
Abstract
Salpa thompsoni is an important grazer in the Southern Ocean. Their abundance in the western Antarctic Peninsula is highly variable, varying by up to 5000-fold inter-annually. Here, we use a particle-tracking model to simulate the potential dispersal of salp populations from a source location in the Antarctic Circumpolar Current (ACC) to the Palmer Long Term Ecological Research (PAL LTER) study area. Tracking simulations are run from 1998 to 2015, and compared against both a stationary salp population model simulated at the PAL LTER study area and observations from the PAL LTER program. The tracking simulation was able to recreate closely the long-term trend and the higher abundances at the slope stations. The higher abundances observed at slope stations are likely due to the advection of salp populations from a source location in the ACC, highlighting the significant role of water mass circulation in the distribution and abundance of Southern Ocean salp populations.
Similar content being viewed by others


Introduction
The salp, Salpa thompsoni, is an important grazer in the Southern Ocean1, with a wide distribution spanning from the Subtropical Convergence to the Antarctic continent2. Hence, S. thompsoni can exist over a broad thermal range (− 1.5 to 7 °C), yet have a preference to warmer waters (2–5 °C)2 because cooler temperatures reduce their reproductive fitness3. As a result, it is more difficult for S. thompsoni populations to complete their lifecycle in cooler, high latitude waters, which explains low and highly variable abundances of S. thompsoni close to the Antarctic continent3,4,5,6,7.
The western Antarctic Peninsula (WAP) is a productive region with high biomasses of Antarctic krill8 supporting numerous top predators, including penguins, seals and whales, as well as commercial fisheries9,10. It is one of the fastest warming regions in the world, resulting in reduced sea ice extent and duration11,12,13, and in the past two decades has seen increases in S. thompsoni abundance14,15. Salps are efficient filter feeders and have a grazing pressure that can regionally exceed the total daily primary production16. Hence, their presence can increase competitive interactions with other zooplankton, with regional decreases in Euphausia superba (hereafter Antarctic krill) abundance and increases in S. thompsoni14. In the WAP, abundances of S. thompsoni are generally low (climatological mean of ~ 0.1 individuals (ind.) m−3) yet can vary inter-annually by a factor of 50007. Large scale atmospheric processes, in addition to biological processes (i.e., mortality, recruitment, survival), were shown to explain the large variability observed in Antarctic krill density and demography17. Thus, the large variability in salp density in the WAP may also be driven by these processes that influence sea ice extent and water mass circulation18 linking long-term S. thompsoni abundances to large atmospheric processes such as the Southern Annular Mode (SAM)15. Increasing SAM has been associated with earlier sea ice retreat, later sea ice advance and increased upwelling of warm circumpolar deep water (CDW) in the WAP18.
Because S. thompsoni reproduction is physiologically limited by cold water3, it has been hypothesized that the high biomass years, hereafter “salp years”, are a result of warmer water intrusions onto the shelf7,19. These intrusions of warmer water could increase local salp abundance in two ways, either by making the environmental conditions more conducive for the salp reproduction, or by the influx of seeding populations occurring from more optimal source locations. Favourable environmental conditions may not persist throughout the winter affecting the reproductive success of S. thompsoni5. Hence, the “source population” hypothesis is more plausible with recent modeling efforts in the WAP indicating that immigration of S. thompsoni individuals is critical to recreate the observed large variations in abundance20. While the study by Groeneveld et al.20 is an individual-based local salp model that identified that large salp blooms could not occur without a population boost in early spring, it did not model the likelihood of salp populations being transported from a source location in the Southern Ocean into the WAP. This study builds upon the static approach by Groeneveld et al.20 by incorporating the effect of water mass circulation on the dispersal and growth of salp populations.
Particle tracking and ocean models are used to explore the connectivity among oceanic systems and improve, (1) the understanding of the plankton dispersal, (2) the identification of spawning grounds and nurseries21,22 and/or (3) high-risk areas for exotic species invasions23. In the WAP, annual sampling of S. thompsoni have been undertaken in the austral summer (January–February) as part of the Palmer Long Term Ecological Research (PAL LTER) program along the Antarctic Peninsula. In this study, we aim to test the “source population” hypothesis by using the results from the ICHTHYOP particle tracking model24 fed with 3D velocity fields combined with a S. thompsoni population model25 to uncover the physical and ecological processes driving the S. thompsoni population distribution and abundance at the PAL LTER study region. Specifically, we use modeled velocities to virtually transport salp populations from a potential source location in the ACC, southeast of the PAL LTER study region. While being transported, the salp population experiences the changes in the water parcel’s temperature and chlorophyll a concentration, which influences salp growth, mortality and reproductive rates. Given that simulations were run from 1998 to 2015, this study provides a framework to explore how water mass circulation affects S. thompsoni distribution dynamics in the WAP, and in particular to determine how much advection may contribute to the large inter-annual variability observed in S. thompsoni populations.
Results
Environmental conditions
Long-term (1998–2015) mean summer (Dec–Feb) modelled sea surface temperature (SST) at the source location was significantly higher than slope and shelf stations in the PAL LTER study region (F2,162 = 53, p < 0.001; Fig. 1). Summer (Dec–Feb) modelled chlorophyll a concentrations were significantly higher in the shelf stations of the PAL LTER study region compared to the source location (F2,162 = 3.39, p = 0.04). There was no significant difference in SST or chlorophyll a between shelf and slope stations. While chlorophyll a is a necessary input into the S. thompsoni model, advection proves to be a more significant driver of variation as seen in the results below.

Long-term (1998–2015) mean summer (Dec–Feb) modelled (a) SST and (b) chlorophyll a at the source, slope and shelf regions. Letters denote significant differences at p < 0.05.
Dispersal patterns
The movement of simulated S. thompsoni populations were generally consistent with the Antarctic Circumpolar Current, travelling north-east, parallel to the Antarctic Peninsula. The proportion of particles reaching the PAL LTER study region during the sampling period varied each year, ranging from 0.002 (2005) to 0.06 (2008; Fig. 2). The majority (99.5 ± 1.6%) of particles that reached the PAL LTER study region remained on the slope, with particles only reaching the shelf in 5 of the 18 simulated years (2008–2011, 2013).

Proportion of particles entering sampling grid during sampling period. Particles only reached the shelf between 2008 to 2011 and in 2013.
PAL LTER observations
The long-term (1998–2015) mean (± SD) S. thompsoni abundance observed at the PAL LTER study region is 0.07 ± 0.11 ind. m−3. Inter-annual variation in S. thompsoni abundance at shelf stations correlated significantly with the inter-annual variation from the whole PAL LTER study region (r = 0.98, p < 0.001), whereas there was no correlation between slope stations and the whole study region (Fig. 3). Long-term mean abundances were significantly higher in slope stations compared to shelf stations (F1,1128 = 15, p < 0.001; Fig. 4). The long-term trend indicates a period of low salp abundance from 1998 to 2008 and a period of high abundance from 2009 to 2013 (Fig. 5).

Annual mean salp abundance (salp index) for the PAL LTER study region (overall; solid line), shelf stations (grey line) and slope stations (dashed line).

Long-term (1998–2015) mean (± SD) observed and modeled Salpa thompsoni abundance in slope and shelf stations of the PAL LTER study region. *significant differences at p < 0.05.

Long-term trends (5-yr mean) in observed and modelled Salpa thompsoni abundance at the PAL LTER study region.
Simulated salp abundance
The long-term (1998–2015) mean (± SD) S. thompsoni abundance simulated by the stationary model (0.14 ± 0.03 ind. m−3) and the tracking model (0.19 ± 0.05 ind. m−3) were not significantly different from the observations. However, the stationary model could not recreate the higher abundance in slope stations (Fig. 4), or the long-term trend (Fig. 5). In the tracking model, simulated abundances were significantly higher in the slope than in the shelf (F1,35 = 30.75, p < 0.001; Fig. 4) and the long-term trend was consistent with the observations (r = 0.75, p = 0.002; Fig. 5), indicating a similar high period from 2009 to 2013. Higher abundances in the tracking model were driven by the number of particles reaching the sampling area (r = 0.78, p < 0.001).
Discussion
This is the first study to explore the effect of water mass advection on the population density dynamics of S. thompsoni populations in the WAP. S. thompsoni abundance is highly variable along the WAP, with “salp years” potentially shifting the local ecosystem due to their efficient grazing rates and ability to alter the ecosystem C:N:P stoichiometry5,26. Since limited seasonal sampling restricts determination of environmental drivers of inter-annual S. thompsoni abundance variability in the WAP, we compared two simulations of a salp population model at the PAL LTER region. A stationary model run precluded the immigration of new individuals while a tracking model through inferred modeled velocities was able to transport S. thompsoni populations from a source location in the ACC.
Long-term observations revealed high inter-annual variance in abundance, 20-fold higher abundances in slope stations and a high period of abundance from 2009 to 2013. Salps are historically found in low abundances in the study region, particularly in shelf stations (0.01 ind. m−3)7,15. Periods of high salp abundance have been related to large scale atmospheric processes, positively correlated with the Southern Annular Mode (SAM) and negatively correlated with the Multivariate El Niño Southern Oscillation Index (MEI)15. In the WAP, positive SAM and negative MEI years result in increased northwesterly winds, unfavourable conditions for sea ice and potentially warmer surface waters15. Local salp populations in the PAL LTER study region can benefit from warmer surface waters, however, if the increased northwesterly winds alter surface circulation, these conditions could also result in more ACC encroachment into the PAL LTER study region, allowing for increased immigration of new salps15.
Both the stationary and tracking models were able to recreate the long-term mean abundance observed for the entire (shelf and slope) PAL LTER study region. This indicates that the model is sufficiently robust to mimic the S. thompsoni population in the PAL LTER region by using only temperature and chlorophyll a as drivers. However, higher abundances in slope stations were only confirmed when the immigration of new individuals was allowed through the tracking simulations. Subsequently, only the tracking simulations were able to recreate the regional long-term trend of S. thompsoni densities. We note, however, that while the 1/4° resolution physical-biogeochemical MOM6-COBALT resolves large-scale spatial and seasonal chlorophyll patterns around the WAP reasonably well, it cannot capture small-scale circulation and frontal features.
The stationary simulation could not recreate the variation in abundance between slope and shelf stations suggesting that thermal and biogeochemical conditions alone were not sufficient to result in consistently higher S. thompsoni abundances over the slope. Indeed, SST and chlorophyll a concentrations were not significantly different between shelf and slope stations. At the same time, the source location was significantly warmer, and had lower chlorophyll a concentration. S. thompsoni abundances generally increased with increasing temperature and decreasing chlorophyll a concentrations3,25, indicating that the source location could be a more optimal habitat for S. thompsoni populations to thrive. The observed long-term trend in S. thompsoni abundance was highly correlated with the number of particles arriving in the PAL LTER study region, and these particles rarely reached the shelf stations. Thus, if populations at the source location were able to develop more successfully and then were advected onto the slope, this could explain the higher salp abundances observed at the slope stations as well as the large inter-annual variation in the S. thompsoni abundance.
The agreement between the number of particles reaching the slope stations, and the higher abundance observed in slope stations highlights the significant role of circulation in Southern Ocean salp populations, and provides a strong support for the “source population” hypothesis. While there is uncertainty surrounding the frequency and magnitude of the advection of S. thompsoni populations into the PAL LTER study region, this study postulates that the immigration of new populations is necessary to recreate the observed shelf-slope gradients and long-term trends in salp abundance. This study highlights the critical importance of understanding the circum-Antarctic connectivity of the S. thompsoni metapopulation in the Southern Ocean and the significance of regional specifics in salp population developments. Identifying if there are genetic links between areas will be useful in determining how connected the S. thompsoni populations are in the Southern Ocean.
In the long-term, as the ACC moves poleward and closer to the shelf break27,28,29, the advection of S. thompsoni into the PAL LTER study region is likely to increase in the future. While temperature and chlorophyll a are important drivers of salp distribution and abundance, in order to understand the large-scale variations in inter-annual salp abundance, it is necessary to consider the effect of water mass advection and identify the source location of potential seeding populations. A critical reason to increase our understanding of these impacts are to understand the effect that shifting salp populations could have on the food web dynamics in the region30,31,32.
Methods
Study site and observations
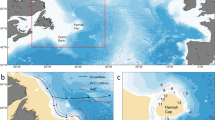
The PAL LTER sampling stations are located on the western side of the Antarctic Peninsula. The sampling region extends southwards from Anvers Island (~ 63 °S) to Charcot Island (~ 69° S; Fig. 6). Grid lines are 100 km apart, and extend perpendicular from the coast, extending approximately 200 km offshore to slope waters. Stations along grid lines are spaced approximately 20 km apart and are separated as shelf (0–750 m) or slope (> 750 m). Zooplankton sampling was undertaken annually (1998–2015) during the austral summer (Jan–Feb; Table S1) using a 1 m2 (335 μm Nytex mesh) and a 2 m2 (700 μm knotless nylon square mesh) Metro net towed obliquely from the surface to 300 m and 120 m respectively. For more detail on the study site and sampling method see Ross et al.7. These data are sourced from the PAL LTER data catalog (https://pal.lternet.edu/data).

Sampling area indicating the PAL LTER study region and the locations of particles for the 1998 forward simulation. Particle locations during the simulation are represented by the coloured points: the initial release site (i.e. source location; day 0; grey points), day 50 (blue points), day 100 (red points) and the final location (day 151; green). Approximate Salpa thompsoni sampling stations in the PAL LTER study region are represented by black (shelf) and white (slope) circles. The southern boundary of the Antarctic Circumpolar Current is indicated by the dashed line. Figure was created using MATLAB version 9.7 (R2019b), Natick, Massachusetts: The MathWorks Inc.; 2019.
Particle tracking
The velocity fields were extracted from the GLORYS12V1 product distributed by the Copernicus platform (marine.copernicus.eu). The GLORYS12V1 product (https://doi.org/10.48670/moi-00024) is the CMEMS global ocean eddy-resolving reanalysis covering the altimetry era 1993–201833,34. We used daily mean files of currents (U, V and W) from the surface to the 1000 m depth, displayed on a standard regular grid at 1/12° (approximately 8 km) and on 35 standard levels. It is based largely on the current real-time global forecasting CMEMS system. The model component is the NEMO platform driven at the surface by ECfMWF ERA-Interim reanalysis. Observations are assimilated by means of a reduced-order Kalman filter. Along track altimeter data (Sea Level Anomaly), satellite Sea Surface Temperature, Sea Ice Concentration and in situ temperature and salinity vertical profiles are jointly assimilated.
Particle trajectories were simulated using ICHTHYOP (www.ichthyop.org), an individual-based model designed to study how physical and biological factors affect plankton dynamics24. Daily velocity fields from GLORYS12V1 were used to compute Lagrangian trajectories with the forward Runge–Kutta method (RK4). The computational time step was set to 1800s, ensuring that a particle does not move longer than one grid cell per time step.
A patch consisting of 3000 particles was released into a circular domain with a 200 km diameter at the source location (67° S and 90.5° W; Fig. 6). Horizontal diffusion was implemented according to Peliz et al.35 where the Lagrangian horizontal diffusion (Kh) is defined as:
where ε is the turbulent dissipation rate and l the grid cell size (8 km in GLORYS12v1). The turbulent dissipation rate has been set to 10–10 resulting in Kh equal to 75 m2 s−1, which is in accordance with other studies36,37. The particles were released the October 1st of each year (1997–2014) and the trajectories were simulated for 151 days until the end of February. Salp diel vertical migration (DVM) was included with particles close to the surface (50 m depth) during the night and distributed deeper in the water column (200 m) during the day, assuming sunrise and sunset time of 6 am and 9 pm, respectively.
The source location and release time (Fig. 6) was defined by backtracking particles from the PAL LTER study region for several years (Fig. S1). Several sources were considered. Most of the particles originated from the surface layer (0–300 m) upstream the ACC while another regular source was located directly offshore the study area, with particles slowly advected onto the shelf from deeper layers. The former source was found to better reproduce observed patterns and was therefore selected. The distribution of particles from the selected source location resulted in a significant portion of particles being transported onto the shelf with their position on the shelf agreeing with locations of S. thompsoni documented in the literature7,15. An October 1st release date was chosen as it took approximately 3–4 months for particles to reach an ACC source location south of the PAL LTER study region, and because S. thompsoni reproduction begins in spring2.
Virtual salp populations
Each particle is assumed to be a 1 m3 parcel of water, containing a S. thompsoni population. The salp population is simulated by the model developed by Henschke et al.25. This is a size-structured S. thompsoni population model that incorporates temperature and chlorophyll a dependent growth, consumption, reproduction and mortality25.
S. thompsoni population dynamics are forced with sea surface temperature (SST) and chlorophyll a extracted from a ¼° horizontal resolution global ocean simulation from the Modular Ocean Model 6 (MOM6)38 that included the Carbon Ocean Biogeochemistry and Lower Trophics (COBALT) plankton ecosystem model39,40. COBALT was used as it contains both SST and chlorophyll a values as opposed to GLORYS12V1 from where the particle tracking fields were derived which does not contain chlorophyll a. Modelled SST and chlorophyll a data were used to ensure the most robust coverage in the area, as satellite data from the Southern Ocean are limited due to the region often being obscured by clouds. The COBALT model has proven to be effective in recreating long-term patterns in SST (r = 0.97, p < 0.001) and chlorophyll a (r = 0.71, p < 0.01) globally when compared against in situ observations and satellite data (SST and chlorophyll a) (Table S2). The COBALT model could also recreate the monthly (r = 0.59, p < 0.04, n = 12) and inter-annual (1996–2007; r = 0.66, p = 0.02, n = 12) trends observed from in situ samples of chlorophyll a from the Palmer Station seawater intake (1992–2018). SST trends in COBALT correspond well with large scale monthly SST (r = 0.94, p < 0.001, n = 240) trends from GLORYS12V1 in the WAP.
To compare the effect of advection, two simulations are performed: a stationary simulation and a tracking simulation. Both simulations are run using a 5-year model spin-up prior to commencing to ensure the salp population has reached steady state using the local climatological mean values for SST and chlorophyll a. The stationary simulation is run for the sampling period (1998–2015) across grid cells in the PAL LTER study region. The tracking simulations begin at the source location (Fig. 6) where the 5-year spin up is run, ending by October 1. The particles begin within a circle, and after October 1 until the end of the simulation (February 28), as the particles are moved their temperature and chlorophyll a concentration trajectory will vary from other particles based on their movement pattern. Each year of the tracking simulation is run separately, so populations will not cumulatively increase throughout the years as advection continues. The S. thompsoni life cycle can be completed within a year, hence accumulation is not likely25. Inter-annual abundances are calculated based on the mean abundance within the PAL LTER study region from the duration of the sampling period (Table S1). Inter-annual abundances for the tracking simulation are the combined abundances of the local salp populations (stationary simulation) and the influx of additional salps (tracking simulation). The sampling period occurred during January/February (Table S1), and as the tracking simulation was still running until February 28th, this ensured that particles were able to be advected out of the study region as well. Modelled and observed salp abundances were both standardised to an annual mean of 0 and a standard deviation of 1 to create a salp index. A similar method was used to compare jellyfish abundances across diverse metrics41. An analysis of variance was used to compare differences in environmental conditions or salp abundances between regions (source, shelf, slope). Pearson correlations were used to compare interannual variation in abundances between regions.
Data availability
The zooplankton dataset was obtained from the Palmer LTER data catalog: Palmer Station Antarctica LTER, D. Steinberg, R. Ross, and L. Quetin. 2020. Zooplankton collected aboard Palmer Station LTER annual cruises off the western antarctic peninsula, 1993–2008. ver 4. Environmental Data Initiative. https://doi.org/10.6073/pasta/03e6d72a78bc2512ef5bb327e686f8fa (Accessed 2023-02-09). Additional datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Voronina, N. M. Comparative abundance and distribution of major filter-feeders in the Antarctic pelagic zone. J. Mar. Syst. 17, 375–390 (1998).
Foxton, P. The distribution and life-history of Salpa thompsoni Foxton with observations on a related species, Salpa gerlachei Foxton. Discov. Rep. 34, 1–116 (1966).
Henschke, N. & Pakhomov, E. A. Latitudinal variations in Salpa thompsoni reproductive fitness. Limnol. Oceanogr. 64, 575–584 (2018).
Chiba, S., Ishimaru, T., Hosie, G. W. & Wright, S. W. Population structure change of Salpa thompsoni from austral mid-summer to autumn. Polar Biol. 22, 341–349 (1999).
Pakhomov, E. A., Froneman, P. W. & Perissinotto, R. Salp/krill interactions in the Southern Ocean: Spatial segregation and implications for the carbon flux. Deep Sea Res. Part II 49, 1881–1907 (2002).
Ono, A., Ishimaru, T. & Tanaka, Y. Distribution and population structure of salps off Adelie Land in the Southern Ocean during austral summer, 2003 and 2005. La mer 48, 55–70 (2010).
Ross, R. M. et al. Trends, cycles, interannual variability for three pelagic species west of the Antarctic Peninsula 1993–2008. Mar. Ecol. Prog. Ser. 515, 11–32 (2014).
Atkinson, A., Siegel, V., Pakhomov, E. A., Jessop, M. J. & Loeb, V. J. A re-appraisal of the total biomass and annual production of Antarctic krill. Deep Sea Res. Part I 56, 727–740 (2009).
Smetacek, V. & Nicol, S. Polar ocean ecosystems in a changing world. Nature 437, 362–368 (2005).
Nowacek, D. P. et al. Super-aggregations of krill and humpback whales in Wilhelmina Bay, Antarctic Peninsula. PLoS ONE 6, e19173 (2011).
Parkinson, C. L. Trends in the length of the Southern Ocean sea-ice season, 1979–99. Ann. Glaciol. 34, 435–440 (2002).
Stammerjohn, S. E., Massom, R., Rind, D. & Martinson, D. Regions of rapid sea ice change: An inter-hemispheric seasonal comparison. Geophys. Res. Lett. 39, L06501 (2012).
Henley, S. F. et al. Variability and change in the west Antarctic Peninsula marine system: Research priorities and opportunities. Prog. Oceanogr. 173, 208–237 (2019).
Atkinson, A., Siegel, V., Pakhomov, E. & Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 432, 100–103 (2004).
Steinberg, D. K. et al. Long-term (1993–2013) changes in macrozooplankton off the Western Antarctic Peninsula. Deep Sea Res. Part I(101), 54–70 (2015).
Dubischar, C. D. & Bathmann, U. V. Grazing impact of copepods and salps on phytoplankton in the Atlantic sector of the Southern Ocean. Deep-Sea Res. Part II Top. Stud. Oceanogr. 44, 415–433 (1997).
Saba, G. K. et al. Winter and spring controls on the summer food web of the coastal West Antarctic Peninsula. Nat. Commun. 5, 4318 (2014).
Stammerjohn, S. E., Martinson, D. G., Smith, R. C., Yuan, X. & Rind, D. Trends in Antarctic annual sea ice retreat and advance and their relation to El Niño and Southern Annular Mode variability. J. Geophys. Res. 113, CS0S90 (2008).
Pakhomov, E. A., Hall, J., Williams, M. J. M., Hunt, B. P. V. & Stevens, C. J. Biology of Salpa thompsoni in waters adjacent to the Ross Sea, Southern Ocean, during austral summer 2008. Polar Biol. 34, 257–271 (2011).
Groeneveld, J. et al. Blooms of a key grazer in the Southern Ocean—an individual based model of Salpa thompsoni. Prog. Oceanogr. 185, 56 (2020).
Cetina-Heredia, P., Roughan, M., Van Sebille, E., Feng, M. & Coleman, M. A. Strengthened currents override the effect of warming on lobster larval dispersal and survival. Glob. Change Biol. 21, 4377–4386 (2015).
Everett, J. D. et al. Dispersal of Eastern King Prawn larvae in a western boundary current: New insights from particle tracking. Fish. Oceanogr. 26, 513–525 (2017).
Jaspers, C. et al. Ocean current connectivity propelling the secondary spread of a marine invasive comb jelly across western Eurasia. Global Ecol. Biogeogr. 27, 814–827 (2018).
Lett, C. et al. A Lagrangian tool for modelling ichthyoplankton dynamics. Environ. Model. Softw. 23, 1210–1214 (2008).
Henschke, N., Pakhomov, E. A., Groeneveld, J. & Meyer, B. Modelling the life cycle of Salpa thompsoni. Ecol. Model. 387, 17–26 (2018).
Alcaraz, M. et al. Changes in the C, N and P cycles by the predicted salps-krill shift in the Southern Ocean. Front. Mar. Sci. 1, 1–13 (2014).
Marshall, G. J. Trends in the southern annular mode from observations and reanalyses. J. Clim. 16, 4134–4143 (2003).
Böning, C. W., Dispert, A., Visbeck, M., Rintoul, S. R. & Schwarzkopf, F. U. The response of the Antarctic circumpolar current to recent climate change. Nat. Geosci. 1, 864–869 (2008).
Schofield, O. M. E. et al. How do polar marine ecosystems respond to rapid climate change. Science 328, 1520–1523 (2010).
Plum, C., Hillebrand, H. & Moorthi, S. Krill vs salps: Dominance shift form krill to salps as associated with higher dissolved N: P ratios. Sci. Rep. 10, 5911. https://doi.org/10.1038/s41598-020-62829-8 (2020).
Pauli, N.-C. et al. Selective feeding in Southern Ocean key grazers—diet compositon of krill and salps. Commun. Biol. 4, 1061. https://doi.org/10.1038/s42003-021-02581-5 (2021).
Pauli, N.-C. et al. Krill and salp faecal pellets contribute equally to the carbon flux at the Antarctic Peninsula. Nature Commun. 12, 7168. https://doi.org/10.1038/s41467-021-27436-9 (2021).
Lellouche, J. M. et al. Recent updates to the Copernicus Marine Service global ocean monitoring and forecasting real-time 1/12º high-resolution system. Ocean Sci. 14, 1093–1126 (2018).
Lellouche, N. et al. The Copernicus Global 1/12° Oceanic and Sea Ice GLORYS12 Reanalysis. Front. Earth Sci. 9, 25 (2021).
Peliz, A. et al. A study of crab larvae dispersal on the Western Iberian Shelf: Physical processes. J. Mar. Syst. 68, 215–236 (2007).
Espinasse, B. et al. Mechanisms regulating inter-annual variability in zooplankton advection over the Lofoten shelf, implications for cod larvae survival. Fish. Oceanogr. 26, 299–315 (2017).
Pepin, P., Han, G. & Head, E. J. Modelling the dispersal of Calanus finmarchicus on the Newfoundland Shelf: Implications for the analysis of population dynamics from a high frequency monitoring site. Fish. Oceanogr. 22, 371–387 (2013).
Adcroft, A. et al. The GFDL global ocean and sea ice model OM4.0: Model description and simulation features. J. Adv. Model. Earth Syst. https://doi.org/10.1029/2019ms001726 (2019).
Stock, C. A., Dunne, J. P. & John, J. G. Global-scale carbon and energy flows through the marine planktonic food web: An analysis with a coupled physical-biological model. Prog. Oceanogr. 120, 1–28 (2014).
Liu, X. et al. Simulated global coastal ecosystem responses to a half-century increase in river nitrogen loads. Geophys. Res. Lett. 48, 25 (2021).
Condon, R. H. et al. Recurrent jellyfish blooms are a consequence of global oscillations. Proc. Natl. Acad. Sci. U. S. A. 110, 1000–1005 (2013).
Author information
Authors and Affiliations
Contributions
This study was conceived by N.H. and E.A.P.; N.H., B.E. and E.A.P. designed the study; N.H. and B.E. ran the simulations; C.A.S., X.L. and N.B. provided model runs, data and assistance; N.H. and B.E. wrote the manuscript with comments from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Henschke, N., Espinasse, B., Stock, C.A. et al. The role of water mass advection in staging of the Southern Ocean Salpa thompsoni populations. Sci Rep 13, 7088 (2023). https://doi.org/10.1038/s41598-023-34231-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-34231-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
