was estimated to determine the periods with the most favorable hydrodynamic conditions for trapping young Pelagia individuals within MS.
EXP3 The third numerical experiment was designed to simulate stranding of P. noctiluca in MS during summer, when habitat conditions are most favorable for jellyfish growth and the highest number of strandings was observed. The approximation of a purely horizontal displacement is motivated by considering that landing of the jellyfish individuals is promoted and ruled by surface forcings only, such as the wind and surface currents. In this case, the seeding of the particles within MS (between the yellow lines in Fig. 4) was performed only during the summer months (June, July, and August), resulting in the release of approximately 8 million particles throughout the simulation. GIS processing was performed on modelled particle positions to count stranding events for each of the 5-km coastal sections (Fig. 15).
The numerical experiments were designed to investigate the possible path of early life stages of P. noctiluca specimens from their spawning sites to feeding grounds and to explore the fate of young individuals in the MS during specific periods of the year. The questions the chosen approach sought to answer were: "Where do they come from? When do they arrive at MS? How long does it take them to reach MS? How long do they stay?".
Results
Observational data and habitat features
Environmental data and habitat features in the Strait of Messina
Analysis of the CTD vertical data show strong upwelling of deeper Ionian water with T and S values within a narrow range (14.0 ± 1 °C and 38.5 ± 0.3, respectively), especially near the submarine sill. Taking the density anomaly of 28.5 σt as a upper limit of the deeper waters rising to the surface, Fig. 5a highlights their recurrent presence throughout the whole water column. This limit is confined below 40–50 m depth from early spring (March–April) to autumn (November) leading to the formation of a two-layer system, while during winter with the onset of thermal homogenization these dense waters invade the whole column. This phenomenon regularly draws nutrient-rich intermediate Ionian waters into the euphotic layer, benefiting the entire food chain.

Habitat features relevant to P. noctiluca presence within the Messina Strait: (a) time series of bimonthly density anomaly profiles collected near the sill from February 2003 to April 2004: evidence of recurrent upwelling; (b) time series of water temperature at 20 m depth: optimal thermal range during the whole year; (c) frequency distribution of current intensity at 10 m depth near the Sicilian coast: active swimming very difficult and high risk of stranding.
A second important effect of this phenomenon can be observed by examining the dynamics of water temperature at a depth of 20 m (see Fig. 5b). In summer, strong temperature variations are observed due to the semi-diurnal tides that mix warmer Tyrrhenian surface water and colder upwelled Ionian water in the area. This contributes to the average temperature in the MS being lower than at the surface STS, where the temperature is often above 28 °C for several days in summer. On the other hand, the same phenomenon causes the water temperature in the Strait not to fall below 13.8–14 °C in winter. From a physiological point of view, the habitat in the MS is therefore in an optimal temperature range for Pelagia79, which is neither too hot in summer nor too cold in winter.
The third distinctive element of the habitat is the high current intensity which makes active swimming against current difficult for jellyfish. The currents dominate jellyfish transport which, given the turbulent regime, exposes jellyfish to significant stranding risks. The frequency distribution of the horizontal current intensity at 10 m depth is presented in Fig. 5c. It is interesting to note that less than 9% of time during slack the intensity is below 5 cm s−1, a threshold indicated by previous studies as the routine swimming speed for jellyfish80. Moreover, the intensity is on average about one order of magnitude above such value in both tidal phases (0.36 and 0.55 m s−1 respectively) and strongly asymmetric, being at least for 10% of time beyond 100 cm s−1.
Past documented P. noctiluca blooms and spawning events
Most of the documented and filmed intense blooms occurred between February and June. The first two documented blooms near Filicudi Island (see Fig. 1 for locations) are dated April 2008 and March 2009. One of the highest concentrations of Pelagia individuals was filmed near Vulcano Island in February 2010, while a large bloom was observed near Filicudi Island in May of the same year. In spring 2014, blooms were documented with videos and photos in both Salina (March) and Filicudi (March, May and June) (Lopez, pers. comm.). In particular, a dense bloom of Pelagia was documented in Filicudi on 2 June 2014, consisting of individuals in all developmental stages, from larvae to adults. The observed dense white cloud consisted of numerous planulae ranging in size from 0.34 to 0.36 mm. Among the larger specimens, adult jellyfish with a bell diameter of 70–80 mm were pink in colour (about 20%), while juvenile jellyfish (20–30 mm bell diameter) accounted for about 80%. Large aggregations of small (20–30 mm) and medium (50–60 mm) Pelagia specimens were observed in early July (3 and 16 July) near Basiluzzo Island (see Fig. 1 for location). In the same year, a massive occurrence of 2.5–3.5 mm ephyrae was noted during the winter months. Selected photos and videos of the reported mass events can be found in Supplementary Materials (V1).
These data confirm the assumption that AIA can be considered a natural ecosystem suitable for P. noctiluca reproduction from winter to late spring.
P. noctiluca occurrence and abundance patterns during 2015–2016
The boat surveys conducted in this study yielded nearly 2500 h of observations over a total distance of about 5000 nautical miles in the coastal and open waters of the region (Fig. 6). Direct observations confirmed the widespread occurrence of P. noctiluca at AIA and in the MS. Overall, Pelagia specimens were detected in about 80% of the surveys, and their abundance was high in almost 20% of the cases (> 100 ind., see red dots in Fig. 6). The occurrence was equally frequent in the MS and along the Tyrrhenian coast of Sicily (40–45% of reports each), while it was only occasional along the Calabrian coast. P. noctiluca were generally more abundant in the late morning (median 11:00 am) and rarely just before sunset, especially in summer.

Presence and abundance patterns of Pelagia noctiluca specimens in the study area as observed during the 181 surveys of the fishing boat “Suerte” in the period March 2015–January 2016. Abundance classes are in agreement with62. Maps were generated with QGIS v. 3.22.6-Białowieża (https://qgis.org/it/site/) under GNU General Public License. Coordinate Reference System: EPSG:4326 (WGS84).
Higher densities of P. noctiluca were observed mainly in spring and summer (Fig. 7). Two events of large Pelagia aggregation were recorded in front of the Tyrrhenian beaches of Eastern Sicily in May and June, respectively (Figs. 6, 7). More specifically, the two blooms were observed on May 6th and June 21st 2015 in the afternoon (5 pm) (lat 38° 16′ 41″ N, lon 15° 36′ 10″ E) both in calm seas and without wind. In May the extremely high abundance of P. noctiluca was associated with huge numbers of the colonial hydrozoan Velella velella.

Visual observations of Pelagia noctiluca presence in the study area (March 2015–January 2016): Seasonal distribution of relative percentages of events in the five classes of abundance.
The highest frequency of Pelagia observed in the Cape Peloro area (see Fig. 2 for location) and in the surrounding areas of the STS occurred during prevailing NW winds (about 50% in the year), the relative frequency of which was highest in May and June (95% and 80%, respectively). In contrast, variable winds from NE and south prevailed from July onwards, with the relative frequency of NW winds being lowest in October (8%). According to the repeated observations, the large quantities of Pelagia observed along the Tyrrhenian coast from Cape Rasocolmo (see Fig. 2 for location) to Cape Peloro are often carried at the Timpazzi site, about 15 km west of Cape Peloro, by the ebbing ("scendente" N-S) current into the Strait of Messina, where they are driven back to the coast by the flooding ("montante" S–N) current. From 8 to 10 July large quantities of small and medium-sized Pelagia specimens were observed between Pace and Ganzirri (see Fig. 2 for locations) in the ebbing swirls, which disappeared almost entirely with the onset of the tide. Large quantities of Pelagia were also observed on April 7th at the entrance of Messina Harbour during the high tide phase, when the wind was blowing from the NW.
Beach strandings
Observations of stranded Pelagia were conducted from March 2015 to February 2016. A total of 3162 specimens were counted, of which 58% along the Sicilian coast (five beaches plus Messina Harbour, see Fig. 2) and 42% along the Calabrian coast (three beaches). On a yearly basis, Faro was the one with the highest number of stranded specimens (28%) among the Sicilian beaches, followed by Ganzirri (22%), S. Francesco (9%), Pace (6%) and S. Raineri (5%). Natural or artificial bays seemed to influence strandings: among the studied beaches, Faro and Ganzirri, where most of the strandings occurred, are the richest in such structures. The highest number of Pelagia specimens was observed along the docks of Messina Harbour (31%, Fig. 8). Among the Calabrian beaches, Scilla was the one with the highest number of stranded Pelagia (43%), followed by Cannitello (35%) and P.ta Pezzo (22%).

Jellyfish beach strandings: Upper and central panels: Sicilian coast (from N to S); Lower panel: Calabrian coast (from N to S).
The relative frequency of stranded Pelagia on the Sicilian coast was high on days with easterly and southeasterly winds. At Faro and Ganzirri beaches, the highest number of stranded individuals was observed during southeasterly winds. Strandings at the southernmost beaches (e.g. S. Francesco) and along the Calabrian coast were mainly favoured by N/NE/NW winds. Strandings were highest at most sites during the winter and spring months (Fig. 8). They decreased in summer (July and August) and occurred in calm seas and light winds with variable directions. The stranded individuals collected were mostly damaged, colourless and usually without oral arms. The average percentage of bell diameter size classes recorded on the Sicilian and Calabrian beaches in the different seasons is presented in Table 1. The two largest stranded specimens with bell diameters of 102 mm and 124 mm were collected in January and February.
The biomass of stranded P. noctiluca was estimated using the equation W = 0.0002 D2.8786 developed for the study area21. W is the total wet weight in grammes, and D is the bell diameter in mm. As shown in Fig. 9, the highest stranded Pelagia biomass was recorded on the Sicilian coast during the winter months (WW = 13,000 g, 42% of the total stranded biomass), with a decreasing seasonal trend until autumn, when the lowest value was recorded (WW = 2500 g, 8%). On the Calabrian coast, the highest biomass was obtained in spring (WW = 15,000 g, 47%), followed by similar values in winter and summer, to reach the lowest value in autumn (WW = 500 g, 2%). Although fewer beaches were considered on the Calabrian coast than on the Sicilian coast (3 versus 5 beaches), the cumulative biomass values were the same: 31,000 g WW (50%).

Seasonal trend of cumulated biomass of P. noctiluca stranded specimens along the coasts of Sicily (4 sites) and Calabria (3 sites) from March 2015 to February 2016.
The numerical experiments
From spawning to nursery (EXP1)
The first experiment results showed that two separate spawning areas contribute a relatively high number of particles to MS. These are clearly distinguishable in Fig. 10. The smaller area, located between the northern Sicilian coast and south of AIA, has a lower NSI with values between 0.7 and 0.8, while the larger area is located between the Aeolian islands of Salina, Stromboli and Vulcano and has an NSI above 0.9. The coastal waters around Lipari and Panarea have NSI values close to 1 and are therefore the most likely source of P. noctiluca in MS. The particle concentration timeline for MS is shown in Fig. 11. The first particles enter the MS about 25 days after the first release (January), and the maximum number is reached in June (day 170–175). After reaching the maximum concentration, the fraction of particles decreases until the end of the year with an almost constant rate and minimal disturbances. These results show the maximum influx of P. noctiluca in late spring and early summer and agree well with field observations. Transport time from spawning areas to MS was estimated based on particle age and is shown in Fig. 12. Particles from nearby areas reach MS within a few days, while others take up to 30 days. Particles from the areas with the highest NSI (Lipari and Panarea) reach MS in about 20 days.

Distribution of the relative probability of arriving in the Strait starting from the selected spawning areas. The probability has been computed through the Normalized Source Index (NSI, hereafter), a non-dimensional index obtained by normalizing and averaging the ratio between the number of numerical particles entering in the Strait of Messina and the total number of particles released for each element of the computational domain. Map was generated by SHYFEM v. 7.5 with the embedded visualization tool ”shyplot” (https://github.com/SHYFEM-model/shyfem).

The cumulative ratio between the total amount of particles entering in the Strait and the amount of particles released within the spawning areas and still within the model domain. The results were post-processed and averaged on a daily basis in order to obtain a whole year dataset from the two-year simulation runs results. The time series of the ratio is reported as a function of the time.

Distribution of average travel times to the Strait of the particles starting from the spawning areas with NSI higher than 0.6. These times correspond to the ages of the particles at the entering into the Strait of Messina. Map was generated by SHYFEM v. 7.5 with the embedded visualization tool ”shyplot” (https://github.com/SHYFEM-model/shyfem).
Living in the Strait (EXP2)
In the second experiment, we investigated the suitability of hydrographic conditions for the retention of P. noctiluca in the MS. Figure 13 shows the number of particles in the MS for each day of the year. There are three distinct peaks in April, June and July. These are the periods that should provide the most favourable environment for P. noctiluca growth. Late spring and early summer also coincide with the time when the jellyfishes from AIA should arrive. We also evaluated the fraction of particles that end up in the STS (Fig. 14). The results suggest that particles are retained in the MS during the first part of the year, while the escape of particles into the STS is favoured thereafter.

Daily amount of numerical particles inside the Strait throughout the year. A total of 20,000 particles were released every 24 h in the selected spawning areas around the Aeolian Island Archipelago.

Evaluation of the retention efficiency of the Strait: ratio between the amount of numerical particles leaving the Strait to the Tyrrhenian Sea and the amount of numerical particles entering in the Strait.
Stranding (EXP3)
In Fig. 15, we plot the percentage of particles that are stranded at different parts of the coast. The stranding probability is only slightly higher on the Calabrian coast (53%) than on Sicilian Coast (47%). In Sicily, strandings are concentrated in the section between Cape Peloro (north) and Messina Harbour (south; see Fig. S1). Here, the shape of the coastline favours the trapping of numerical particles. In Calabria, most strandings occur between Scilla and P.ta Pezzo (about 40%). The remaining strandings extend over a much longer part of the coast than in Sicily.

Modeled stranding shares (%) per each coastal element along the coasts of the Strait of Messina. Sicilian and Calabrian coasts were subdivided into 5 km-long stretches (clockwise from Sic1 to Sic17 for Sicily and counter clockwise from Cal1 to Cal25 for Calabria respectively) to count the stranding events. Along the Sicilian coast: Faro and Ganzirri beaches lay in Sic6; Pace beach lays in Sic7; S. Francesco beach lays in Sic8; Messina Harbour lays in Sic9; S. Raineri lays in Sic10. Along the Calabrian coast: Scilla beach lays in Cal10; Cannitello beach lays in Cal11, P.ta Pezzo beach lays in Cal12. Map was generated with QGis v. 3.22.6-Białowieża (https://qgis.org/it/site/) under GNU General Public License. Projected Coordinate System: EPSG:32,633 (WGS 84/UTM zone 33 N).
Strandings comparison with in situ observations (EXP3)
We compared the results of the EXP3 experiment with observed P. noctiluca strandings. Considering the fragility of stranded individuals, they should not last much longer than one day, and therefore the number of strandings should be approximately equal to the daily stranding rate. We compared observed strandings for the same three months used in the simulation (June, July, and August = 92 days). According to our observational estimate, there were about 440,000 stranded individuals on the Calabrian beaches and about 280,000 stranded individuals on the Sicilian beaches during this period corresponding to a ratio between the individuals stranded along the two coastlines of about 1.5. Similar behavior was observed from the model simulations results: the ratio between the total particles stranded on the Calabrian coast and the particles stranded on the Sicilian coast was around 1.4, slightly underestimating the value obtained from the observations. This means that the model results are in good agreement with the empirical estimates.
Discussion and conclusions
Considering that, during the early life stages, P. noctiluca spends most of the time on the surface layers without performing intense vertical migration77 and that during the maturity within the MS, the horizontal components of the transport are prevalent with respect to the vertical one, the approximation of a purely horizontal displacement of the particles was selected for all the numerical simulations. Specifically, in the MS the intense horizontal currents, up to 2 m s−1, are mainly barotropic and quite homogeneous along the vertical52. Therefore, the horizontal transport of the adult individuals, which perform intense nictemeral displacement, can be only partially affected by the vertical variability of the tidal flow, and the transport at the surface can be considered as an acceptable approximation for investigating the trapping processes within this area. Furthermore, the vertical components of the velocity fields computed by the hydrodynamic model were quite weak and around 1, 2 magnitude orders lower than the horizontal flow52, and, therefore, neglectable in the transport computation.
It should be noted that climatic factors are, however, expected to impact on the interannual fluctuations of P. noctiluca in this area. Indeed, the analysis of available long-term series of ocean dynamics and thermodynamics numerical model solutions81,82 and in situ and satellite observations83 have highlighted how interannual-to-decadal changes in the Mediterranean Sea and sub basins are a result of the decadal scale changes of the Northern Hemisphere atmospheric regimes, related to the variability of the North Atlantic Oscillation. Specifically, in the surroundings of the MS, the South-Eastern Tyrrhenian cyclonic Gyre81 and the Northern Sicily anticyclone83, at north, and the Atlantic Ionian Stream, at south, show noticeably amplitude variations, and a current reversal can take place in the Ionian Sea81.
The current knowledge of the interannual variability in the surroundings of the MS is hence mostly referred to a strengthening or weakening of the observed ocean circulation structures. Their effects on the highly variable occurrence of jellyfish blooms, over multiple years, should be carefully verified through ad hoc, high-resolution and long-period numerical simulations that should be supported by appropriate collections of blooms observations.
Although very different, the analyzed ecosystems of the Aeolian Islands and Messina Strait are complementary for the success of the P. noctiluca life cycle. Model results support the hypothesis of connectivity between these two ecosystems, especially in the first half of the year, suggesting that the coastal areas around the AIA serve as a potential source site for Pelagia in MS. Conversely, the MS is an area where immature specimens (20–30 mm) reach the average size of 40–50 mm in about three months. A growth rate of 0.28–0.30 mm d−1 and a rapid increase in biomass (0.73–1.13 g d−1) are greater than those recorded for other Mediterranean areas21. On the other hand, the MS does not seem to be optimal for Pelagia's reproduction for two main reasons. The large short-term fluctuations in water temperatures (Fig. 5b), which are generally colder than the average of the Southern Tyrrhenian Sea, are mainly outside the optimal temperature range for Pelagia reproduction79,84. Moreover, the strong currents, far beyond the swimming capabilities of Pelagia80, do not favour large aggregations in which fertilization of broadcasted gametes is promoted. As a matter of fact, even if P.noctiluca specimens with mature gonads have been found, no planulae/ephyrae presence is reported by the authors who consider the MS a reproduction area21,41.
Massive blooms of P. noctiluca have been observed to occur at sites closest to the upper margins of marine canyons9,38, such as AIA basin9. The formation of these large aggregations facilitates the encounter of broadcasted gametes85, especially when thousands of specimens are in close contact. To enhance the success of sexual reproduction, the already observed frequent formation of individuals pairs during swarms9 was confirmed by us in two other configurations (Fig. S2).
Based on our observations and underwater videos showing new cohorts of planulae, ephyrae and juvenile medusae swarms in the Aeolian surface waters, we assume Pelagia reproduction in the AIA occurs from mid-winter to late spring (February-June). Reproduction may also occur in late autumn. Although Pelagia can reproduce throughout the year21,40,86, the main breeding seasons are spring and autumn at temperatures of 16–22 °C79,84,87. Water temperatures in mid-winter to late spring (17–22 °C) and autumn (18–26 °C) are characteristic of the AIA88,89 and are thus within the optimal range for Pelagia reproduction.
The AIA basin is an important breeding, spawning and nursery area for many commercially important pelagic and coastal fishes. It is a typical oligotrophic area90,91 with high species diversity92,93,94,95. The basin is outside the influence of upwelling, the thermocline is deeper, and the primary production and biomass of mesozooplankton are moderate96,97,98,99,100. These environmental and ecological characteristics, together with field observations, suggest that the archipelago may be a breeding site and an area for early larval development but the low productivity does not support the rapid growth of P. noctiluca.
The distribution and intensity of the Pelagia blooms depend largely on shelf topography, wind direction, and food concentration101. In the first half of the year, when northwesterly winds prevail, currents progress eastward from the AIA basin toward MS. As model simulations show, juvenile Pelagia specimens can take advantage of this direct connectivity link to reach MS from spawning grounds in just over 20–30 travel days on average. This is consistent with immature Pelagia specimens of about 20–40 mm bell diameter found in MS during the late winter-early spring months21.
Rosa et al.21 observed that the abundance of juvenile jellyfishes (20–30 mm) recorded from January to April at sampling sites in MS was only 4% of the total yearly Pelagia counts, indicating high larval mortality in winter. In addition, only jellyfishes greater than 20 mm were counted, as the smaller individuals were transparent and barely visible. Based on our data, we believe that the number of juvenile jellyfishes (up to 20–30 mm) entering MS in winter-spring is likely much higher than that already reported21. As shown by the winter-spring model simulations, particles are efficiently trapped during this period thanks to tidal back and forth movements102 and the presence of the residual recirculation cell near Cape Peloro.
From January to June, the number of particles entering the MS is higher than the number of particles moving from the MS to the STS (see “Living in the Strait (EXP2)” section Experiment 2). In contrast, the quantities are similar from June onwards, indicating a significant flux of particles from MS into the STS. Considering that the flow of seeded particles was homogeneous throughout the year (see “The numerical model” section Experiment 2) and that spawning occurs mainly in the first half of the year, the hypothesis that the northward flow drives juvenile Pelagia out of the MS from June to late summer is strengthened.
P. noctiluca was frequently observed at the entrance of the MS, in an area just north of Cape Peloro, which was probably related to the accumulation effect of the residual advective cyclonic eddy. Moreover, the large number of P. noctiluca along the Sicilian coast of the MS is closely associated with the effect of the residual circulation cells: once the Pelagia swarms enter the MS during the ebbing phase, they are driven northward by the main anticyclonic circulation or, alternatively, they can be trapped by the outflowing current along the Sicilian coast and move southward to accumulate within the Messina Harbour. Our results suggest that large numbers of Pelagia enter the MS from the STS when the winds are blowing from the NW and with ebbing currents. Their presence and abundance in the surface waters of the MS can change during the course of a day, from large densities at low tide to few specimens at high tide.
At the end of the spring, when the residual current is still low, and the number of particles in the MS reaches its maximum, the hydrodynamic regime in the Strait is dominated by tides52. The tidal dynamics in the MS, even if characterized by high instantaneous currents of up to 2.5 m s−1, has a relatively low residual circulation pattern, resulting in higher water residence times than in other seasons52. Comparing the obtained results with the numerical analysis, a coherence was observed between the periods with the highest particle arrival rate between the 150th and 175th day and the period with the highest particle residence times in MS, between the 130th and 180th day. These results further support the hypothesis that "Pelagia feed and grow in early summer in MS". In autumn and winter, surface circulation in the MS carries water masses into the Ionian and Tyrrhenian Seas, shortening the residence time of particles in MS52.
This study highlights the limitations of Pelagia (and other gelatinous zooplankton) distribution observations derived from fixed coastal sites. Physical factors such as winds influence the formation of aggregations and the occurrence of strandings and currents move the swarms along103. Thus, it happens that P. noctiluca swarms crossing the waters of MS are not recorded in shore-based observations, such as the bloom (about 600 ind. m−3) reported by fishermen on 28 April 2019, which was carried southward by the strong ebb current in the central part of MS. This strengthens the concept of the pelagic habitat of P. noctiluca and, consequently, the close correlation of this species with the circulation of water masses, upwelling systems and frontal zones9,21,29. Pelagia may indeed be passively transported by horizontal currents, so their temporal and spatial distribution results from the interplay of population dynamics and dispersal by currents31. In addition, some biological traits, such as vertical migration104 could improve the prediction of Pelagia noctiluca dispersal. It is difficult to assess these processes from observations alone, as neither in situ nor remote observations alone provide a continuous picture of circulation31. Therefore, we agree that modelling studies linking physical transport and monitoring of jellyfish outbreaks in coastal waters should be conducted28. Pelagia survival is lower in shallow waters because physical stresses damage their gelatinous bodies (bottom contact, wave action), daily migration is significantly disrupted, and predation by fish plays an important role105.
Stranding events on the Calabrian and Sicilian coasts of the MS are not a constant phenomenon with precise seasonal cycles that repeat themselves over the years. Rather, they must be considered as the result of various causes (biological, hydrological, meteorological) that determine more or less favourable periods for their occurrence. From an ecological point of view, though representing a net demographic and biomass loss due to the habitat boundary conditions, the massive stranding events and overall rates of stranding are accounted for in the adaptation strategy of the species to the habitat and can be sustained by the population of Pelagia, that receives in turn from the MS a sheltered and nutrient-rich environment during the central stages of the life cycle. The areas where Pelagia washed up in large numbers on the coasts are popular tourist areas. Our study agrees with the observations that the element of predictability of jellyfish strandings can help beach management authorities to plan and manage beach use and swimming activity when swarms of jellyfish appear near the shore106.
Finally, to close the biological cycle, we propose two different mechanisms of transporting the adult P. noctiluca individuals to the nursery in the Aeolian Islands. The hypothesis of Boero (in Sacchetti37), currently accepted also by many others9,44,107, seems plausible. Jellyfish are thought to migrate downward to reach intermediate water depths, possibly along canyon corridors. We believe that P. noctiluca individuals from MS are passively transported to deep Tyrrhenian waters through downwelling processes that occur primarily during well-stratified water conditions (summer and late summer). These processes are related to the interaction of internal solitary waves generated by tidal currents at the Ganzirri-P.ta Pezzo sill with seabed morphology (e.g. Gioia Tauro canyon and Calabrian slope), and propagating northwards in the Tyrrhenian Basin108,109,110. This phenomenon has been known for a long time, ever since Lohmann's111 valuable study of the distribution of appendicularians in the MS. Plankton species such as Oikopleura longicauda were used to observe and evaluate the deep downward currents. Later, this process was confirmed by the discovery of surface water indicator species, such as phytoplankton and chaetognaths, in the deep layers of the STS57,112. Marine canyons are known as "superhighways" due to the rapid circulation of water masses, sediments and organisms during active or passive movements from shallow to deeper waters and vice versa9,36,113,114,115. Canyons play an important role in structuring populations and life cycles of planktonic fauna116. On the northern margin of Sicily, a dense network of submarine canyons has been discovered in the depth range 80–2100 m, connecting the Sicilian and Tyrrhenian marine areas117. Intermediate waters may also provide an alternative, abundant crustacean resources for P. noctiluca (e.g., euphausiid crustaceans114). Thus, this jellyfish may spend the warmer months in deeper habitats along the continental slope with abundant food sources and invest more energy in future sexual reproduction through germ cell differentiation and gonad maturation9.
The other option of providing suitable transport of adult P. noctiluca individuals to the AIA would be by cyclonic currents in the south-eastern Tyrrhenian Sea. These could transport the individuals northward along the Italian coastline and then westward towards the AIA. The average summer geostrophic currents indicate possible routes of this type49. These currents are prone to instabilities and meanders and exhibit high interannual variability49. Our results can’t support or reject any of the two proposed mechanisms of transport towards the AIA. In fact both might be at work and due to considerable interannual variability of the Tyrrhenian circulation their contributions can be variable as well.
Our data support the hypothesis that there are two coupled (connected) systems, one of which favours Pelagia's reproduction and serves as a nursery (in our case, the AIA), and the other favours its growth due to higher productivity (in our case, the MS). Therefore, we believe that the reproductive population of the AIA is not permanent but is renewed every year by individuals that have fed and grown rapidly in the MS and are transported by the ocean circulation. The most plausible is the transport by downwelling along the "corridors" of the canyons. This is precisely the phase that would complete the annual life cycle of Pelagia. Such a concept could have a broader impact if proven for other similar 'coupled' systems in the Mediterranean or elsewhere.
References
Lucas, C. H. et al. Gelatinous zooplankton biomass in the global oceans: Geographic variation and environmental drivers. Glob. Ecol. Biogeogr. 23, 701–714. https://doi.org/10.1111/Geb.12169 (2014).
Condon, R. H. et al. Recurrent jellyfish blooms are a consequence of global oscillations. Proc. Natl. Acad. Sci. USA 110, 1000–1005. https://doi.org/10.1073/pnas.1210920110 (2013).
Graham, W. M. et al. Linking human well-being and jellyfish: Ecosystem services, impacts, and societal responses. Front. Ecol. Environ. 12, 515–523. https://doi.org/10.1890/130298 (2014).
Lucas, C. H., Gelcich, S. & Uye, S. I. Living with jellyfish: Management and adaptation strategies. In Jellyfish Blooms (eds Pitt, K. A. & Lucas, C. H.) 129–150 (Springer, 2014).
De Donno, A. et al. Impact of stinging jellyfish proliferations along south Italian coasts: Human health hazards, treatment and social costs. Int. J. Environ. Res. Public Health 11, 2488–2503 (2014).
Bosch-Belmar, M. et al. Consequences of stinging plankton blooms on finfish mariculture in the Mediterranean Sea. Front. Mar. Sci. 4, 240. https://doi.org/10.3389/fmars.2017.0024 (2017).
Mayer, A. G. Medusae of the World: The Hydromedusae 132–498 (Carnegie institution of Washington, 1910).
Kramp, P. L. Synopsis of the medusae of the world. J. Mar. Biol. Assoc. UK 40, 1–469 (1961).
Canepa, A. et al. Pelagia noctiluca in the Mediterranean Sea. In Jellyfish Blooms (eds Pitt, K. A. & Lucas, C. H.) 237–266 (Springer, 2014).
Marambio, M. et al. Unfolding jellyfish bloom dynamics along the Mediterranean basin by transnational citizen science initiatives. Diversity 13, 274. https://doi.org/10.3390/d13060274 (2021).
Mamish, S., Durgham, H. & Ikhtiyar, S. The first Pelagia noctiluca outbreak off the Syrian coast (the eastern Mediterranean Sea), five years after its first appearance. SSRG Int. J. Agric. Environ. Sci. 6, 72–75 (2019).
Daly Yahia, M. N. et al. Are outbreaks of Pelagia noctiluca (Forsskäl, 1775) more frequent in the Mediterranean Basin?. ICES Coop. Res. Rep. 300, 8–14 (2010).
Aissi, M., Touzri, C., Gueroun, S. K. M., Kefi-Daly Yahia, O. & Daly Yahia, M. N. Persistent occurrence and life cycle of Pelagia noctiluca in the channel of Bizerte (Northern Tunisia). Ecol. Environ. Conserv. 20, 1453–1460 (2014).
Kogovsĕk, T., Bogunović, B. & Malej, A. Recurrence of bloom forming scyphomedusae: Wavelet analysis of a 200-year time series. Hydrobiologia 645, 81–96 (2010).
Pestoric, B. et al. Scyphomedusae and ctenophora of the eastern adriatic: Historical overview and new data. Diversity 13, 186. https://doi.org/10.3390/d13050186 (2021).
UNEP (United Nations Environmental Programme). Workshop on Jellyfish Blooms in the Mediterranean, Athens (1984).
UNEP (United Nations Environmental Programme). Jellyfish blooms in the Mediterranean Sea. Proceedings of II Workshop on Jellyfish in the Mediterranean Sea, Athens (1991).
Goy, J., Morand, P. & Etienne, M. Long term fluctuations of Pelagia noctiluca (Cnidaria, Scyphomedusa) in the western Mediterranean. Sea Prediction by climatic variables. Deep-Sea Res. A 36, 269–279 (1989).
Bernard, P., Berline, L. & Gorsky, G. Long term (1981–2008) monitoring of the jellyfish Pelagia noctiluca (Cnidaria, Scyphozoa) on the French Mediterranean Coasts. J. Oceanogr. Res. Data 4, 1–10 (2011).
Brotz, L., Cheung, W. W. L., Kleisner, K., Pakhomov, E. & Pauly, D. Increasing jellyfish population: Trends in large marine ecosystems. Hydrobiologia 690, 3–20 (2012).
Rosa, S., Pansera, M., Granata, A. & Guglielmo, L. Interannual variability, growth, reproduction and feeding of Pelagia noctiluca (Cnidaria: Scyphozoa) in the Straits of Messina (Central Mediterranean Sea): Linkages with temperature and diet. J. Mar. Syst. 111–112, 97–107 (2013).
Aoutien, M., Bekkali, R., Nachit, D., Luan, K. & Mrhraoui, M. Predicting jellyfish strandings in the Moroccan North-West Mediterranean coastline. Eur. Sci. J. 15, 72–84. https://doi.org/10.19044/esj.2019.v15n2p72 (2019).
Lynam, C. P., Hay, S. J. & Brierley, A. S. Interannual variability in abundance of North Sea jellyfish and links to the North Atlantic Oscillation. Limnol. Oceanogr. 49, 637–643 (2004).
Lynam, C. P. et al. Have jellyfish in the Irish Sea benefited from climate change and overfishing?. Glob. Change Biol. 17, 767–782 (2011).
Brodeur, R. D. et al. Rise and fall of jellyfish in the eastern Bering Sea in relation to climate regime shifts. Prog. Oceanogr. 77, 103–111 (2008).
Molinero, J. C. et al. Climate control on the longterm anomalous changes of zooplankton communities in the Northwestern Mediterranean. Glob. Change Biol. 14, 11–26 (2008).
Licandro, P. et al. A blooming jellyfish in the northeast Atlantic and Mediterranean. Biol. Let. 6, 688–691 (2010).
Ferraris, M. et al. Distribution of Pelagia noctiluca (Cnidaria, Scyphozoa) in the Ligurian Sea (NW Mediterranean Sea). J. Plankton Res. 34, 874–885 (2012).
Malačič, V., Petelin, B. & Malej, A. Advection of the jellyfish Pelagia noctiluca (Scyphozoa) studied by the Lagrangian tracking of water mass in the climatic circulation of the Adriatic Sea. Geophys. Res. Abstr. 9, 02802 (2007).
Rubio, P. & Muñoz, J. M. Predicción estival del riesgo de blooms de Pelagia noctiluca (litoral central catalán). In Situaciones de riesgo climático en España (ed. Novau, J. C.) 281–287 (Instituto Pirenaico de Ecología, 1997).
Berline, L., Zakardjian, B., Molcard, A., Ourmieres, Y. & Guihou, K. Modeling jellyfish Pelagia noctiluca transport and stranding in the Ligurian Sea. Mar. Pollut. Bull. 70, 90–99 (2013).
Olds, A. D. et al. Quantifying the conservation value of seascape connectivity: A global synthesis. Glob. Ecol. Biogeogr. 25, 3–15 (2016).
Vodopivec, M., Peliz, A. J. & Malej, A. Offshore marine constructions as propagators of moon jellyfish dispersal. Environ. Res. Lett. 12, 084003 (2017).
Chen, J. Z., Huang, S. L. & Han, Y. S. Impact of long-term habitat loss on the Japanese eel Anguilla japonica. Estuar. Coast. Shelf Sci. 151, 361–369 (2014).
Fernandez-Arcaya, U. et al. Ecological role of submarine canyons and need for canyon conservation: A review. Front. Mar. Sci. 4, 5. https://doi.org/10.3389/fmars.2017.00005 (2017).
Würtz, M. Towards a Mediterranean canyon inventory. Workshop (EBSAs), 7 to 11 April 2014, Málaga, Spain, 1–4 (2014).
Sacchetti, F. Il ritorno di MeteoMedusa. Focus (Madison) 237, 92–94 (2012).
Benedetti-Cecchi, L. et al. Deterministic factors overwhelm stochastic environmental fluctuations as drivers of jellyfish outbreaks. PLoS ONE 10, e0141060. https://doi.org/10.1371/journal.pone.0141060 (2015).
Malej, A. & Malej, M. Population dynamics of the jellyfish Pelagia noctiluca (Forsskäl, 1775). In Proceedings of the 25th EMBS, Marine Eutrophication and Population Dynamics (ed. Colombo, G.A.) 215–219 (Olsen & Olsen, 1992).
Rottini-Sandrini, L., Avian, M., Axiak, V. & Malej, A. The breeding period of Pelagia noctiluca (Scyphozoa, Semaeostomeae) in the Adriatic and central Mediterranean Sea. Nova Thalass. 6, 65–75 (1983).
Milisenda, G. et al. Reproductive and bloom patterns of Pelagia noctiluca in the Strait of Messina, Italy. Estuar. Coast. Shelf Sci. 201, 29–39. https://doi.org/10.1016/j.ecss.2016.01.002 (2018).
Magazzù, G. et al. Picoplankton: Contribution to phytoplankton production in the Strait of Messina. Mar. Ecol. 8, 21–31 (1987).
Guglielmo, L., Crescenti, N., Costanzo, G. & Zagami, G. Zooplankton and micronekton communities in the Straits of Messina. In The Straits of Messina ecosystem, present knowledge for an ecohydrodynamical approach. Proceedings of Symposium held in Messina, 4–6 April 1991, Messina (eds. Guglielmo, L., Manganaro, A. & De Domenico, E.) 247–270 (Dipartimento di Biologia Animale ed Ecologia, 1995).
Guglielmo, L. et al. The Strait of Messina: A key area for Pelagia noctiluca (Cnidaria, Scyphozoa). In Jellyfish: Ecology, Distribution Patterns and Human Interactions (ed. Mariottini, G. L.) 71–90 (Nova Science Publishers Inc., 2017).
Astraldi, M. & Gasparini, G. P. The seasonal characteristics of the circulation in the Tyrrhenian Sea. In: Seasonal and Interannual Variability of the Western Mediterranean Sea, Coast. Estuar. Studies, Vol. 46, 115–134 (American Geophysical Union, 1994).
Krivosheya, V. G. Water circulation and structure in the Tyrrhenian Sea. Oceanology 23, 166–171 (1983).
Millot, C. Circulation in the Western Mediterranean Sea. J. Mar. Syst. 20, 423–442. https://doi.org/10.1016/S0924-7963(98)00078-5 (1999).
Vetrano, A., Napolitano, E., Iacono, R., Schroeder, K. & Gasparini, G. P. Tyrrhenian Sea circulation and water mass fluxes in spring 2004: Observations and model results. J. Geophys. Res. 115, C06023 (2010).
Iacono, R., Napolitano, E., Marullo, S., Artale, V. & Vetrano, A. Seasonal variability of the Tyrrhenian Sea surface geostrophic circulation as assessed by altimeter data. J. Phys. Oceanogr. 43, 1710–1732. https://doi.org/10.1175/JPO-D-12-0112.1 (2013).
Boero, F. et al. CoCoNet: Towards coast to coast networks of Marine Protected Areas (from the shore to the high and deep sea), coupled with sea-based wind energy potential. Sci. Res. Inf. Technol. 6(Suppl.), 1–95 (2016).
Rio, M. H. et al. A mean dynamic topography of the Mediterranean Sea computed from altimetric data, in-situ measurements and a general circulation model. J. Mar. Syst. 65, 484–508 (2007).
Cucco, A. et al. Hydrodynamic modelling of coastal seas: The role of tidal dynamics in the Messina Strait, Western Mediterranean Sea. Nat. Hazards Earth Syst. Sci. 16, 1553–1569 (2016).
Hopkins, T. S., Salusti, E. & Settimi, D. Tidal forcing of the water mass interface in the Straits of Messina. J. Geophys. Res. 89, 2013–2024 (1984).
Bignami, F. & Salusti, E. Tidal currents and transient phenomena in the Strait of Messina: A review. In: The Physical Oceanography of Sea Straits (ed. Pratt, L. J.) 95–124 (Kluwer Academic, 1990).
Azzaro, F., Decembrini, F., Raffa, F. & Crisafi, E. Seasonal variability of phytoplankton fluorescence in relation to the Straits of Messina (Sicily) tidal upwelling. Ocean Sci. Discuss. 4, 415–440 (2007).
De Domenico, E., Cortese, G. & Pulicanò, G. Chemical characteristics of the waters in the Straits of Messina. In The Straits of Messina ecosystem, present knowledge for an ecohydrodynamical approach. Proceedings of Symposium held in Messina, 4–6 April 1991, Messina (eds. Guglielmo, L., Manganaro, A., & De Domenico, E.) 155–167 (Dipartimento di Biologia Animale ed Ecologia Marina, 1995).
Guglielmo, L. Distribuzione di Chetognati nell’area idrografica dello Stretto di Messina. Pubbl. Staz. Zool. Napoli 40, 34–72 (1976).
Sitran, R., Bergamasco, A., Decembrini, F. & Guglielmo, L. Temporal succession of tintinnids in the northern Ionian Sea, Central Mediterranean. J. Plankton Res. 29, 495–508 (2007).
AA.VV. Final Scientific Report of the Project Cluster 10—SAM "Realizzazione ed attivazione di una rete integrata di piattaforme costiere e mezzo mobile attrezzati per Sistemi Avanzati di Monitoraggio delle acque (SAM)", funded by the Italian Ministry of University and Scientifical and Technological Research (MURST), Internal Data File, Istituto Sperimentale Talassografico, National Research Council, Messina, Italy (2005).
Sitran, R. Caratterizzazione dei popolamenti microzooplanctonici nell’area idrografica dello Stretto di Messina, University of Messina, Ph.D. Thesis XVII cycle (2006) (in Italian).
Bergamasco, A. et al. A laboratory for the observation of a highly-energetic coastal marine system: The Straits of Messina. In Volume DTA/06–2011, “Marine Research at CNR” 2185–2202 (Department of Earth and Environment of National Research Council, 2011).
Doyle, T. K. et al. Widespread occurrence of the jellyfish Pelagia noctiluca in Irish coastal and shelf waters. J. Plankton Res. 30, 963–968 (2008).
Guglielmo, L. Spiaggiamenti di eufausiacei lungo la costa messinese dello Stretto dal dicembre 1968 al dicembre 1969. Boll. Pesca Piscic. Idrobiol. 24, 71–77 (1969).
Guglielmo, L., Costanzo, G. & Berdar, A. Ulteriore contributo alla conoscenza dei crostacei spiaggiati lungo il litorale messinese dello Stretto. Atti Soc. Pelorit. 19, 129–156 (1973).
Scotto Di Carlo, B., Costanzo, G., Fresi, E., Guglielmo, L. & Ianora, A. Feeding ecology and stranding mechanisms in two lanternfishes, Hygophum benoiti and Myctophum punctatum. Mar. Ecol. Prog. Ser 9, 13–24 (1982).
Battaglia, P., Ammendolia, G., Cavallaro, M., Consoli, P. & Esposito, V. Influence of lunar phases, winds and seasonality on the stranding of mesopelagic fish in the Strait of Messina (Central Mediterranean Sea). Mar. Ecol. 38, e12459. https://doi.org/10.1111/maec.12459 (2017).
Umgiesser, G., Canu, D. M., Cucco, A. & Solidoro, C. A finite element model for the Venice Lagoon. Development, set up, calibration and validation. J. Mar. Syst. 51, 123–145 (2004).
Ferrarin, C., Bergamasco, A., Umgiesser, G. & Cucco, A. Hydrodynamics and spatial zonation of the Capo Peloro coastal system (Sicily) through 3-D numerical modeling. J. Mar. Syst. 117, 96–107 (2013).
Umgiesser, G., Ferrarin, C., Cucco, A., De Pascalis, F. & Bellafiore, D. Comparative hydrodynamics of 10 Mediterranean lagoons by means of numerical modeling. J. Geophys. Res. Oceans 119, 2212–2226 (2014).
Cucco, A., Quattrocchi, G., Satta, A., Antognarelli, F. & De Biasio, F. Predictability of wind-induced sea surface transport in coastal areas. J. Geophys. Res. Oceans 121, 5847–5871. https://doi.org/10.1002/2016JC011643 (2016).
Cucco, A., Quattrocchi, G. & Zecchetto, S. The role of temporal resolution in modeling the wind induced sea surface transport in coastal seas. J. Mar. Syst. 193, 46–58. https://doi.org/10.1016/j.jmarsys.2019.01.004 (2019).
Quattrocchi, G. et al. An operational numerical system for oil stranding risk assessment in a high-density vessel traffic area. Front. Mar. Sci. 8, 585396. https://doi.org/10.3389/fmars.2021.585396 (2021).
Cucco, A. et al. A high-resolution real-time forecasting system for predicting the fate of oil spills in the Strait of Bonifacio (western Mediterranean Sea). Mar. Pollut. Bull. 64, 1186–1200 (2012).
Cucco, A. & Umgiesser, G. The Trapping Index: How to integrate the Eulerian and the Lagrangian approach for the computation of the transport time scales of semi-enclosed basins. Mar. Pollut. Bull. 98, 210–220 (2015).
Quattrocchi, G. et al. Optimal design of a Lagrangian observing system for hydrodynamic surveys. J. Oper. Oceanogr. 9(suppl.), s77–s88. https://doi.org/10.1080/1755876X.2015.1114805 (2016).
Quattrocchi, G. et al. Hydrodynamic controls on connectivity of the high commercial value shrimp Parapenaeus longirostris (Lucas, 1846) in the Mediterranean Sea. Sci. Rep. 9, 16935. https://doi.org/10.1038/s41598-019-53245-8 (2019).
Pastor-Prieto, M. et al. Spatial heterogeneity of Pelagia noctiluca ephyrae linked to water masses in the Western Mediterranean. PLoS ONE 16, e0249756. https://doi.org/10.1371/journal.pone.0249756 (2021).
Haeckel, E. Das system der medusen. Monographie der Medusen 499–510 (Gustav Fischer Verlag, 1880).
Avian, M. Temperature influence on in vitro reproduction and development of Pelagia noctiluca (Forsskäl, 1775). Boll. Zool. 53, 385–391 (1986).
Fossette, S. et al. Current-oriented swimming by jellyfish and its role in bloom maintenance. Curr. Biol. 25, 342–347. https://doi.org/10.1016/j.cub.2014.11.050 (2015).
Pinardi, N. et al. Mediterranean Sea large-scale low-frequency ocean variability and water mass formation rates from 1987 to 2007: A retrospective analysis. Prog. Oceanogr. 132, 318–332 (2015).
Demirov, E. & Pinardi, N. Simulation of the Mediterranean Sea circulation from 1979 to 1993: Part I. The interannual variability. J. Mar. Syst. 33–34, 23–50 (2002).
Menna, M. et al. New insights of the Sicily channel and Southern Tyrrhenian sea variability. Water 11, 1355 (2019).
Avian, M. & Rottini Sandrini, L. Oocyte development in four species of scyphomedusa in the northern Adriatic Sea. Hydrobiologia 216/217, 189–195 (1991).
Malej, A. Behaviour and trophic ecology of the jellyfish Pelagia noctiluca (Forsskäl, 1775). J. Exp. Mar. Biol. Ecol. 126, 259–270 (1989).
Lo Bianco, S. Notizie biologiche riguardanti specialmente il periodo di maturità sessuale degli animali del golfo di Napoli. Mitt. Zool. Stn. Neapel 19, 513–761 (1909).
Purcell, J. E., Malej, A. & Benović, A. Potential links of jellyfish to eutrophication and fisheries. In Coastal and Estuarine Studies, Ecosystem at the Land-Sea Margin Drainage Basin to Coastal Sea (eds Malone, T. C. et al.) 241–263 (American Geophysical Union, 1999).
Spezie, G. C., Sansone, E., Budillon, G. & Gallarato, A. Caratterizzazione idrodinamica del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia. Campagna oceanografica 1994. Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM94). In Data Rep., (eds. Faranda, F. M. & Povero, P.) 1–82 (1995).
Spezie, G. C. et al. Rilievi idrodinamici nel sistema Eolie e nei bacini limitrofi di Cefalù e Gioia. Campagna oceanografiche 1995. Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM95). In Data Rep. (eds. Faranda, F. M. & Povero, P.) 1–98 (1996).
Carrada, G. C., Ribera D’Alcalà, M. & Saggiomo, V. The pelagic system of the Southern Tyrrhenian Sea. Some comments and working hypotheses. In Proceedings IX Proceedings XII Italian Association of Oceanography and Limnology Congress 151–166 (1992).
Povero, P., Misic, C., Acconero, A. & Fabiano M. Distribuzione e caratterizzazione biochimica della sostanza organica particellata nelle acque del Tirreno Sud Orientale. In Acts 12 Congress of the Italian Association of Oceanology and Limnology 227–237 (1998).
Brancato, G., Minutoli, R., Granata, A., Sidoti, O. & Guglielmo L. Diversity and vertical migration of euphausiids across the Straits of Messina area. In: Mediterranean Ecosystem: Structures and Processes (eds. Faranda, F. M., Guglielmo, L. & Spezie, G.) 131–141 (Springer, 2001).
Sitran, R., Bergamasco, A., Decembrini, F. & Guglielmo, L. Microzooplankton (tintinnid ciliates) diversity: Coastal community structure and driving mechanisms in the Southern Tyrrhenian Sea (Western Mediterranean). J. Plankton Res. 31, 153–170 (2009).
Fonda Umani, S., Monti, M., Minutoli, R. & Guglielmo, L. Recent advances in the Mediterranean researches on zooplankton: from spatial–temporal patterns of distribution to processes oriented studies. Adv. Oceanogr. Limnol. 1, 295–356 (2010).
Giordano, D. et al. Summer larval fish assemblages in the Southern Tyrrhenian Sea (Western Mediterranean Sea). Mar. Ecol. 36, 104–117. https://doi.org/10.1111/maec.12123 (2015).
Fonda Umani, S., Milani, L. & Martecchini, E. Distribuzione dei popolamenti microzooplanctonici durante la campagna oceanografica Eolie 1994. Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM95). In Data Rep. (eds. Faranda, F. M. & Povero, P.) 199–222 (1995).
Carrada, G. C., Mangoni, O. & Sgrosso, S. Distribuzione spaziale di clorofilla a e di feopigmenti in diverse frazioni dimensionali del fitoplancton. Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM95). In Data Rep. (eds. Faranda, F. M. & Povero, P.) 197–216 (1996).
Guglielmo, L. et al. Distribuzione verticale e migrazione giornaliera dello zooplancton e del micronecton nel Tirreno meridionale (Isole Eolie). Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM95). In Data Rep. (eds. Faranda, F. M. & Povero, P.) 217–246 (1996).
Innamorati, M., Lazzara, L., Massi, L., Biondi, N. & Nuccio, C. Fitoplancton, luce e produzione primaria nella’Arcipelago delle Isole Eolie, in estate. Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM95). In Data Rep. (eds. Faranda, F. M. & Povero, P.) 161–196 (1996).
Zunini Sertorio, T., Licandro, P., Giallain, M. & Bernat, P. Distribuzione verticale della biomassa zooplanctonica su una stazione delle Isole Eolie (Luglio 1995). Caratterizzazione ambientale marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EUCUMM95). In Data Rep. (eds. Faranda, F. M. & Povero, P.) 247–254 (1996).
Sabates, A. et al. Pathways for Pelagia noctiluca jellyfish intrusions onto the Catalan shelf and their interactions with early life fish stages. J. Mar. Syst. 187, 52–61 (2018).
Mosetti, F. Currents in the Straits of Messina. In The Straits of Messina ecosystem (eds Guglielmo, L. et al.) 13–29 (University of Messina, Department of Marine Biology and Ecology, 1995).
Zavodnik, D. Spatial aggregations of the swarming jellyfish Pelagia noctiluca (Scyphozoa). Mar. Biol. 94, 265–269 (1987).
El Rahi, J., Weeber, M. P. & El Serafy, G. Modelling the effect of behavior on the distribution of the jellyfish Mauve stinger (Pelagia noctiluca) in the Balearic Sea using an individual-based model. Ecol. Model. 433, 109230 (2020).
Axiak, V. & Civili, F. S. Jellyfish blooms in the Mediterranean: causes, mechanisms, impact on man and the environment. A programme review. In: UNEP: Jellyfish blooms in the Mediterranean. Proceedings of the II Workshop on Jellyfish in the Mediterranean Sea. MAP Tech. Rep. Ser. Vol. 47, 1–10 (UNEP, 1991).
Keesing, J. K. et al. Role of winds and tides in timing of beach strandings, occurrence, and significance of swarms of the jellyfish Crambione mastigophora Mass 1903 (Scyphozoa: Rhizostomeae: Catostylidae) in north-western Australia. Hydrobiologia 768, 19–36. https://doi.org/10.1007/s10750-015-2525-5 (2016).
Aglieri, G. et al. First evidence of inbreeding, relatedness and chaotic genetic patchiness in the holoplanktonic jellyfish Pelagia noctiluca (Scyphozoa, Cnidaria). PLoS ONE 9, e99647. https://doi.org/10.1371/journal.pone.0099647 (2014).
Alpers, W., Brandt, P. & Rubino, A. Internal waves generated in the Strait of Gibraltar and Messina: Observations from space. In Remote Sensing of the European Seas (eds. Barale, V. & Gade, M.) 319–330 (Springer, 2008). https://doi.org/10.1007/978-1-4020-6772.
Droghei, R. et al. The role of Internal Solitary Waves on deep-water sedimentary processes: The case of up-slope migrating sediment waves off the Messina Strait. Sci. Rep. 6, 36376. https://doi.org/10.1038/srep36376 (2016).
La Forgia, G. et al. Sediment resuspension and bedform generation induced by internal solitary waves. Geophys. Res. Abs. Vol. 21, EGU2019-9121, EGU General Assembly (2019).
Lohmann, H. Die Stromunger in der Strasse von Messina und die verteilung des planktons in derselben. Int. Rev. Ges. Hydrobiol. 2, 505–556 (1909).
Magazzù, G. & Andreoli, C. Trasferimenti fitoplanctonici attraverso lo Stretto di Messina in relazione alle condizioni idrologiche. Boll. Pesca Piscic. Idrobiol. 26, 125–193 (1971).
Palanques, A. et al. General patterns of circulation, sediment fluxes and ecology of the Palamòs (La Fonera) submarine canyon, northwestern Mediterranean. Progr. Oceanogr. 66, 89–119 (2005).
Granata, A. et al. Vertical distribution and diel migration of zooplankton and micronekton in Polcevera submarine canyon of the Ligurian mesopelagic zone (NW Mediterranean Sea). Progr. Oceanogr. 183, 102298. https://doi.org/10.1016/j.pocean (2020).
Zagami, G. et al. Spring copepod vertical zonation pattern and diel migration in the open Ligurian Sea (north-western Mediterranean). Progr. Oceanogr. 183, 102297. https://doi.org/10.1016/j.pocean (2020).
Danovaro, R. & Boero, F. Italian seas. In: World Seas: An Environmental Evaluation. Vol. I Europe, The Americas and West Africa. (ed. Sheppard, C.) 283–306 (Elsevier Ltd., 2019). https://doi.org/10.1016/B978-0-12-805068-2.00044-9
Lo Iacono, C., Sulli, A. & Agate, M. Submarine canyons of north-western Sicily (Southern Tyrrhenian Sea): Variability in morphology, sedimentary processes and evolution on a tectonically active margin. Deep-Sea Res. 104, 93–105 (2014).
Acknowledgements
The research leading to these results has received funding from the European Community’s Seventh Framework Programme (FP7/2007–2013) under Grant Agreements n° 266445 for the project “Vectors of Change in Oceans and Seas Marine Life, Impact on Economic Sectors (VECTORS),” n°287844 for the project “Towards COast to Coast NETworks of marine protected areas (from the shore to the high and deep sea), coupled with sea-based wind energy potential” (CoCoNET). Thanks go both to Nino and Giuseppe Donato for giving informations during visual observations in coastal and open waters of the study areas and to Dario Lopes for giving information, pictures and videos of Pelagia blooms near Aeolian archipelago. A.M. and M.V. acknowledge the financial support from the Slovenian Research Agency (research core funding No. P1-0237 “Coastal Sea Research” and project Z7‐1884).
Author information
Authors and Affiliations
Contributions
A.B.: Conceptualization, Methodology, Formal analysis, Writing Original draft; A.C.: Methodology, Numerical modeling, Formal analysis, Writing Original draft; L.G.: Conceptualization, Writing Original draft, Supervision, Funding acquisition; R.M.: Investigation, Validation; G.Q.: Methodology, Numerical modeling, Formal analysis; R.G.: Investigation, Validation; F.P.: Visualization; M.P.: Investigation, Fieldwork Designing; G.Z.: Investigation, Validation; M.V.: Writing Review and Editing; A.M.: Writing Review and Editing; A.G.: Conceptualization, Methodology, Supervision.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bergamasco, A., Cucco, A., Guglielmo, L. et al. Observing and modeling long-term persistence of P. noctiluca in coupled complementary marine systems (Southern Tyrrhenian Sea and Messina Strait). Sci Rep 12, 14905 (2022). https://doi.org/10.1038/s41598-022-18832-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-18832-2
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
