
Abstract
Hormones and placental factors are thought to underlie the maternal immunological changes during pregnancy. However, as several intestinal microbiota are linked to immune modulations, we hypothesized that the intestinal microbiota are altered during pregnancy in favor of species associated with pregnancy associated immune modulations. We studied the fecal microbiota composition (MITchip) and intestinal and peripheral immune cells (microarray and flow cytometry) in pregnant and non-pregnant C57BL/6 and BALB/c mice. Pregnancy influenced intestinal microbiota diversity and composition, however in a mouse strain dependent way. Pregnant BALB/c mice had, among others, a relative higher abundance of Lactobacillus paracasei et rel., Roseburia intestinalis et rel. and Eubacterium hallii et rel., as compared to non-pregnant BALB/c mice, while the microbiota composition in B6 mice hardly changed during pregnancy. Additionally, intestinal immunological pathways were changed during pregnancy, however again in a mouse strain dependent way. Correlations between various bacteria and immunological genes were observed. Our data do support a role for the microbiome in changing immune responses in pregnancy. However, other factors are also involved, such as for instance changes in SCFA or changes in sensitivity to bacteria, since although immunological changes are observed in B6 mice, hardly any changes in microbiota were found in this strain. Follow up studies are needed to study the exact relationship between these parameters.
Similar content being viewed by others
Introduction
Pregnancy is associated with immunological adaptations of the mother in order to tolerate and support the development of the semi-allogeneic fetus. Changes in both in the innate and the specific immune response are reported during pregnancy. A decrease in the T helper 1 (Th1)/T helper 2 (Th2) ratio1,2 and in natural killer (NK) cells, and an increase in regulatory T cell (Treg) numbers3,4 has been found during pregnancy. It is known that peripheral immune responses are influenced by the placenta by direct contact with fetal tissue in the placenta, due to circulation of the immune cells through the placenta, or due to contact with soluble products produced by the placenta, such as cytokines, micro-particles and exosomes5,6,7. However, recent studies showed that the intestinal microbiome changes during pregnancy, which may also underlie the immunological changes during pregnancy8.
Modulation of immune cells can be induced by microbiota species present in the intestinal lumen9,10,11,12. Segmented filamentous bacteria for example showed to be able to induce the development of intestinal T helper 17 (Th17) cells13, while several bacteria species from the genus Clostridium have been shown to promote the accumulation of intestinal Tregs9,10. Additionally, Lactobacillus plantarum has been shown to enhance the number of peripheral Tregs12. One of the mechanisms by which bacteria can modulate immune cells is by producing short chain fatty acids (SCFA), such as butyrate, which has been shown to induce the differentiation of Tregs in the colon14. This is particularly true for Clostridium species from the clusters IV and XIVa9. However, whether intestinal bacteria are also involved in changes in peripheral immune cells during pregnancy, and whether intestinal immune cells are involved in this, is unknown.
In this study we investigated a possible relationship between microbiota and the immunological adaptations at the end of pregnancy in mice. To this end, we studied the effect of pregnancy on the microbiota composition and the effect of pregnancy on both intestinal immune cells and peripheral immune cells with a microarray (colon tissue) and flow cytometry (spleen and mesenteric lymph nodes (MLN)). We combined the transcriptomics data and the microbiota data and performed a bio-mathematical analysis, in order to find bacterial species, which may potentially be related to pregnancy induced differences in intestinal immunity. Non-pregnant mice served as control. We used two different mouse strains (C57BL/6 and BALB/c) that were shown to have different microbiota profiles and intestinal immunological responses15,16, to study the consequences of a different pre-pregnancy starting point in the intestinal microbiota-host interactions.
Results
Intestinal microbiota composition changed during pregnancy, but in a mouse strain dependent way
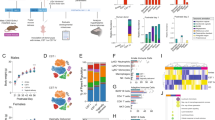
We determined the microbiota composition of non-pregnant and pregnant mice in both strains using the phylogenetic microarray, the mouse intestinal tract Chip (MITChip). We determined the richness (number of unique species) and Shannon diversity (calculation between richness and evenness) of the microbiota composition. The data was analyzed with a Two-way ANOVA (TWA), to assess if there was an interaction between pregnancy and strain on the microbiota composition in the colon. A Bonferroni post-test was performed when interaction was found, to determine whether the effect of pregnancy was the same in both mouse strains. Overall, pregnancy decreased both microbiota diversity and richness (TWA, p < 0.05) (Fig. 1A,B), but the ratio of Firmicutes/Bacteroidetes was not influenced by pregnancy (Fig. 1C).

Effect of pregnancy on fecal microbiota characteristics. Shannon diversity (A), richness (B) and the firmicutes/bacteroidetes ratio (C) in the fecal microbiota of pregnant and non-pregnant BALB/c and B6 (n = 5). Results are shown as dot plots + means and were tested for overall strain and pregnancy effects using a Two-way ANOVA, followed by a Bonferroni post-hoc test to test for strain specific pregnancy effects when interaction was found. Significant pregnancy effects are indicated with dashed lines (p < 0.05). RDA plot (D) showing the variation explained by the components genotype and pregnancy (n = 5). The total variation that can be explained by the variables genotype (14.3%) and pregnancy (12.5%) is 26.8%. Both variables are significant in explaining the variation (Monte Carlo permutation, p < 0.05).
Redundancy analysis showed that the total variation in microbiota composition explained by the variables genotype and pregnancy is 26.8%. Pregnancy explained 12.5% of the variance in microbiota composition (Fig. 1D). However, pregnancy-induced differences in microbiota at species like level were mainly observed in BALB/c mice. In B6 mice, pregnancy did not affect the microbiota at species like level. Pregnant BALB/c mice had a significantly relative higher abundance of, among others, Allobaculum et rel., Lactobacillus salivarius et rel., Lactobacillus plantarum et rel., Unclassified Clostridiales XVI, Clostridium perfringens et rel., Lactobacillus paracasei et rel., and Roseburia intestinalis et rel., as compared to non-pregnant BALB/c mice (Table 1).
Pregnancy influenced the expression of genes related to intestinal immune pathways in a mouse strain dependent way
Next, we determined the effect of pregnancy on intestinal immunological pathways with a microarray on colonic tissue. In the colon, 1032 genes were differentially expressed between pregnant and non-pregnant BALB/c mice, and 789 genes were differentially expressed in B6 mice. The two strains shared 161 differentially expressed genes between pregnant and non-pregnant females in the colon (both up- and down-regulated).
To gain insight into the biological role of the genes which were influenced by pregnancy, we looked into the pathways in which these genes are involved using Ingenuity Pathway Analysis (IPA), with special focus on pathways related to immunology (Table 2). In both mouse strains, IPA showed that pregnancy significantly influenced the expression of genes related to both the innate and adaptive immunity. However, in the B6 strain, the effect of pregnancy was more pronounced concerning pathways related to immunology. In the colon of B6 mice, pregnancy influenced, among others, the antigen presentation pathway, T cell co-stimulation (CD28/iCOS), dendritic cell (DC) maturation and cytokine signaling (IL-4/IL-8/IL-9). In the colon of BALB/c mice, among others, the antigen presentation pathway, DC maturation and the IL-10 signaling pathway were influenced by pregnancy (Table 2).
Correlation between microbiota species and gene expression profile in the colon
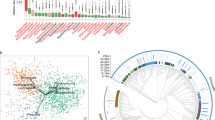
To investigate the relation between microbiota species and immunological gene expression, we combined microbiota and colonic gene expression data from individual pregnant and non-pregnant BALB/c mice, to evaluate direct correlations between gene expression and microbiota composition in these samples (Fig. 2). We only did this for the BALB/c strain as no microbiota species were significantly changed during pregnancy in the B6 strain. We integrated the datasets using a PLS-based canonical correlation approach. In total 600 genes and 30 bacterial groups were retained for the first three components, and clustering of the correlation coefficients showed six main clusters of host genes that correlated negatively (blue) or positively (red) with three main clusters of bacteria.

Correlation between microbiota species and gene expression in the colon. This figure shows a heat-map which correlates the MITChip data (vertical) and the microarray data (horizontal) of pregnant and non-pregnant BALB/c mice. For each mouse the datasets were integrated individually mouse (5 mice per group) and it gives the direct correlations between gene expression and microbiota composition from these samples. Cluster of genes that correlated most positively with a respective group of bacteria are indicated in deep red, while genes that correlated most negatively with a respective group of bacteria are indicated in deep blue. Clusters which are framed in black are discussed in more detail in the text. Six main gene clusters (1–6) were identified and three bacterial clusters (A–C).
Strongest correlations were found between bacterial cluster A with gene expression cluster two and six, bacterial cluster B with gene expression cluster one and four and bacterial cluster C with gene expression cluster three and five. All bacteria in cluster A (such as Allobaculum et rel. and Unclassified Mollicutes) had a higher relative abundances in pregnant BALB/c mice as compared to non-pregnant BALB/c mice. Gene cluster two showed a positive correlation, while gene cluster six showed a negative correlation with bacteria in cluster A. Some genes in cluster two and six were involved in immunological functions, particularly in T cell development (Supplementary Table S1). Also all bacteria in cluster B (such as Eubacterium cylindroides et rel. and Faecalibacterium prausnitzii et rel.) had a higher relative abundances in pregnant BALB/c mice as compared to non-pregnant BALB/c mice. Gene cluster one showed a positive correlation, while gene cluster four showed a negative correlation with bacteria in cluster B. However, almost no genes from cluster one and four were involved in immunological functions (Supplementary Table S2). Interestingly, cluster C contained most of the bacteria which had a significantly lower abundance (Clostridium lactifermentans et rel., Clostridium leptum et rel. and Sporobacter termitidis et rel.) in pregnant BALB/c mice as compared to the non-pregnant BALB/c mice. Gene cluster three showed a negative correlation, while gene cluster five showed a positive correlation with bacteria in cluster C. Many genes from cluster three and five were involved in immunological functions, especially in lymphocyte and T cell development (Supplementary Table S3).
Pregnancy influenced T cells in the MLN and spleen in a mouse strain dependent way
As we found effects of pregnancy on the microbiota composition and intestinal immunological gene expression using a microarray, we subsequently studied immune cell subsets in the MLN and the spleens with flow cytometry. We used the MLN as they are the draining lymph nodes for the intestines. The spleen was used as a reference for systemic immunity. We focused on several T cell subsets, as many genes which were related to pregnancy induced microbiota changes were related to T cells. Again we analyzed the data with a TWA, to understand if there was an interaction between pregnancy and mouse strain on the immune populations. When an interaction was present we tested with a Bonferroni post-test whether the effect of pregnancy was the same in both mouse strains.
In the MLN, overall the percentage of CD4+ cells was higher in pregnant mice (TWA, p < 0.05) (Fig. 3A). The percentage of CD8+ cells in the MLN was not affected by pregnancy (Fig. 3B). Pregnant mice also tended to have a higher percentage of FoxP3+CD4+ cells in the MLN than non-pregnant mice (TWA, p = 0.0776) (Fig. 3C). Pregnancy changed the percentage of CD25+FoxP3− CD4+ cells in the MLN, although in a mouse strain dependent way (interaction between pregnancy and strain TWA, p < 0.05). In pregnant B6 mice, but not in pregnant BALB/c mice, the percentage of CD25+FoxP3− CD4+ cells was higher than in non-pregnant mice (Bonferroni, p < 0.05) (Fig. 3D). Additionally, the percentage of both CD4+ and CD8+ cells expressing CD69 in the MLN was higher in pregnant than non-pregnant mice (TWA, p < 0.05) (Fig. 3E,F).

Effect of pregnancy on T cell populations in the mesenteric lymph nodes. Frequency of T helper cells (CD4+) (A), cytotoxic T cells (CD8+) (B), FoxP3+ Tregs (C), CD25+ FoxP3− effector CD4+ cells (D), CD69+ CD4+ cells (E), and CD69+ CD8+ cells (F) in the mesenteric lymph nodes (MLN) of pregnant and non-pregnant BALB/c and B6 mice. T cytotoxic and T helper cells are expressed as the frequency of CD8+ and CD8− cells within the CD3+ population respectively. Due to technical problems, several MLNs could not be used for immune cell analysis, which reduced the number of mice in these groups. Results are shown as dot plots + means and were tested for overall strain and pregnancy effects using a Two-way ANOVA, followed by a Bonferroni post-hoc test to test for strain specific pregnancy effects when interaction was found. Significant strain effects are indicated with solid lines and significant pregnancy effects are indicated with dashed lines (p < 0.05).
In the spleen, pregnant mice had an overall higher percentage of CD4+ cells than non-pregnant mice (TWA, p < 0.05), while pregnancy had no effect of on the percentage of CD8+ cells (Fig. 4A,B). Pregnancy also influenced the percentage of FoxP3+CD4+ cells in the spleen (Fig. 4C), however, in a mouse strain dependent way (interaction between pregnancy and strain in TWA, p < 0.05). The percentage of FoxP3+CD4+ cells was not changed in pregnant BALB/c mice as compared to non-pregnant BALB/c mice, whereas pregnant B6 mice had a significantly higher percentage of FoxP3+CD4+ cells than non-pregnant B6 mice (Bonferroni, p < 0.05). Pregnancy also affected the percentage of CD25+FoxP3− CD4+ cells in the spleen (Fig. 4D), again in a mouse strain depended way (interaction between pregnancy and strain in TWA, p < 0.05). Only pregnant B6 mice, but not pregnant BALB/c mice, had a significantly higher percentage of CD25+FoxP3− CD4+ cells than non-pregnant B6 mice (Bonferroni, p < 0.05). The percentage of CD4+ cells expressing the early activation marker CD69 was affected by pregnancy in the spleen, although also in a mouse strain dependent way (interaction between pregnancy and strain, TWA, p < 0.05) (Fig. 4E): the percentage of CD4+ cells expressing CD69 was not affected in pregnant BALB/c mice, while it was significantly higher in pregnant B6 mice than non-pregnant mice (Bonferroni, p < 0.05). Pregnancy did not alter the percentage of CD8+ expressing CD69 (Fig. 4F) in the spleen.

Effect of pregnancy on T cell populations in the spleen. Frequency of T helper cells (CD4+) (A), cytotoxic T cells (CD8+) (B), FoxP3+ Tregs (C), CD25+FoxP3− effector CD4+ cells (D), CD69+ CD4+ cells (E), and CD69+ CD8+ cells (F) in the spleens of pregnant and non-pregnant BALB/c and B6 mice. T cytotoxic and T helper cells are expressed as the frequency of CD8+ and CD4+ cells within the CD3+ population respectively. Results are shown as dot plots + means and were tested for overall strain and pregnancy effects using a Two-way ANOVA, followed by a Bonferroni post-hoc test to test for strain specific pregnancy effects when interaction was found. Significant strain effects are indicated with solid lines and significant pregnancy effects are indicated with dashed lines (p < 0.05).
Pregnancy increased CD103 DCs in the mesenteric lymph nodes
There was an interaction between pregnancy and strain for the percentage of DCs in MLN (TWA, p < 0.05) (Fig. 5A). The percentage of DCs expressing CD103 was higher in pregnant mice than non-pregnant mice (TWA, p < 0.05) (Fig. 5B).

Effect of pregnancy on CD103+ dendritic cells in the mesenteric lymph nodes. Percentage of dendritic cells (DCs) (A) and their expression of CD103 (B) in the mesenteric lymph nodes (MLN) of pregnant and non-pregnant BALB/c and B6 mice. DCs are expressed as the percentage of CD11c+ cells within the F4/80−MHC2+ population. Due to technical problems, several MLNs could not be used for immune cell analysis, which reduced the number of mice in these groups. Results are shown as dot plots + means and were tested for overall strain and pregnancy effects using a Two-way ANOVA, followed by a Bonferroni post-hoc test to test for strain specific pregnancy effects when interaction was found. Significant strain effects are indicated with solid lines and significant pregnancy effects are indicated with dashed lines (p < 0.05).
Discussion
Pregnancy is associated with peripheral immune changes3,4. We hypothesized that this may be related to changes in the gut microbiome. We found that pregnancy influenced the intestinal microbiota composition as well as the expression of genes related to immunological pathways in the colon, but in a mouse strain dependent way.
We demonstrate that pregnancy impacts the intestinal microbiota composition in mice, however, in a mouse strain depended way. We showed that at the end of pregnancy, pregnant mice had a lower microbiota diversity and richness than non-pregnant mice, but this decrease was more pronounced in BALB/c mice. Our data appear to be in line with a human study, which demonstrated a decreased diversity in the third trimester as compared to the first trimester of pregnancy in humans8. In general, it is assumed that a microbiome with a low diversity and richness is not beneficial for the host health, as disorders like obesity are general associated with a low microbial diversity and richness17,18. It remains therefore to be investigated why diversity and richness are decreased in pregnancy. However, it might be a functional change as Koren et al. showed that transferring microbiota from third trimester pregnant women to germ-free mice, induced adiposity and insulin insensitivity in these mice8. Therefore, it may be suggested that during pregnancy, one of the functions of the changes in microbiome is enhancing energy absorption in the mother.
When looking at bacterial species like level, we also found changes during pregnancy, however only in the BALB/c strain. During pregnancy, BALB/c mice had a higher abundance of, among others, Eubacterium hallii et rel., Lactobacillus paracasei et rel., Lactobacillus plantarum et rel., and Roseburia intestinalis et rel. These species have been shown to modulate immune responses, and are especially associated with immune tolerance and anti-inflammatory properties12,19,20,21. Lactobacillus paracasei et rel. has been shown to increase the amount of both CD103 expressing DCs, i.e. DCs inducing immune tolerance, and Tregs in the MLN of mice21. This is in accordance with the higher percentage of CD103+ DCs we found in the MLNs of pregnant BALB/c mice. Eubacterium hallii, as well as Roseburia intestinalis are well known butyrate producers19,20, which can induce the differentiation of Tregs in the lamina propria of the colon14. Species of the Lactobacillus genus are well known for their beneficial effects on health, including the fact that they can upregulate Tregs and have anti-inflammatory properties12.
Interestingly, in contrast to the BALB/c strain, pregnancy induced changes in microbiota composition were less apparent in the B6 strain. This may be in contradiction with a study of Gohir et al. (2015), who found a shift in microbiota composition in B6 mice during pregnancy, however they took fecal samples at earlier time-points (day 5, day 10 and day 15 of gestation), which may explain the different findings22. A potential explanation for the lack of changes in the B6 microbiome during pregnancy maybe the fact that non-pregnant B6 females, in contrast to non-pregnant BALB/c, already had a high abundance of bacteria from the Lactobacillus genus. Bacteria from this genus increase in pregnant BALB/c mice. Therefore, there may be no need for changes in bacterial composition during pregnancy in the B6 mice. Despite the lack of changes in the B6 microbiome, peripheral Tregs were increased in these mice during pregnancy. This may indicate that changes in Tregs in B6 mice are induced by other factors than microbiota or that pregnant B6 mice are more sensitive to the effects of microbiota, such as bacteria from the Lactobacillus genus. Additionally, a change in bacterial metabolic products during pregnancy, such as SCFA, may also underlie the change in Tregs in B6. These hypotheses need to be tested in future studies, and currently we investigate the immune modulatory effects of the specific pregnancy enriched bacteria in germ free mice.
We also evaluated the effect of pregnancy on the expression of genes involved in immune pathways in the colon. By using a microarray and Ingenuity Pathway Analysis, we demonstrated that pregnancy altered the expression of many genes involved in immune pathways in the colon. In both mouse strains we found immunological pathways which were differentially regulated by pregnancy. These include, among others, the antigen presenting pathway, DC maturation, T helper cell differentiation and IL-10 signaling. The effect of pregnancy on these pathways, however, was dependent on the mouse strain. Some pathways were modulated by pregnancy in both strains, such as DC maturation and T helper cell differentiation. However, other pathways were only modulated by pregnancy in B6 mice, such as the pathways related to CD28/ICOS, or were only modulated by pregnancy in BALB/c mice, such as interferon signaling. Despite the lack of changes in the B6 microbiome, the expression of immunological genes in the colon was also changed during pregnancy. Again, this may indicate that changes in the immune system in B6 mice are induced by other factors than microbiota or that immunological genes in pregnant B6 mice are more sensitive to the modulating effects of microbiota.
As the present study was an observational study, more experimental data are needed in order to state that the microbiome is driving immune changes during pregnancy. However, our data do support a role for the microbiome in immunological changes during pregnancy, as our data show that the presence of several bacterial species which were up- or down-regulated during pregnancy in BALB/c mice were correlated to clusters of genes in the colon (Fig. 2), of which many are related to immunological functions, such as T cell development. With flow cytometry we aimed to confirm the immunological changes found in the intestine, and indeed we found changes in T cell populations in both the MLN and spleen during pregnancy, however again in a mouse strain dependent way. We demonstrated that pregnancy tended (p = 0.0776) to increase Treg cell numbers in the MLNs, while the percentage of CD103+ DCs in the MLN was only increased in pregnant BALB/c mice. DCs expressing CD103 in the MLN are able to induce the generation of FoxP3+ Treg cells23. Interestingly, in our study there does not seem to be a relation between CD103+ DCs and FoxP3+ Tregs, as in the BALB/c mice we found a higher percentage of intestinal CD103+ DC in pregnant mice, while the percentage of peripheral Tregs was not increased; in B6 mice we found an increased percentage of peripheral Tregs in pregnant mice, while the percentage of intestinal CD103+ DCs was not increased. The reason for this discrepancy is unknown, however, it may be due to differences in expression of co-factors such as TGF-ß or retinoic acid. These cofactors are necessary for CD103+ DCs to drive Treg generation24. Another explanation may be that the induced Tregs are not targeted to the spleen but recruited to the uterus draining lymph nodes25.
In summary, our data showed changes in microbiota composition and expression of genes related to immunological pathways in the colon, during late pregnancy. Although our data do support a potential role for the microbiome in changing immune responses in pregnancy, other factors are also involved, such as for instance changes in SCFA or changes in sensitivity to bacteria, since although immunological changes are observed in B6 mice, hardly any changes in microbiota were found in this strain. Further studies are needed to evaluate the exact relationship between these parameters. Measurements, such the sensitivity of the immune system for bacteria, immune cells in the decidua and earlier time-points in pregnancy should be included in follow up studies to further investigate the relation between the microbiome and pregnancy induced immune adaptations.
Materials and Methods
Mice
All animal experiments were performed after receiving approval of the Animal Care Committee of the Groningen University (DEC#6349 A) and we confirm that all methods were performed in accordance with their guidelines and regulations. Female wild-type B6 and BALB/c mice (n = 20 per strain) were purchased from Harlan (Harlan, Horst, the Netherlands) at an age of eight weeks. All mice were co-housed (four cages, five mice per cage, pregnant and non-pregnant mice were separately housed) in isolated ventilated cages to limit environmental influences. The animals had ad libitum access to a D12450B diet (Research Diets Services, Wijk bij Duurstede, the Netherlands) and water. This diet was given immediately after the mice arrived in the animal facility, this was at least three weeks before sacrifice. Between an age of 12 weeks and 32 weeks (mean age of the mouse per groups is given in Tables 3 and 4) all mice were sacrificed by cervical dislocation under anesthesia. All non-pregnant female mice (n = 10 from each strain) were sacrificed during the diestrus phase of their ovarian cycle to ensure low stable levels progesterone and estrogens. For the pregnant mice, smears were taken daily and, when in the proestrus phase of their ovarian cycle, they were placed (overnight) in a cage with a male wild-type mice from the same strain and placed back in their original cage the next morning. The day of the vaginal plug was day 0 of pregnancy. Pregnant mice were sacrificed at day 18 of gestation. At sacrifice, spleens and MLN’s were removed for immune cell analysis. Approximately 1 cm of proximal colon was frozen in liquid nitrogen and stored at −80 °C prior to RNA isolation and transcriptomics. Feces from the last part of the colon were collected for microbiota profiling. Table 3 provides an overview of the characteristics of all mice and Table 4 provides an overview of the characteristics of the mice which were, per strain, randomly selected for microbiota and gene array analysis (n = 5).
Bacterial DNA extraction and microbiota profiling
Luminal contents from the last part of the distal colon (n = 5 per group) were analyzed by a Mouse Intestinal Tract Chip (MITChip), as previously described26. The Redundancy analysis (RDA) was performed in Canoco 5.0, and visualized in triplots or a PRC plot27,28,29,.
Intestinal microarray analysis
RNA was purified from the proximal colon of mice (n = 5 per group) and RNA expression profiling was performed with an Affymetrix Mouse Gene 1.1 ST array plate (Affymetrix, Santa Clara, CA, USA) as previously described26. Only probe sets with a fold-change of at least 1.2 (up/down) and a p-value < 0.05 were considered to be significantly different.
To gain insight into the biological role of the differently expressed genes during pregnancy, we investigated the pathways in which these genes are involved using Ingenuity Pathway Analysis (IPA) (Ingenuity System). Our IPA analyses included comparison of differentially regulated genes in the colon of pregnant and non-pregnant B6 and BALB/c mice.
Multivariate integration and correlation analysis
To gain insight in the relationship between the genes differently expressed during pregnancy in the colon and the microbiota composition in BALB/c mice, the microarray and MITChip datasets were combined, using the linear multivariate method partial least squares (PLS)28, as described previously29. This integration of datasets per individual mouse gives a direct correlation between gene expression and microbiota composition in these samples (n = 5 per group). Before analysis, the sets of data were log2 transformed, and the canonical correlation framework of PLS was used30. The correlation matrices were visualized in clustered image maps31. Analyses were performed in R using the library mixOmics32.
Isolation of splenic and mesenteric lymph node cells
Single cell suspensions of spleens and MLNs were made as previously described26.
Cell staining and flow cytometry
Spleen and MLN cells were stained for T cell populations. T cells were determined using CD3, T cytotoxic cells with CD8 and T helper cells with CD4. CD69 expression was determined within both the CD8+ and CD4+ cell subsets. In another panel we determined the expression of FoxP3 and CD25, using CD3 and CD8. Within the MLN, also the percentage of DCs, and DCs expressing CD103 were determined using F4/80, MHC2, CD11c and CD103. Antibody specifications are described in Supplementary Table S4.
The antibodies were diluted in a total volume of 25 µl with FACS buffer (PBS + 10% FCS (v/v)). Approximately 1 × 106 spleen or MLN cells were incubated for 20 minutes in FACS buffer (10% FCS (v/v)) with 20% (v/v) normal rat serum (Jackson, Newmarket, UK) and 2% (v/v) Fc block (CD16/32) (Biolegend, Uithoorn, the Netherlands). Next, they were incubated with an extracellular antibody mix for 30 minutes. Then, samples were washed twice with a permeabilization buffer (eBioscience, Vienna, Austria) after which they were incubated with an intracellular blocking medium (20% (v/v) rat serum in permeabilization buffer (eBioscience, Vienna, Austria) for 20 minutes. Subsequently, these cells were incubated with an intracellular antibody mix for 30 minutes and fixed in FACS lysing BD Biosciences, Breda, The Netherlands) solution for 30 minutes. In between all incubation steps washing was performed all incubation steps and the whole procedure was performed on ice and in the dark. Isotype control antibodies were used at the same dilution and purchased from the same company as the extracellular and intracellular antibodies.
Cell samples were analyzed using the LSR-II Flow Cytometer system (BD Biosciences, Breda, the Netherlands), using FACS Diva software. Analysis was performed using FlowJo version 10 software (FlowJo, LLC, Oregon, USA). The gating strategy for T lymphocytes is shown in Supplementary Figure S1. To gate T cells, first lymphocytes were gated based on size in the forward side scatter plot. Next, T cells were determined by selecting CD3+ cells. CD4+ and CD8+ cells were selected within the CD3+ cells. Next, within both the CD4+ and CD8+ population, the percentage of cells expressing CD69 was measured and within the CD4+ cells also the percentage of cells expressing FoxP3 and CD25 was assessed (Supplementary Figure S2). To gate DCs, first living cells were selected based on size in the forward side scatter plot. Within the MHC2+ and F4/80−, DCs were gated as CD11c+. In this population the expression of CD103 was determined (Supplementary Figure S3). All the isotype controls were set at 1% and these gates were copied to the samples with the antibody mix.
Statistics
For Shannon diversity, microbiota richness, the firmicutes/bacteroidetes ratio and flow cytometry, data are expressed as dot plots + means. The Kolmogorov-Smirnov test was used to determine normal distribution of the data. When the data were not normally distributed a log transformation was performed before analysis. The overall effect of pregnancy and strain was determined with a Two-way ANOVA (TWA), followed by a Bonferroni post-hoc test when an interaction between pregnancy and strain was found. For analyzing significant effects of pregnancy on the abundance of bacterial groups within each strain, a Mann–Whitney U test was used. P-values of < 0.05 were considered statistically significant and p-values between 0.05 and 0.1 were defined as a trend. P-values were adjusted for multiple comparisons with the Benjamini-Hochberg procedure.
Availability of data and materials
The gene array datasets generated and analyzed during the current study will be available in the gene expression omnibus from NCBI under ID number xxxx.
References
Veenstra van Nieuwenhoven, A. L. et al. Cytokine production in natural killer cells and lymphocytes in pregnant women compared with women in the follicular phase of the ovarian cycle. Fertil. Steril. 77, 1032–1037 (2002).
Saito, S. et al. Quantitative analysis of peripheral blood Th0, Th1, Th2 and the Th1:Th2 cell ratio during normal human pregnancy and preeclampsia. Clin. Exp. Immunol. 117, 550–555 (1999).
Leber, A., Teles, A. & Zenclussen, A. C. Regulatory T cells and their role in pregnancy. Am. J. Reprod. Immunol. 63, 445–459 (2010).
Svensson-Arvelund, J. et al. The placenta in toxicology. Part II: Systemic and local immune adaptations in pregnancy. Toxicol. Pathol. 42, 327–338 (2014).
Faas, M. M. et al. Plasma from preeclamptic women activates endothelial cells via monocyte activation in vitro. J. Reprod. Immunol. 87, 28–38 (2010).
Sacks, G. P., Clover, L. M., Bainbridge, D. R. J., Redman, C. W. G. & Sargent, I. L. Flow Cytometric Measurement of Intracellular Th1 and Th2 Cytokine Production by Human Villous and Extravillous Cytotrophoblast. Placenta 22, 550–559 (2001).
Mellembakken, J. R. et al. Activation of leukocytes during the uteroplacental passage in preeclampsia. Hypertension 39, 155–160 (2002).
Koren, O. et al. Host Remodeling of the Gut Microbiome and Metabolic Changes during Pregnancy. Cell 150, 470–480 (2012).
Atarashi, K. et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 500, 232–236 (2013).
Atarashi, K. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341 (2011).
Ivanov, I. I. et al. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell. Host Microbe 4, 337–349 (2008).
Smelt, M. J. et al. L. plantarum, L. salivarius, and L. lactis attenuate Th2 responses and increase Treg frequencies in healthy mice in a strain dependent manner. PLoS One 7, e47244 (2012).
Ivanov, I. I. et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 139, 485–498 (2009).
Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450 (2013).
Elderman, M. et al. Sex impacts Th1, Treg, and DCs in both the intestinal and systemic immunity in a mouse strain and location dependent manner. Biology of Sex Differences 7 (2016).
Fransen, F. et al. BALB/c and C57BL/6 Mice Differ in Polyreactive IgA Abundance, which Impacts the Generation of Antigen-Specific IgA and Microbiota Diversity. Immunity 43, 527–540 (2015).
Le Chatelier, E. et al. Richness of human gut microbiome correlates with metabolic markers. Nature 500, 541–546 (2015).
Turnbaugh, P. J. et al. A core gut microbiome in obese and lean twins. Nature 457, 480–484 (2009).
Belenguer, A. et al. Impact of pH on lactate formation and utilization by human fecal microbial communities. Appl. Environ. Microbiol. 73, 6526–6533 (2007).
Duncan, S. H., Hold, G. L., Barcenilla, A., Stewart, C. S. & Flint, H. J. Roseburia intestinalis sp. nov., a novel saccharolytic, butyrate-producing bacterium from human faeces. Int. J. Syst. Evol. Microbiol. 52, 1615–1620 (2002).
Yang, J. et al. Induction of Regulatory Dendritic Cells by Lactobacillus paracasei L9 Prevents Allergic Sensitization to Bovine beta-Lactoglobulin in Mice. J. Microbiol. Biotechnol. 25, 1687–1696 (2015).
Gohir, W. et al. Pregnancy-related changes in the maternal gut microbiota are dependent upon the mother’s periconceptional diet. Gut Microbes 6, 310–320 (2015).
Coombes, J. L. et al. A functionally specialized population of mucosal CD103(+) DCs induces Foxp3(+) regulatory T cells via a TGF-beta- and retinoic acid-dependent mechanism. J Exp Med 204, 1757–1764 (2007).
del Rio, M. L., Bernhardt, G., Rodriguez-Barbosa, J. I. & Forster, R. Development and functional specialization of CD103 + dendritic cells. Immunol. Rev. 234, 268–281 (2010).
Chen, T. et al. Self-specific memory regulatory T cells protect embryos at implantation in mice. J. Immunol. 191, 2273–2281 (2013).
Elderman, M. et al. The effect of age on the intestinal mucus thickness, microbiota composition and immunity in relation to sex in mice. PLoS One 12, e0184274 (2017).
Braak, C. J. F. & Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (Version 5.0). Ithaca, NY: NH (2012).
Boulesteix, A. L. & Strimmer, K. Partial least squares: a versatile tool for the analysis of high-dimensional genomic data. Brief Bioinform 8, 32–44 (2007).
Lange, K. et al. Comparison of the effects of five dietary fibers on mucosal transcriptional profiles, and luminal microbiota composition and SCFA concentrations in murine colon. Mol. Nutr. Food Res. 59, 1590–1602 (2015).
Le Cao, K. A., Martin, P. G., Robert-Granie, C. & Besse, P. Sparse canonical methods for biological data integration: application to a cross-platform study. BMC Bioinformatics 10, 34-2105-10-34 (2009).
Gonzalez, I., Cao, K. A., Davis, M. J. & Dejean, S. Visualising associations between paired ‘omics’ data sets. BioData Min. 5, 19-0381-5-19 (2012).
Le Cao, K. A., Gonzalez, I. & Dejean, S. integrOmics: an R package to unravel relationships between two omics datasets. Bioinformatics 25, 2855–2856 (2009).
Acknowledgements
The authors would like to thank Jenny Jansen (Division of Human Nutrition, Wageningen University) for technical support in the microarray analysis. This work was supported by a project from the Top Institute Food and Nutrition (grant GH002), Wageningen, The Netherlands. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: M.E., F.H., C.B., M.B., P.d.V., M.F. Performed the experiments: M.E., F.H., B.d.H. Analyzed the data: M.E., F.H. Wrote the paper: M.E., F.H., P.d.V., M.F.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Elderman, M., Hugenholtz, F., Belzer, C. et al. Changes in intestinal gene expression and microbiota composition during late pregnancy are mouse strain dependent. Sci Rep 8, 10001 (2018). https://doi.org/10.1038/s41598-018-28292-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-28292-2
This article is cited by
-
A comprehensive analysis of the microbiota composition and gene expression in colorectal cancer
BMC Microbiology (2020)
-
Reproductive hormones mediate changes in the gut microbiome during pregnancy and lactation in Phayre’s leaf monkeys
Scientific Reports (2020)
-
Of men in mice: the development and application of a humanized gnotobiotic mouse model for microbiome therapeutics
Experimental & Molecular Medicine (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
