
Abstract
To evaluate the influence that parasites have on the losses of Apis mellifera it is essential to monitor their presence in the colonies over time. Here we analysed the occurrence of nosematids, trypanosomatids and neogregarines in five homogeneous colonies for up to 21 months until they collapsed. The study, which combined the use of several molecular markers with the application of a massive parallel sequencing technology, provided valuable insights into the epidemiology of these parasites: (I) it enabled the detection of parasite species rarely reported in honeybees (Nosema thomsoni, Crithidia bombi, Crithidia acanthocephali) and the identification of two novel taxa; (II) it revealed the existence of a high rate of co-infections (80% of the samples harboured more than one parasite species); (III) it uncovered an identical pattern of seasonal variation for nosematids and trypanosomatids, that was different from that of neogregarines; (IV) it showed that there were no significant differences in the fraction of positive samples, nor in the levels of species diversity, between interior and exterior bees; and (V) it unveiled that the variation in the number of parasite species was not directly linked with the failure of the colonies.
Similar content being viewed by others


Introduction
The activity of the European or Western honeybee, Apis mellifera, is crucial for ensuring food production and maintaining the biodiversity of the ecosystems worldwide1. During the last decades, regional losses of honeybee colonies have been frequently reported2,3,4,5, although at a global scale the number of managed colonies seem to have risen, either due to their high demand for honey production and pollination services in certain areas6,7 or to the lack of adequate tools to quantify their mortality on a harmonized basis8.
The long-term declines observed in local stocks of managed honeybees is usually attributed to politic and socio-economic factors7,9, whereas annual losses (which are often amended by beekeepers replacing lost colonies) respond to a wider spectrum of causes4,5,8,9. These are multiple and interlinked, and go from degradation of the ecosystems due to global warming (e.g. weather events) and changes in land use (e.g. urbanisation, intensive agriculture), to health issues (e.g. stress due to beekeeping management, pollution entailed by the use of agrochemicals or dispersal of native and exotic parasites)1,2,8,9,10,11,12,13,14,15,16,17,18,19,20.
Given that the environment provides an optimal scenario for parasite transmission among social insects15,21,22,23,24, and taking into account that the fate of the colonies may depend on the timing and sequence of infections25, it is important to survey if the occurrence and diversity of these agents vary over time26,27,28,29,30,31.
Nowadays, the most comprehensive and cost-effective mode of assessing parasite diversity is to apply massive parallel sequencing, which allows the simultaneous identification of multiple pathogens across large numbers of samples, overcoming the need for cloning prior to Sanger sequencing, which is expensive and time-consuming32. Following this approach, we analysed the diversity of three major groups of unicellular parasites (nosematids, trypanosomatids and neogregarines) in five homogeneous colonies that were screened sequentially until their death.
Results and Discussion
Here we describe the results of a longitudinal study of the presence of parasites of the groups Nosematidae, Trypanosomatidae and Neogregarinorida in five homogenous colonies, which were monitored since the last trimester of 2014 until their collapse (the latest in August 2016).
Molecular detection of parasites
PCRs with specific primers allowed the detection and molecular identification of parasites in 76 of the 80 pooled DNA samples (95%; Table 1 and Supplementary Table S1). Nosematids were the most prevalent group (76.3%), followed by trypanosomatids and neogregarines (72.5% and 33.8%, respectively).
The primers targeting the Actin and RPB1 (RNA polymerase II large subunit RPB1) loci of nosematids and trypanosomatids, respectively, displayed higher sensitivity (96.7% and 84.5% of the positive samples) than the ones designed for the SSU (small-subunit ribosomal DNA) loci (78.7% and 55.2%, respectively; Supplementary Table S1). The rate of coincidence between the two markers was 75.4% for nosematids (i.e. both primers sets produced a band of the expected size in 46 out of 61 of the positive samples) and substantially lower for trypanosomatids (39.7%). This means that the PCR approach systematically misses a fraction of the positive samples, so that the results might underestimate the true parasite prevalence. To minimize this effect, in this study the presence of a parasite was determined by a positive result with either marker in a given sample.
The choice of primers had an effect on the sensitivity but also on the variety of species detected. Indeed, the pair targeting the Actin locus allowed the detection of Nosema ceranae and Nosema thomsoni, whereas the one targeting the SSU only permitted the identification of N. ceranae (Supplementary Table S1). Similarly, the primers used to amplify RPB1 revealed the presence of Lotmaria passim, Crithidia mellificae, Crithidia bombi, Crithidia acanthocephali and a new undescribed taxon (Trypanosomatidae sp.), while those amplifying the SSU only allowed the identification of L. passim and C. bombi. This lower performance of SSU is rather surprising as the amplification of ribosomal genes is one of the gold standards for molecular barcoding due to their sequence conservation and large number of copies. Nevertheless, the amplification success of a priori universal primers might be influenced by multiple factors like the GC-content within the amplified region, the number of mismatches between species33, or even the targeted species34, as the presence of previously undetected polymorphisms within the priming sites could drastically reduce their efficiency. These results emphasize the convenience of using a multilocus approach in order to produce a more accurate description of the diversity patterns of honeybee parasites, as many of them would have gone otherwise unnoticed.
Parasite diversity
N. ceranae was the most frequent pathogen of the series and was found in all Nosema positive samples. This is in line with previous epidemiologic analyses carried out in the same area31,35, whereas N. thomsoni was detected along with N. ceranae in two isolates (PR-05 I and PR-21 F; Supplementary Table S1). N. thomsoni was first described infecting a moth, Choristoneura conflictana36, and has now been identified in many other insect species, including bumblebees, beet webworms, ladybirds and solitary bees37,38,39,40,41. To the best of our knowledge, this is the first time it is reported in honeybees, although its presence in these pollinators might not be necessarily associated with tissue infection, as shown by Pereira et al.42 in bumblebees.
It must be highlighted that none of the samples tested positive for other nosematids previously detected in A. mellifera, like Nosema apis43 or Nosema neumanni44. The only known sequence of the latter taxon corresponds to the SSU locus, and it contains a mismatch in the region complementary to the forward primer used here that might have hindered its amplification.
Trypanosomatids displayed the highest species diversity, with an average of 2.1 ± 0.32 species per sample (Supplementary Table S1). In agreement with prior studies35,45,46,47, L. passim was the most prevalent trypanosomatid species of the dataset (68.8%; Table 1), closely followed by C. mellificae (61.3%) and C. bombi (53.8%), which are usually found at much lower rates of prevalence45,48. In fact, until now C. bombi was very seldom reported in honeybees21,49. It is also interesting to note the first identification of C. acanthocephali50 in Apis mellifera, with an occurrence of 22.5%, as well as the detection of a putatively new taxon, Trypanosomatidae sp. (GenBank accession number MN038411) which, according to the phylogeny constructed with other trypanosomatid RPB1 sequences, is closely related to C. mellificae, C. acanthocephali and L. passim (Fig. 1). However, the synonymous divergence between this taxon and the later species estimated at the RPB1 locus was 49.6%, 42.9% and 50.4%, respectively, which suggests that it likely corresponds to a different species.

Neighbor-joining phylogeny (RPB1 sequences) showing the evolutionary relationships of trypanosomatid species, including the new taxon detected in this work (Trypanosomatidae sp.; highlighted in bold).
The finding of these and other species rarely detected in honeybees, like C. bombi or C. acanthocephali, suggests that trypanosomatids may have a broader spectrum of hosts than previously reported, although their status as incidental vectors or natural reservoirs, where they may cause a disease, is still to be clarified. Besides, given that whole abdomens were used for DNA extraction, we cannot rule out that the presence of these and other novel species could represent an external contamination of the exoskeleton by flower-visiting rather than a true gut infection.
The massive parallel sequencing approach used here also allowed the detection of two Neogregarinorida species, Apycistis bombi and a second taxon (GenBank accession number MN031271) that was nearly as frequent as the former (Table 1). Its phylogenetic position within the group is displayed in Fig. 2 and the estimates of silent nucleotide divergence between this new taxon and A. bombi and Mattesia spp. were 4.6% and 2.3%, respectively, which are of the same magnitude as those observed between other neogregarine species. However, it must be emphasized that the SSU is not ideal to infer taxonomic relationships among gregarine species, which show very heterogeneous rates of evolution at this locus51.

Neighbor-joining phylogeny (SSU sequences) showing the evolutionary relationships of neogregarine species, including the new taxon detected in this work (Neogregarinorida sp; highlighted in bold).
The analyses of diversity showed that parasites of the same groups were found in the same sample more often than would be expected assuming random expectations (Table 2). In principle such an effect could be attributed to the lack of stochasticity of parasites present in consecutive samples. For instance, if a parasite persists for long periods of time in a colony (low turnover) its presence at any given point in time would make its detection more likely in the following samples. In addition, parasites would tend to be associated with other species found in the preceding samples. But in contrast with these predictions, the observed surplus of co-infections was only detected amongst species of the same group (specifically amongst trypanosomatids and neogregarines) but not between species of different groups. In fact, no significant pairwise associations were observed between N. ceranae, L. passim, and A. bombi, the three species with lower turnover rates of their groups (approximated as the inverse of the mean number of consecutive positive samples; Supplementary Table S2). These observations argue against the hypothesis of a correlation in parasite composition between consecutive samples and suggest a role for other parasite group-specific factors in modelling the patterns of infection over time (Supplementary Table S3).
Seasonal analysis of parasite diversity
Eighty percent of the samples harboured more than one parasite and the average number of parasite species detected per sample was 3.5 ± 0.20 (Supplementary Table S4). No significant differences between interior and exterior bees were observed. These results go in line with previous recent data49 and suggest that mixed infections are the norm rather than the exception in honeybees28.
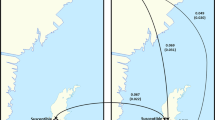
Considering that environmental factors are thought to have a strong influence on the parasite composition of the colonies26, we analysed the seasonal variation of parasite frequency and diversity in the five colonies over time. Samples were grouped by season according to the sampling scheme detailed in the Experimental Procedures section (Fig. 3).

Representation of the collection scheme used for Ion PGM sequencing. Sampling dates are indicated (dotted lines). Interior and exterior honeybees are represented by white and black arrows, respectively.
Nosematids and trypanosomatids displayed a nearly identical pattern of seasonal variation with slight fluctuations that reached the highest frequencies during spring and the lowest in summer (Fig. 4). The strong within-group associations described above meant that most species of these groups shared this seasonal pattern (Supplementary Table S5), with punctual deviations such as C. bombi, which was slightly less prevalent in winter, or C. acanthocephali, which was more common in autumn. Current samples sizes limited our ability to determine the statistical significance of these within-group variation.

Mean fraction of positive samples (%) across seasons. Raw data as in Supplementary Table S5; N. thomsoni and Trypanosomatidae sp. were excluded due to their low frequency. Error bars represent 95% confidence intervals.
These trends were consistent along the two consecutive complete annual cycles (autumn-winter-spring-summer) included in the study (Fig. 3) and suggest that, although nosematid and trypanosomatid species could be found all year round, they find it harder to overcome the hot dry summers in the Iberian Meseta, with average daily high temperatures over 30 °C and monthly rainfall below 20 mm in July and August. These climatic conditions, together with the simultaneous increase in the number of individuals in the colonies52, contribute to explain the reduction in occurrence of these parasites in summer, which was already observed in previous reports based either on prevalence data (e.g. Stevanovic et al., 201353 and Gisder et al., 201754 for N. ceranae; or Glenny et al., 201726 for L. passim) or parasite loads (e.g. Higes et al., 200852 and Chen et al., 201255 for N. ceranae; Glenny et al., 201726 and D’Alvise et al., 201956 for L. passim; or Vejnovic et al., 201830 for L. passim and N. ceranae).
Contrastingly, the occurrence of the two neogregarine species followed a distinct pattern (P < 0.001 in a Chi-square test of goodness of fit) with little variation from winter through summer and a sharp drop in autumn (Fig. 4 and Supplementary Table S5). This seasonal trend could be related with the change of activity (reduction of foraging and breeding) and progressive reduction of the population of the colony during autumn. Indeed, it has been demonstrated that the foraging activity of adult honeybees has an important influence on their exposure to pathogens, as fruit, flowers and other food sources are frequently contaminated by other pollinators and function as fomites, contributing to the spread of parasites across visiting insect species15,21,52,57,58. However, no significant differences were detected in the fraction of positive samples (Supplementary Tables S1 and S6), nor in the levels of species diversity (Supplementary Table S7) or seasonal trends (Supplementary Figure S1) between interior and exterior bees throughout this study, which suggests a very rapid transmission of enteric parasites between the two groups of bees, most likely through mouth-to-mouth transmission of fluids (trophallaxis). Given that our survey involved a limited number of colonies, it would be of great interest to carry out larger studies (including samples from different locations) to verify if their seasonal trends are similar to those described above.
It should be noticed that the variation of species diversity was associated with seasonal changes but it did not display any obvious relationship with the collapse of the colonies. For instance, three colonies collapsed in mid-autumn 2015 and the other two, with no clinical signs by then, died in late summer 2016 (Fig. 3). In addition, high parasite diversity values were already recorded at early stages of the experiment (e.g. between six and seven species were detected in colonies PR-05, PR-10 and PR-16 in the second sampling date; data from Supplementary Table S1). Also, the average number of species per sample across colonies did not reveal any tendency to increase in the five sampling dates prior to their collapse: 5.8, 4.6, 5.6, 4.6 and 2.6, respectively (data from Supplementary Table S1). These results argue against the hypothesis of a build-up of parasite diversity before the colonies’ collapse, which means that the role of these parasites as drivers of the death of the colonies does not probably reside on the actual number of species but on the presence of one or a combination of them. In this regard, N. ceranae was the only species under study present in the samples of all colonies prior to their collapse. The pathogenic effects of this parasite on honeybees, which lead to the shortening of the workers’ lifespan and ultimately to the failure of the colonies52,59, are well known in Southern Europe where this highly prevalent pathogen is considered a serious threat.
This work was focused on the analysis of parasite diversity from a qualitative point of view, so pathogens were identified but not quantified. Given that parasite load might be an important factor to explain the effect of the infections on colony health, future research should take this aspect into account, as well as extend the screening to additional infectious agents, such as viruses (e.g. deformed wing virus, black queen cell virus, among others), bacteria-like (e.g. Melissococcus plutonius, Paenibacillus larvae) and mites (e.g. Varroa, Acarapis woodi)60,61 in order to get a more comprehensive picture of the influence that these factors (pathogen diversity, load and interactions) may have on colony losses.
Methods
Samples and DNA extraction
Five homogeneous nucleus hives of A. mellifera iberiensis (PR-01, PR-05, PR-10, PR-16 and PR-21), whose sister queens were born in 2013, were introduced in May of that same year in one of the experimental apiaries of the CIAPA-IRIAF in Marchamalo (Spain). All of them were managed in the same way (including a treatment with Apitraz, once a year, to comply with the legislation regarding the control of varroosis in Spain).
The sampling scheme started 16 months later (autumn 2014) in order to allow time for the colonies to set and to minimize potential confounding effects caused by the stochastic variation of the parasite populations in the initial stages of the experiment (Fig. 3).
By late November 2015 one of the colonies died (PR-10) and two others displayed no harvesting activity but kept interior bees for a few more weeks (PR-01 and PR-16). The other two colonies were asymptomatic at that time and lasted until the end of the following summer (August 2016).
The samples were selected with the objectives of having a representation of all seasons and, above all, investigating if the latest phases of survival of the colonies displayed different patterns of pathogen diversity than the previous ones. Thus, the selection covered one collection per trimester during, roughly, the first year and two from October 2015 until the collapse of the colonies (Fig. 3).
Twenty five exterior and twenty-five interior honeybees were collected from each colony at the selected dates (Fig. 3). Genomic DNA was separately extracted from the abdomen of each individual using the BS96 DNA Tissue extraction protocol in a BioSprint robot (QIAGEN), as previously described35. DNA aliquots from 15 individuals were then combined to obtain pooled DNA samples representing the exterior and interior bees collected at each sampling date. In total there were 80 pooled samples, which means that there was a ≥95% likelihood of detecting a parasite present at prevalence ≥20% over the course of the experiment. DNA concentrations were measured with a spectrophotometer (Nanodrop 2000; Thermo Fisher Scientific) and set at final concentration of approximately 40 ng/ µl for PCR-amplification.
Primers and PCR amplification
The design of universal primers amplifying the greatest possible number of nosematid, trypanosomatid and neogregarine species was performed with Primer Blast (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) after identifying conserved regions within the alignments of multiple Nosematidae (SSU and Actin), Leishmaniinae (SSU and RPB1) and Neogregarinorida (SSU) sequences available in GenBank. Expected amplicon sizes were shorter than 300 bp, as required for subsequent Ion-PGM sequencing (Table 3).
PCRs were performed using the Phusion High-Fidelity DNA Polymerase (Thermo Fisher Scientific), which works at slightly higher annealing temperatures than other polymerases. PCR reactions were performed in 20 μl volumes containing 10.4 μl of H2O, 4 μl of 5X Phusion HF Buffer, 0.4 μl of dNTP mix 10 mM, 2 μl of each primer 5 μM, 0.2 μl of Phusion DNA Polymerase and 1 μl of DNA. Cycling conditions were set according to manufacturer’s instructions and consisted of an initial denaturalization at 98 °C for 30 s, followed by 45 cycles of 98 °C for 10 s, 60–66 °C for 30 s (Table 3) and 72 °C for 10 s, and a final extension of 8 min at 72 °C. Negative controls were included in all reactions to monitor for any potential contamination.
To avoid amplification biases in multi-template PCRs, two independent 20 μl reactions were run for each sample. PCRs were checked by 2.0% agarose gel electrophoresis. The resulting amplicons from each sample were pooled, purified with Agentcourt AMPure XP (Beckman Coulter) and quantified with Qubit 2.0 (Thermo Fisher Scientific).
Ion PGM sequencing
Library preparation and sequencing
Barcoded libraries were prepared at the Fundación Pública Galega de Medicina Xenómica (Santiago de Compostela, Spain) by combining equimolar concentrations of the amplicons obtained for each sample with the aid of the Ion Plus Fragment Library Kit and Ion Xpress Barcode Adaptors 1-96 Kit (Thermo Fisher Scientific).
Template preparation and enrichment were carried out on the Ion Chef System (Thermo Fisher Scientific). The template-coated Ion Sphere particles were loaded on an Ion 318 Chip v2 (Thermo Fisher Scientific) and sequenced on an Ion PGM (Thermo Fisher Scientific), which can provide up to 5 million reads/ run, with an average length of 200-300 base pairs32.
Data analysis
Raw Ion PGM reads were processed with the Ion Torrent Suite software, which scored the quality of the runs and sorted the data according to the barcodes. The assignment of reads to a specific amplicon was made based on the sequences of the primers (which were removed from further analyses). Once identified, these were divided into individual fasta files providing the number and sequence of the haplotypes detected for each amplicon in a sample, as well as the number of reads on each direction (forward and reverse). The latter was used as a control to differentiate sequencing errors from genuine mutations, as indel formation is a frequent and strand-biased artefact of this sequencing technology62.
Evolutionary analyses
Phylogenetic analyses were performed with MEGA763 using both sequences retrieved from GenBank (whole-genome shotgun contigs for trypanosomatids and nucleotides for neogregarines, respectively) and sequences generated during the current study. The best model for each locus was selected by applying the Akaike information criterion (AIC) and the reliability of the resulting tree topologies was tested by bootstrap support (1000 replicates). The evolutionary distances among trypanosomatid sequences (RPB1) were estimated using the Tamura 3-parameter model following a gamma distribution whereas those among neogregarine species (SSU) were computed using the Kimura 2-parameter method (also following a gamma distribution). Divergence estimates between species were calculated with DnaSP v664.
Data availability
Sequences from previously undescribed parasites were submitted to GenBank (Accession Numbers MN031271.1 and MN038411.1).
References
Potts, S. G. et al. Safeguarding pollinators and their values to human well-being. Nature 540, 220–229, https://doi.org/10.1038/nature20588 (2016).
VanEngelsdorp, D. & Meixner, M. D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 103, https://doi.org/10.1016/j.jip.2009.06.011 (2010).
Neumann, P. & Carreck, N. L. Honey bee colony losses. J. Apic. Res. 49, 1–6, https://doi.org/10.3896/IBRA.1.49.1.01 (2010).
Gray, A. et al. Loss rates of honey bee colonies during winter 2017/18 in 36 countries participating in the COLOSS survey, including effects of forage sources. J. Apic. Res. 58, 479–485, https://doi.org/10.1080/00218839.2019.1615661 (2019).
Brodschneider, R. et al. Multi-country loss rates of honey bee colonies during winter 2016/2017 from the COLOSS survey. J. Apic. Res. 57, 452–457, https://doi.org/10.1080/00218839.2018.1460911 (2018).
Aizen, M. A. & Harder, L. D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 19, 915–918, https://doi.org/10.1016/j.cub.2009.03.071 (2009).
Moritz, R. F. A. & Erler, S. Lost colonies found in a data mine: global honey trade but not pests or pesticides as a major cause of regional honeybee colony declines. Agric. Ecosyst. Env. 216, 44–50, https://doi.org/10.1016/j.agee.2015.09.027 (2016).
Chauzat, M. P. et al. Risk indicators affecting honeybee colony survival in Europe: one year of surveillance. Apidologie 47, 348–378, https://doi.org/10.1007/s13592-016-0440-z (2016).
Smith, K. M. et al. Pathogens, pests, and economics: drivers of honey bee colony declines and losses. EcoHealth 10, 434–445, https://doi.org/10.1007/s10393-013-0870-2 (2013).
Potts, S. G. et al. Global pollinator declines: trends, impacts and drivers. Trend Ecol. Evol. 25, 345–353, https://doi.org/10.1016/j.tree.2010.01.007 (2010).
Goulson, D., Nicholls, E., Botías, C. & Rotheray, E. L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, 1255957, https://doi.org/10.1126/science.1255957 (2015).
Sánchez-Bayo, F. & Wyckhuys, K. A. G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 232, 8–27, https://doi.org/10.1016/j.biocon.2019.01.020 (2019).
Cable, J. et al. Global change, parasite transmission and disease control: lessons from ecology. Philos. Trans. R. Soc. B: Biol. Sci. 372, 20160088, https://doi.org/10.1098/rstb.2016.0088 (2017).
Vanbergen, A. J. & Initiative, t. I. P. Threats to an ecosystem service: pressures on pollinators. Front. Ecol. Environ. 11, 251–259, https://doi.org/10.1890/120126 (2013).
Fürst, M. A., McMahon, D. P., Osborne, J. L., Paxton, R. J. & Brown, M. J. F. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 506, 364–366, https://doi.org/10.1038/nature12977 (2014).
Seibold, S. et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 574, 671–674, https://doi.org/10.1038/s41586-019-1684-3 (2019).
González-Varo, J. P. et al. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 28, 524–530, https://doi.org/10.1016/j.tree.2013.05.008 (2013).
Evison, S. E. et al. Pervasiveness of parasites in pollinators. Plos One 7, e30641, https://doi.org/10.1371/journal.pone.0030641 (2012).
Higes, M. et al. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 1, 110–113, https://doi.org/10.1111/j.1758-2229.2009.00014.x (2009).
Cornman, R. S. et al. Pathogen webs in collapsing honey bee colonies. PLoS One 7, e43562, https://doi.org/10.1371/journal.pone.0043562 (2012).
Graystock, P., Goulson, D. & Hughes, W. O. Parasites in bloom: flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proc. Biol. Sci. 282, 20151371, https://doi.org/10.1098/rspb.2015.1371 (2015).
Fries, I. & Camazine, S. Implications of horizontal and vertical pathogen transmission for honey bee epidemiology. Apidologie 32, 199–214, https://doi.org/10.1051/apido:2001122 (2001).
Loope, K. J., Baty, J. W., Lester, P. J. & Wilson Rankin, E. E. Pathogen shifts in a honeybee predator following the arrival of the Varroa mite. Proc. Biol. Sci. 286, 20182499, https://doi.org/10.1098/rspb.2018.2499 (2019).
Graystock, P. et al. The Trojan hives: pollinator pathogens, imported and distributed in bumblebee colonies. J. Appl. Ecol. 50, 1207–1215, https://doi.org/10.1111/1365-2664.12134 (2013).
Telfer, S. et al. Parasite interactions in natural populations: insights from longitudinal data. Parasitology 135, 767–781, https://doi.org/10.1017/S0031182008000395 (2008).
Glenny, W. et al. Honey bee (Apis mellifera) colony health and pathogen composition in migratory beekeeping operations involved in California almond pollination. PLoS One 12, e0182814, https://doi.org/10.1371/journal.pone.0182814 (2017).
Cavigli, I. et al. Pathogen prevalence and abundance in honey bee colonies involved in almond pollination. Apidologie 47, 251–266, https://doi.org/10.1007/s13592-015-0395-5 (2016).
Runckel, C. et al. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS One 6, e20656, https://doi.org/10.1371/journal.pone.0020656 (2011).
Waters, T. L. The distribution and population dynamics of the honey bee pathogens Crithidia mellificae and Lotmaria passim in New Zealand (Masters’ Thesis), http://hdl.handle.net/10063/7058, (2018).
Vejnovic, B. et al. Quantitative PCR assessment of Lotmaria passim in Apis mellifera colonies co-infected naturally with Nosema ceranae. J. Invertebr. Pathol. 151, 76–81, https://doi.org/10.1016/j.jip.2017.11.003 (2018).
Buendía, M. et al. Epidemiological study of honeybee pathogens in Europe: The results of Castilla-La Mancha (Spain). Span. J. Agric. Res. 16, e0502, https://doi.org/10.5424/sjar/2018162-11474 (2018).
Kulski, J. K. In Next generation sequencing - Advances, applications and challenges; available from: https://www.intechopen.com/books/next-generation-sequencing-advances-applications-and-challenges/next-generation-sequencing-an-overview-of-the-history-tools-and-omic-applications (ed Jerzy Kulsi) (IntechOpen, 2016).
Housley, D. J., Zalewski, Z. A., Beckett, S. E. & Venta, P. J. Design factors that influence PCR amplification success of cross-species primers among 1147 mammalian primer pairs. BMC Genomics 7, 253, https://doi.org/10.1186/1471-2164-7-253 (2006).
Fredriksson, N. J., Hermansson, M. & Wilen, B. M. The choice of PCR primers has great impact on assessments of bacterial community diversity and dynamics in a wastewater treatment plant. Plos One 8, e76431, https://doi.org/10.1371/journal.pone.0076431 (2013).
Cepero, A. et al. Holistic screening of collapsing honey bee colonies in Spain: a case study. BMC Res. Notes 7, 649, https://doi.org/10.1186/1756-0500-7-649 (2014).
Wilson, G. G. & Burke, J. M. Nosema thomsoni n. sp., a microsporidian from Choristoneura conflictana (Lepidoptera: Tortricidae). Can. J. Zool. 49, 786–788, https://doi.org/10.1139/z71-118 (1971).
Malysh, J. M. et al. Morphological and genetic diversity of microsporidia infecting beet webworm Loxostege sticticalis (L.) (Pyraloidea, Crambidae) in Russia. Evraziat. Entomol. Z. 12, 543–548 (2013).
Li, J. et al. Diversity of Nosema associated with bumblebees (Bombus spp.) from China. Int. J. Parasitol. 42, 49–61, https://doi.org/10.1016/j.ijpara.2011.10.005 (2012).
Vilcinskas, A., Stoecker, K., Schmidtberg, H., Röhrich, C. R. & Vogel, H. Invasive harlequin ladybird carries biological weapons against native competitors. Science 340, 862, https://doi.org/10.1126/science.1234032 (2013).
Ravoet, J. et al. Widespread occurrence of honey bee pathogens in solitary bees. J. Invertebr. Pathol. 122, 55–58, https://doi.org/10.1016/j.jip.2014.08.007 (2014).
Schoonvaere, K., Smagghe, G., Francis, F. & de Graaf, D. C. Study of the metatranscriptome of eight social and solitary wild bee species reveals novel viruses and bee parasites. Front. Microbiol. 9, 177, https://doi.org/10.3389/fmicb.2018.00177 (2018).
Pereira, Kd. S., Meeus, I. & Smagghe, G. Honey bee-collected pollen is a potential source of Ascosphaera apis infection in managed bumble bees. Sci. Rep. 9, 4241, https://doi.org/10.1038/s41598-019-40804-2 (2019).
Kudo, R. Notes on Nosema apis Zander. J. Parasitol. 7, 85–90, https://doi.org/10.2307/3270960 (1920).
Chemurot, M., De Smet, L., Brunain, M., De Rycke, R. & de Graaf, D. C. Nosema neumanni n. sp. (Microsporidia, Nosematidae), a new microsporidian parasite of honeybees, Apis mellifera in Uganda. Eur. J. Protistol. 61, 13–19, https://doi.org/10.1016/j.ejop.2017.07.002 (2017).
Ravoet, J. et al. Differential diagnosis of the honey bee trypanosomatids Crithidia mellificae and Lotmaria passim. J. Invertebr. Pathol. 130, 21–27, https://doi.org/10.1016/j.jip.2015.06.007 (2015).
Arismendi, N., Bruna, A., Zapata, N. & Vargas, M. PCR-specific detection of recently described Lotmaria passim (Trypanosomatidae) in Chilean apiaries. J. Invertebr. Pathol. 134, 1–5, https://doi.org/10.1016/j.jip.2015.12.008 (2016).
Stevanovic, J. et al. Species-specific diagnostics of Apis mellifera trypanosomatids: A nine-year survey (2007-2015) for trypanosomatids and microsporidians in Serbian honey bees. J. Invertebr. Pathol. 139, 6–11, https://doi.org/10.1016/j.jip.2016.07.001 (2016).
Xu, G. et al. Triplex real-time PCR for detection of Crithidia mellificae and Lotmaria passim in honey bees. Parasitol. Res. 117, 623–628, https://doi.org/10.1007/s00436-017-5733-2 (2017).
Bartolomé, C. et al. A new multiplex PCR protocol to detect mixed trypanosomatid infections in species of Apis and Bombus. J. Invertebr. Pathol. 154, 37–41, https://doi.org/10.1016/j.jip.2018.03.015 (2018).
Hanson, W. L. & McGhee, R. B. The biology and morphology of Crithidia acanthocephali n. sp., Leptomonas leptoglossi n. sp., and Blastocrithidia euschisti n. sp.*. J. Protozool. 8, 200–204, https://doi.org/10.1111/j.1550-7408.1961.tb01204.x (1961).
Cavalier-Smith, T. Gregarine site-heterogeneous 18S rDNA trees, revision of gregarine higher classification, and the evolutionary diversification of Sporozoa. Eur. J. Protistol. 50, 472–495, https://doi.org/10.1016/j.ejop.2014.07.002 (2014).
Higes, M. et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 10, https://doi.org/10.1111/j.1462-2920.2008.01687.x (2008).
Stevanovic, J. et al. Characteristics of Nosema ceranae infection in Serbian honey bee colonies. Apidologie 44, 522–536, https://doi.org/10.1007/s13592-013-0203-z (2013).
Gisder, S., Schuler, V., Horchler, L. L., Groth, D. & Genersch, E. Long-term temporal trends of Nosema spp. Infection prevalence in Northeast Germany: Continuous spread of Nosema ceranae, an emerging pathogen of honey bees (Apis mellifera), but no general replacement of Nosema apis. Front. Cell Infect. Microbiol. 7, 301, https://doi.org/10.3389/fcimb.2017.00301 (2017).
Chen, Y. W., Chung, W. P., Wang, C. H., Solter, L. F. & Huang, W. F. Nosema ceranae infection intensity highly correlates with temperature. J. Invertebr. Pathol. 111, 264–267, https://doi.org/10.1016/j.jip.2012.08.014 (2012).
D’Alvise, P., Seeburger, V., Gihring, K., Kieboom, M. & Hasselmann, M. Seasonal dynamics and co-occurrence patterns of honey bee pathogens revealed by high-throughput RT-qPCR analysis. Ecol. Evol. 9, 10241–10252, https://doi.org/10.1002/ece3.5544 (2019).
Durrer, S. & Schmid-Hempel, P. Shared use of flowers leads to horizontal pathogen transmission. Proc. R. Soc. Lond. B: Biol. Sci. 258, 299–302, https://doi.org/10.1098/rspb.1994.0176 (1994).
Bodden, J. M., Hazlehurst, J. A. & Wilson Rankin, E. E. Floral traits predict frequency of defecation on flowers by foraging bumble bees. J. Insect Sci. 19, https://doi.org/10.1093/jisesa/iez091 (2019).
Martín-Hernández, R. et al. Nosema ceranae in Apis mellifera: a 12 years postdetection perspective. Environ. Microbiol. 20, 1302–1329, https://doi.org/10.1111/1462-2920.14103 (2018).
Evans, J. D. & Schwarz, R. S. Bees brought to their knees: microbes affecting honey bee health. Trends Microbiol. 19, 614–620, https://doi.org/10.1016/j.tim.2011.09.003 (2011).
Steinhauer, N. et al. Drivers of colony losses. Curr. Opin. Insect Sci. 26, 142–148, https://doi.org/10.1016/j.cois.2018.02.004 (2018).
Bragg, L. M., Stone, G., Butler, M. K., Hugenholtz, P. & Tyson, G. W. Shining a light on dark sequencing: characterising errors in Ion Torrent PGM data. PLoS Comput. Biol. 9, e1003031, https://doi.org/10.1371/journal.pcbi.1003031 (2013).
Kumar, S., Stecher, G. & Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 33, 1870–1874, https://doi.org/10.1093/molbev/msw054 (2016).
Rozas, J. et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 34, 3299–3302, https://doi.org/10.1093/molbev/msx248 (2017).
Acknowledgements
This work was funded by the Instituto Nacional de Investigación y Tecnología Agraria (INIA) [grant numbers RTA2013-00042-C10-6 and E-RTA2014-00003-C03-01and 03]. The authors wish to thank J. Almagro, J. García, V. Albendea, C. Uceta, M. Gajero, T. Corrales (Laboratorio de Patología Apícola, Centro de Investigación Apícola y Agroambiental, IRIAF, Junta de Comunidades de Castilla-La Mancha), and N. Franjo (Fundación Pública Galega de Medicina Xenómica) for their technical support.
Author information
Authors and Affiliations
Contributions
C.B., R.M.-H., M.H. and X.M. conceived the experiments and designed the study. C.B. and X.M. performed the experiments with the help of M.B.-A., M.B., and B.S. The bioinformatics analysis was carried out by J.A. and the results were analysed by C.B. and X.M., who also contributed to write the manuscript with the collaboration of R.M.-H. and M.-H. All authors (C.B., M.B.-A., M.B., B.S., J.A., A.C., R.M.-H., M.H. and X.M.) reviewed and approved the submitted manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bartolomé, C., Buendía-Abad, M., Benito, M. et al. Longitudinal analysis on parasite diversity in honeybee colonies: new taxa, high frequency of mixed infections and seasonal patterns of variation. Sci Rep 10, 10454 (2020). https://doi.org/10.1038/s41598-020-67183-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-67183-3
This article is cited by
-
Current status of Nosema spp. infection cases in apis mellifera in eurasian countries and Ptp3 gene haplotypes in the Republic of Tatarstan, Russia
Veterinary Research Communications (2024)
-
A tale of two parasites: Responses of honey bees infected with Nosema ceranae and Lotmaria passim
Scientific Reports (2023)
-
Frequent Parasitism of Apis mellifera by Trypanosomatids in Geographically Isolated Areas with Restricted Beekeeping Movements
Microbial Ecology (2023)
-
Bee Trypanosomatids: First Steps in the Analysis of the Genetic Variation and Population Structure of Lotmaria passim, Crithidia bombi and Crithidia mellificae
Microbial Ecology (2022)
-
Identification of pathogens in the invasive hornet Vespa velutina and in native Hymenoptera (Apidae, Vespidae) from SW-Europe
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
