
Abstract
Blood vessels in the CNS form a specialized and critical structure, the blood–brain barrier (BBB). We present a resource to understand the molecular mechanisms that regulate BBB function in health and dysfunction during disease. Using endothelial cell enrichment and RNA sequencing, we analyzed the gene expression of endothelial cells in mice, comparing brain endothelial cells with peripheral endothelial cells. We also assessed the regulation of CNS endothelial gene expression in models of stroke, multiple sclerosis, traumatic brain injury and seizure, each having profound BBB disruption. We found that although each is caused by a distinct trigger, they exhibit strikingly similar endothelial gene expression changes during BBB disruption, comprising a core BBB dysfunction module that shifts the CNS endothelial cells into a peripheral endothelial cell-like state. The identification of a common pathway for BBB dysfunction suggests that targeting therapeutic agents to limit it may be effective across multiple neurological disorders.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
The data that support the findings of this study are available from the corresponding author upon request. RNA-sequencing files were deposited in the Gene Expression Omnibus repository and are available to use. Specific experimental details are also organized in the accompanying Nature Research Reporting Summary.
References
Daneman, R. The blood–brain barrier in health and disease. Ann. Neurol. 72, 648–672 (2012).
Zlokovic, B. V. The blood–brain barrier in health and chronic neurodegenerative disorders. Neuron 57, 178–201 (2008).
Rubin, L. L. & Staddon, J. M. The cell biology of the blood–brain barrier. Annu. Rev. Neurosci. 22, 11–28 (1999).
Janzer, R. C. & Raff, M. C. Astrocytes induce blood–brain barrier properties in endothelial cells. Nature 325, 253–257 (1987).
Stewart, P. A. & Wiley, M. J. Developing nervous tissue induces formation of blood–brain barrier characteristics in invading endothelial cells: a study using quail-chick transplantation chimeras. Dev. Biol. 84, 183–192 (1981).
Armulik, A. et al. Pericytes regulate the blood–brain barrier. Nature 468, 557–561 (2010).
Daneman, R., Zhou, L., Kebede, A. A. & Barres, B. A. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature 468, 562–566 (2010).
Rubin, L. L. et al. A cell culture model of the blood–brain barrier. J. Cell Biol. 115, 1725–1735 (1991).
Weidenfeller, C., Svendsen, C. N. & Shusta, E. V. Differentiating embryonic neural progenitor cells induce blood–brain barrier properties. J. Neurochem. 101, 555–565 (2007).
Daneman, R. et al. Wnt/beta-catenin signaling is required for CNS, but not non-CNS, angiogenesis. Proc. Natl Acad. Sci. USA 106, 641–646 (2009).
Liebner, S. et al. Wnt/beta-catenin signaling controls development of the blood–brain barrier. J. Cell Biol. 183, 409–417 (2008).
Stenman, J. M. et al. Canonical Wnt signaling regulates organ-specific assembly and differentiation of CNS vasculature. Science 322, 1247–1250 (2008).
Wang, Y. et al. Norrin/Frizzled4 signaling in retinal vascular development and blood brain barrier plasticity. Cell 151, 1332–1344 (2012).
Zhou, Y. & Nathans, J. Gpr124 controls CNS angiogenesis and blood–brain barrier integrity by promoting ligand-specific canonical Wnt signaling. Dev. Cell 31, 248–256 (2014).
Zhou, Y. et al. Canonical WNT signaling components in vascular development and barrier formation. J. Clin. Invest. 124, 3825–3846 (2014).
Pinzon-Daza, M. L. et al. The cross-talk between canonical and non-canonical Wnt-dependent pathways regulates P-glycoprotein expression in human blood–brain barrier cells. J. Cereb. Blood Flow Metab. 34, 1258–1269 (2014).
Ioannidou, S. et al. An in vitro assay reveals a role for the diaphragm protein PV-1 in endothelial fenestra morphogenesis. Proc. Natl Acad. Sci. USA 103, 16770–16775 (2006).
Keuschnigg, J. et al. The prototype endothelial marker PAL-E is a leukocyte trafficking molecule. Blood 114, 478–484 (2009).
Shue, E. H. et al. Plasmalemmal vesicle associated protein-1 (PV-1) is a marker of blood–brain barrier disruption in rodent models. BMC Neurosci. 9, 29 (2008).
Doring, A., Wild, M., Vestweber, D., Deutsch, U. & Engelhardt, B. E- and P-selectin are not required for the development of experimental autoimmune encephalomyelitis in C57BL/6 and SJL mice. J. Immunol. 179, 8470–8479 (2007).
Love, S. & Barber, R. Expression of P-selectin and intercellular adhesion molecule-1 in human brain after focal infarction or cardiac arrest. Neuropathol. Appl. Neurobiol. 27, 465–473 (2001).
Okada, Y. et al. P-selectin and intercellular adhesion molecule-1 expression after focal brain ischemia and reperfusion. Stroke 25, 202–211 (1994).
Mayadas, T. N., Johnson, R. C., Rayburn, H., Hynes, R. O. & Wagner, D. D. Leukocyte rolling and extravasation are severely compromised in P selectin-deficient mice. Cell 74, 541–554 (1993).
Huang, J. et al. Postischemic cerebrovascular E-selectin expression mediates tissue injury in murine stroke. Stroke 31, 3047–3053 (2000).
Jin, A. Y. et al. Reduced blood brain barrier breakdown in P-selectin deficient mice following transient ischemic stroke: a future therapeutic target for treatment of stroke. BMC Neurosci. 11, 12 (2010).
Bowes, M. P., Zivin, J. A. & Rothlein, R. Monoclonal antibody to the ICAM-1 adhesion site reduces neurological damage in a rabbit cerebral embolism stroke model. Exp. Neurol. 119, 215–219 (1993).
Connolly, E. S. Jr. et al. Cerebral protection in homozygous null ICAM-1 mice after middle cerebral artery occlusion. Role of neutrophil adhesion in the pathogenesis of stroke. J. Clin. Invest. 97, 209–216 (1996).
Henninger, D. D. et al. Cytokine-induced VCAM-1 and ICAM-1 expression in different organs of the mouse. J. Immunol. 158, 1825–1832 (1997).
Lyck, R. et al. T-cell interaction with ICAM-1/ICAM-2 double-deficient brain endothelium in vitro: the cytoplasmic tail of endothelial ICAM-1 is necessary for transendothelial migration of T cells. Blood 102, 3675–3683 (2003).
Sharief, M. K., Noori, M. A., Ciardi, M., Cirelli, A. & Thompson, E. J. Increased levels of circulating ICAM-1 in serum and cerebrospinal fluid of patients with active multiple sclerosis. Correlation with TNF-ɑ and blood–brain barrier damage. J. Neuroimmunol. 43, 15–21 (1993).
Derada Troletti, C., de Goede, P., Kamermans, A. & de Vries, H. E. Molecular alterations of the blood–brain barrier under inflammatory conditions: the role of endothelial to mesenchymal transition. Biochim. Biophys. Acta 1862, 452–460 (2016).
Zhou, T. et al. Microvascular endothelial cells engulf myelin debris and promote macrophage recruitment and fibrosis after neural injury. Nat. Neurosci. 22, 421–435 (2019).
Ley, K. The role of selectins in inflammation and disease. Trends Mol. Med. 9, 263–268 (2003).
Conductier, G., Blondeau, N., Guyon, A., Nahon, J. L. & Rovere, C. The role of monocyte chemoattractant protein MCP1/CCL2 in neuroinflammatory diseases. J. Neuroimmunol. 224, 93–100 (2010).
Aggarwal, B. B., Shishodia, S., Ashikawa, K. & Bharti, A. C. The role of TNF and its family members in inflammation and cancer: lessons from gene deletion. Curr. Drug Targets Inflamm. Allergy 1, 327–341 (2002).
Xing, C. et al. Neuronal production of lipocalin-2 as a help-me signal for glial activation. Stroke 45, 2085–2092 (2014).
Ferreira, A. C. et al. From the periphery to the brain: lipocalin-2, a friend or foe? Prog. Neurobiol. 131, 120–136 (2015).
Cheng, B., Chen, J., Bai, B. & Xin, Q. Neuroprotection of apelin and its signaling pathway. Peptides 37, 171–173 (2012).
Ma, S., Kwon, H. J., Johng, H., Zang, K. & Huang, Z. Radial glial neural progenitors regulate nascent brain vascular network stabilization via inhibition of Wnt signaling. PLoS Biol. 11, e1001469 (2013).
Sun, C. et al. Conditional ablation of neuroprogenitor cells in adult mice impedes recovery of poststroke cognitive function and reduces synaptic connectivity in the perforant pathway. J. Neurosci. 33, 17314–17325 (2013).
Akamatsu, Y. et al. Impaired leptomeningeal collateral flow contributes to the poor outcome following experimental stroke in the type 2 diabetic mice. J. Neurosci. 35, 3851–3864 (2015).
Semple, B. D. et al. Early gelatinase activity is not a determinant of long-term recovery after traumatic brain injury in the immature mouse. PLoS One 10, e0143386 (2015).
Semple, B. D., Blomgren, K., Gimlin, K., Ferriero, D. M. & Noble-Haeusslein, L. J. Brain development in rodents and humans: identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 106-107, 1–16 (2013).
Daneman, R. et al. The mouse blood–brain barrier transcriptome: a new resource for understanding the development and function of brain endothelial cells. PLoS One 5, e13741 (2010).
Lui, J. H. et al. Radial glia require PDGFD-PDGFRβ signalling in human but not mouse neocortex. Nature 515, 264–268 (2014).
Langfelder, P. & Horvath, S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics 9, 559 (2008).
Oldham, M. C. et al. Functional organization of the transcriptome in human brain. Nat. Neurosci. 11, 1271–1282 (2008).
Acknowledgements
We would like to thank N. Lescano and A. Schroeder and the Parnassus Flow Cytometry Core, T. Rambaldo and the UCSD Veterans Hospital Flow Cytometry Core, R. Chadwick and the Gladstone Genomics Core, A. Williams and The Gladstone Bioinformatics Core and K. Jepsen and the UCSD Institute for Genomics Medicine Genomics Center. R.D. is funded by the National Institutes of Health (NIH)/National Institue of Neurological Disorders and Stroke (NINDS) (grant no. R01 NS091281-01A1), a National Multiple Sclerosis Society Pilot Grant and Takeda Pharmaceuticals New Frontier Science Program. R.N.M. was funded by the UCSF Department of Clinical Pharmacology and Therapeutics (grant no. GM007546) and the Department of Anesthesia and Perioperative Care (grant no. GM008440) NIH T32 grants. M.C.O. is funded by the NIH/National Institue of Mental Health (grant no. R01 MH113896). L.J.N.-H. is funded by the NIH/NINDS (grant nos. R01 NS050159 and NS077767). T.H. is funded by the NIH/NINDS (grant nos. R01NS055876 and R01NS082280) and the Barrow Neurological Foundation grant.
Author information
Authors and Affiliations
Contributions
R.D. designed and participated in the analysis of all experiments. R.N.M. designed the seizure and Wnt signaling studies, performed cell purification of health and Wnt signaling models, and participated in the analysis of health, disease and Wnt signaling studies and writing of the manuscript. A.L.S. performed the endothelial cell purification and analysis of health and disease models. G.A.W. performed the alignment and normalization of RNA-sequencing data. P.G.S. and M.C.O. designed and performed the gene coexpression analysis. F.S. participated in the design and analysis of the EAE model. L.J.N.-H., B.D.S., A.T. and K.G. participated in the design and analysis of all TBI experiments. T.H., M.Korai and M.Kotoda participated in the design and analysis of all MCAO experiments. S.A. participated in the design and analysis of the Wnt signaling study. A.B. and A.C.C. participated in the design and analysis of disease models.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Neuroscience thanks Britta Engelhardt and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 Cell markers in endothelial cell samples.
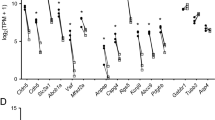
Comparison of the expression levels of endothelial cell markers and non-endothelial cell markers in endothelial cells enriched from brain tissue. A) The brain vascular (black bars; samples-red dots) and brain endothelial samples (gray bars; samples-blue dots) are enriched with RNA of markers for CNS endothelial cells but not RNA of markers for CNS pericytes, astrocytes, neurons, oligodendrocytes (oligoden) and microglia (micrg). B) A graph representing the same data in A with the y-axis set to 220CPM and excluding the highly enriched endothelial cell markers to demonstrate the lack of enrichment of pericyte cell marker expression in the brain endothelial samples compared to the brain vascular samples. A,B) Data is presented as mean ±SEM; n=2 mice per condition.
Supplementary Figure 2 Quantification of blood-brain barrier biotin permeability.
Quantified pixel intensity of biotin:streptavidin-alexa-488 fluorescence on brain/spinal cord tissue sections from seizure, EAE, stroke and TBI disease models and corresponding controls during acute, subacute and chronic timepoints. Biotin was intravenously perfused in anesthetized mice. Data is presented as median fold change of control with arbitrary units (AU). Box plot: 25th–75th percentile box, minimum-maximum whiskers. Statistical test: t test (unpaired, parametric, equal standard deviation, two-tailed), * P-value<0.05, ** P-value<0.01, *** P-value<0.001, **** P-value<0.0001. n=number of mice (Control/Acute/Subacute/Chronic): Seizure: 5/3/4/4, EAE: 4/4/5/5, Stroke: 9/3/3/3, TBI: 8/3/3/3.
Supplementary Figure 3 Quantification of blood-brain barrier fibrinogen permeability.
A-H) Representative images of brain/spinal cord sections immunostained for the endogenous serum component fibrinogen (red) and vessel marker CD31 (green) in control and subacute mice. Fibrinogen is increased in seizure, EAE, stroke and TBI disease models (B,D,F,H) compared to controls (A,C,E,G). B/D/F/H-prime) Higher magnification of boxed regions in subacute showing colocalization of fibrinogen with CD31+ vessels (arrowheads highlight examples). Scale bars: 100 microns; 200 microns in E and G. n=number of independent experiments: Seizure: 2, EAE:2, Stroke: 3, TBI: 4. I) Quantified pixel intensity of fibrinogen immunostaining with alexa-594 fluorophore on tissue sections from seizure, EAE, stroke and TBI disease models and corresponding controls during the subacute timepoint. Data is presented as median fold change of control with arbitrary units (AU). Box plot: 25th-75th percentile box, minimum-maximum whiskers. Statistical test: t test (unpaired, parametric, equal standard deviation, two-tailed), ** P-value<0.01, *** P-value<0.001. n=number of mice (Control/Subacute): Seizure: 3/4, EAE: 4/3 Stroke: 4/4, TBI: 3/3.
Supplementary Figure 4 Immunoglobulin G localization in the CNS.
Representative images of brain/spinal cord sections immunostained for endogenous immunoglobulin G (IgG) in control and subacute mice. Peripherally synthesized IgG is increased in seizure, EAE, stroke and TBI disease models (B,D,F,H) compared to controls (A,C,E,G). Scale bars: 200 microns; 100 microns in EAE. n=number of mice (Control/Subacute): Seizure: 3/5, EAE: 2/3, Stroke: 4/4, TBI: 3/3.
Supplementary Figure 5 Claudin 5 organization in CNS vessels.
Representative images of brain/spinal cord sections immunostained for the tight junction protein Caludin 5 (red) and vessel marker CD31 (green) in control and subacute mice. Claudin 5 localization in vessels is disorganized in hypertrophic vessels of EAE, stroke and TBI disease models (D,F,H) compard to controls (C,E,G) (arrowheads highlight examples). Hypertrophic vessels in the seizure model was observed but was less common. Scale bar: 100 microns. n=number of mice (Control/Subacute): Seizure: 3/7, EAE: 3/3, Stroke: 4/4, TBI: 3/3.
Supplementary Figure 6 Expression of collagen I in the CNS.
Representative images of brain/spinal cord sections immunostained for extracellular matrix protein collagen I (red) and vessel marker CD31 (green) in control and subacute mice. Collagen I colocalized with CD31 is increased in seizure, EAE, stroke and TBI disease models (B,D,F,H) compared to controls (A,C,E,G). Arrow in seizure model control (A) highlight an example of a medium size vessel without collagen expression. Dense DAPI labeling in EAE subacute identify a lesion. Arrowheads highlight examples of ectopic expression of collagen I colocalized with CD31+ vessels. Scale bars: 50 microns. n=number of mice (Control/Subacute): Seizure: 3/4, EAE: 4/3, Stroke: 4/4, TBI: 3/3.
Supplementary Figure 7 Quantification of the expression of BBB dysfunction module genes.
Quantified vessel length expressing the gene of interest (GOI). Expression of extracellular matrix genes collagen I, collagen III, decorin, lumican and SPP1 is increased in brain/spinal cord from seizure, EAE, stroke and TBI disease models compared to corresponding controls. Data is presented as mean percent vascular length (±SEM) expressing the GOI of the total length of CD31 positive vessels. Statistical test: t test (equal standard deviation, two-tailed, unpaired: Seizure and EAE, paired ipsilateral/contralateral: MCAO and TBI), * P-value<0.05, ** P-value<0.01, *** P-value<0.001, **** P-value<0.0001. n=number of mice (Control/Subacute): Seizure: 3/3, EAE: 3/3, Stroke: 4/4, TBI: 3/3.
Supplementary Figure 8 Endothelial cell enrichment.
A graphical representation of the key steps of the endothelial cell enrichment procedure used in this study. The regions of interests in health or disease experiments for brain, heart, lung, liver, kidney or spinal cord are identified in box A. The blue flow path (B) is the standard procedure. The violet flow path (C) adds the second enzymatic digestion step to dissociate remaining adherent cells used in the health “Brain endothelial” samples. The red flow path (D) adds the modifications for all peripheral organs. The green flow path (E) further adds modifications to the peripheral organ procedures for the activated beta-catenin studies.
Supplementary Figure 9 FACS gating strategy for endothelial cell enrichment.
Representative sample. (A) Gating for single cells with side scatter (SSC) and forward scatter (FSC). (B) Gating to exclude dead cells (DAPI+), and immune cells and pericytes (A488+ or FITC+). (C) Gating for endothelial cells (tdTomato+). (D) Number of events detected and percent of events relative to parent population (All Events) or previous gate.
Supplementary information
Supplementary Figs. 1–9, Supplementary Results & Discussion and Supplementary Tables 1–5
Supplementary Figs. 1–9, Supplementary Results & Discussion and Supplementary Tables 1–5.
Supplementary File 1
RNA sequencing of brain endothelial cells in health and disease. Master excel spreadsheet containing the expression values in counts per million (c.p.m.) for each annotated gene in samples of brain vascular cells, brain endothelial cells, heart endothelial cells, kidney endothelial cells, lung endothelial cells, liver endothelial cells and whole brain, as well as control, acute, subacute and chronic timepoints in CNS endothelial cells following the kainic acid model of seizures, EAE model of multiple sclerosis, MCAO model of stroke and impact model of pediatric TBI. The log2 fold comparisons are given for each gene for critical pair-wise comparisons in health and for each disease. Statistics are displayed on additional tabs: The P value and false discovery rate (FDR) values are given for the critical comparisons in health (second tab, M-AU) and diseases (third tab, E-CC). For each gene we also identify whether it is enriched given comparisons in health (second tab, AW-BM) and disease (third tab, CE-DJ). In the fourth tab we present relevant metrics from gene coexpression network analysis. ‘Seed’ genes (column C) met statistical criteria for initial module assignments (Methods). After identifying coexpression modules, module membership strength (kME) was calculated for all genes with respect to all modules. Columns H-BW report the kME value and corresponding nominal P value for each gene with respect to each module eigengene. Columns F and G indicate the module with the most significant, positive kME value for that gene meeting either Bonferroni- (column F) or false discovery rate- (column G) corrected levels of significance. We also report the mean expression value for each gene over all samples (column D), as well as its percentile rank (column E). Statistical tests: Wald test for P values and Benjamini–Hochberg for FDR. n = 2 mice each condition for healthy organs as source of enriched endothelial cells and 4 mice for whole brain tissue. n = 3 mice each for disease model control and experimental conditions as source of enriched endothelial cells with exception of n = 2 mice for TBI control subacute and chronic conditions.
Supplementary File 2
Mural cell genes. The list of brain mural cell-enriched genes identified as those genes enriched in the brain vascular over brain endothelial samples (BV c.p.m. > 5, log2 fold > 1.5, P < 0.05, BE < 10 c.p.m.). Statistical tests: Wald test for P values and Benjamini–Hochberg for FDR. n = 2 mice each condition as source of enriched endothelial cells and 4 mice for whole brain tissue.
Supplementary File 3
BBB-enriched genes. The list of BBB-enriched genes identified as genes with at least 5 counts per million (c.p.m.) in brain endothelial cells, and at log2 fold > 1.00 enrichment in brain endothelial cells compared with each peripheral endothelial cell with P < 0.05. Mural cell genes were excluded by identifying genes with brain vascular c.p.m. > brain endo c.p.m., log2 fold > 1.00, brain vascular c.p.m. > 5, P < 0.05). Statistical tests: Wald test for P values and Benjamini–Hochberg for FDR. n = 2 mice each condition as source of enriched endothelial cells and 4 mice for whole brain tissue.
Supplementary File 4
Tight junction proteins expressed in brain endothelial cells. Excel spreadsheet of the tight junction proteins (bicellular and tricellular), as annotated by mouse genome informatics (MGI), with at least 5 c.p.m. expression value in the brain endothelial cell sample. Statistical test: Wald test. n = 2 mice each condition as source of enriched endothelial cells and 4 mice for whole brain tissue.
Supplementary File 5
Peripheral endothelial-enriched genes. The list of peripheral endothelial-enriched genes identified as those genes enriched (c.p.m. > 5, log2 fold > –1.00, P < 0.05) in at least three of the peripheral endothelial samples compared with the brain endothelial cells. Statistical tests: Wald test for P values and Benjamini–Hochberg for FDR. n = 2 mice each condition as source of enriched endothelial cells and 4 mice for whole brain tissue.
Supplementary File 6
BBB dysfunction module. The list of genes that are upregulated (log2 fold > 1.00, c.p.m. > 5 at subacute timepoint, P < 0.05) in at least three of the four diseases at the subacute timepoint. Statistical tests: Wald test for P values and Benjamini–Hochberg for FDR. n = 3 mice each condition as source of enriched endothelial cells with exception of n = 2 mice for TBI control subacute and chronic conditions.
Supplementary File 7
RNA sequencing of peripheral endothelial cells following activated beta-catenin signaling. Master excel spreadsheet containing the expression values in counts per million (c.p.m.) for each annotated gene in samples taken from liver and lung endothelial cells of control (VE-Cadherin-CreERT2) and mice with activated beta-catenin induced in their endothelial cells (Rosa-Bcat-GOF; VE-Cadherin-CreERT2). log2 fold comparisons, P values and false discovery rate (FDR) values are given for each gene comparing controls with activated beta-catenin conditions of liver and lung samples. Additional tabs are given listing genes that are up- and downregulated due to activated beta-catenin in the liver endothelial cells, lung endothelial cells and both as given by P < 0.05 and an absolute value of >10 c.p.m. in the sample with the greater value. Statistical tests: Wald test for P values and Benjamini–Hochberg for FDR. n = 4 mice each condition as source of enriched endothelial cells.
Rights and permissions
About this article

Cite this article
Munji, R.N., Soung, A.L., Weiner, G.A. et al. Profiling the mouse brain endothelial transcriptome in health and disease models reveals a core blood–brain barrier dysfunction module. Nat Neurosci 22, 1892–1902 (2019). https://doi.org/10.1038/s41593-019-0497-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41593-019-0497-x
This article is cited by
-
A brain-specific angiogenic mechanism enabled by tip cell specialization
Nature (2024)
-
LET-381/FoxF and its target UNC-30/Pitx2 specify and maintain the molecular identity of C. elegans mesodermal glia that regulate motor behavior
The EMBO Journal (2024)
-
Teriflunomide Promotes Blood–Brain Barrier Integrity by Upregulating Claudin-1 via the Wnt/β-catenin Signaling Pathway in Multiple Sclerosis
Molecular Neurobiology (2024)
-
Chronic social defeat alters brain vascular-associated cell gene expression patterns leading to vascular dysfunction and immune system activation
Journal of Neuroinflammation (2023)
-
ENGEP: advancing spatial transcriptomics with accurate unmeasured gene expression prediction
Genome Biology (2023)
