
Abstract
This study investigates a contact zone between two silverside fish species (marine Odontesthes argentinensis and freshwater O. bonariensis) in the estuarine Mar Chiquita lagoon along the Atlantic coast in Argentina (MChL), in which intermediate morphs had been reported. It has been suggested that admixture and introgression occur in MChL between these two species, but direct genetic evidence is lacking. Leveraging samples collected over several years (n = 676), we document the spatial distribution of both species and intermediate morphs within this habitat and collect landmark-based morphometric and multilocus genetic data (9876 loci for n = 110 individuals) to test the hypothesis of hybridization. Our analysis unambiguously characterizes intermediate morphs as F1 or F2 hybrids. We show that the low frequency of hybrid individuals in MChL may be explained by uneven abundance of parental species, which in turn are strongly affected by water salinity, limiting the size of the contact zone. Although hybrids seem to be fertile, their fitness may be reduced by external and intrinsic factors that may limit their success and suggest that this is an unstable hybrid zone. Genetic distinctiveness of both parental species is strongly supported by genome-wide data, explaining a known pattern of mitonuclear discordance as a consequence of hybridization followed by mitochondrial introgression. A clear signature of population genetic structure was detected in O. argentinensis, distinguishing MChL residents from marine populations of this species, that also was supported by distinctive morphometric features among these groups. Previous hypotheses of speciation in these fishes are discussed in the light of the new findings.
Similar content being viewed by others

Introduction
Hybridization between closely related species is ubiquitous in nature, with both ecological and behavioral factors playing a role in the process (Taylor and Larson 2019). Hybrid zones form when the ranges of two species or diverged populations meet and cross-fertilize. Within this zone, it is common to see the breakdown of secondary properties defining the species category (de Queiroz 2007), such as overlaps of ecological niches, phenotype, diet, or the failure of assortative mating (Montanari et al. 2016). In the context of speciation and the maintenance of isolating mechanisms, hybridization may have several distinct outcomes determined by the fitness of hybrid individuals (Barton and Hewitt 1985). Unstable hybrid zones are defined when hybrids have lower fitness than pure parental individuals, and are only maintained by continuous invasion into the area of parental types (Moore and Buchanan 1985). Although some level of cross-species reproduction takes place, parental species remain differentiated. Strong natural selection can relegate hybrid individuals to marginal or intermediate habitats even under conditions of relatively high gene flow (DiVittorio et al. 2020), and hybrids may have reduced fertility or be sterile. But if natural selection is weak or shifts due to a change in the environment and hybrids are fertile, species boundaries may collapse entirely (Taylor et al. 2006). Therefore, the strength of selection on hybrids has a significant effect on the outcome of hybridization. Alternatively, stable hybrid zones occur when hybrid offspring have higher fitness than either parental type under intermediate ecological conditions, potentially leading to hybrid speciation. Most of the described animal hybrid species succeed by occupying available ecological space unused by the parental taxa. The outcomes of hybridization in a stable hybrid zone, however, also may differ when environmental conditions fluctuate over time and space (Rhymer and Simberloff 1996; Todesco et al. 2016; Zhang et al. 2019). Estuaries are uniquely dynamic ecosystems of brackish water that form transition zones connecting maritime and freshwater environments. They usually have high productivity, serve as mating or nursery grounds for aquatic organisms (Sheaves et al. 2015), and often hold secondary contact zones for species pairs adapted to divergent salinities, providing opportunities for hybridization.
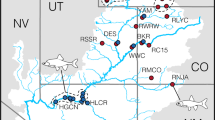
The Mar Chiquita Coastal Lagoon (MChL) is a shallow estuary along the South Atlantic coast of Buenos Aires Province (Argentina) with approximate surface area of 46 km² that developed after fluctuations in sea level during the Holocene formed a sandy barrier with a narrow inlet (Fig. 1). The lagoon continues to be shaped by littoral drift, winds, and vegetation. It serves as an important nursery, feeding and reproduction area for several fish species (González-Castro et al. 2009, 2016, 2019). Its salinity fluctuates substantially between 0 and 36 ppt, shifting due to the influx of fresh water into the lagoon, the tides, and the direction and intensity of the wind. The unusual distribution in this lagoon of three species of silverside fishes in the genus Odontesthes (family Atherinopsidae, locally known as “pejerrey”) offers an opportunity to study the effects of secondary and putative hybridization across a strong environmental gradient. Two species in particular, the freshwater species Odontesthes bonariensis and the marine species O. argentinensis are of interest due to their local importance in aquaculture and sport fishing (Somoza et al. 2008; González-Castro et al. 2016). A third species, the marine silverside O. platensis is thought to be more distantly related to the other two (García et al. 2014; González-Castro et al. 2019) and only is present in MChL occasionally (González-Castro et al. 2009).

a Map of sample localities outside of the Mar Chiquita Coastal Lagoon (see Table S1 for description of population codes). b Map of sampling locations within MChL. The approximate surface area of the lagoon is 46 km2.
Odontesthes bonariensis is widely distributed in freshwater habitats (rivers, lakes, and lagoons) of the Pampas region of Argentina, in the Río de la Plata, Lower Paraná, and Uruguay river basins, and in coastal Atlantic drainages from southern Buenos Aires Province in Argentina to the Dos Patos coastal lagoon system in Brazil (Dyer 2006; Somoza et al. 2008). Odontesthes argentinensis is a marine species that ranges from the South Atlantic coast of Chubut Province in Argentina to São Paulo State in Brazil. These closely-related taxa are distinguishable on the basis of morphological and meristic traits, but molecular analysis based on mitochondrial DNA (mtDNA) failed to find genetic differences between these species (García et al. 2014; Calvelo and D’Anatro 2021). In contrast, a comparative transcriptomic study revealed 1417 transcripts with putatively fixed non-synonymous differences (SNPs), plus even a larger number (>3000) of differentially expressed transcripts in the gills of these two species (Hughes et al. 2017). Phylogenetic analysis of a genome-wide dataset of restriction-enzyme associated DNA (RAD) resolved the relationships among species of Odontesthes with high confidence (Hughes et al. 2020), placing marine O. argentinensis as the sister-group of a clade of freshwater species with partially overlapping ranges that include O. bonariensis. Most importantly, the RAD dataset resolved both species (O. argentinensis and O. bonariensis) as reciprocally monophyletic groups, suggesting that lack of differentiation in mtDNA may be due to introgression following hybridization with limited effects on the genome of these species. But studies identifying and characterizing hybrids between these species are lacking. Throughout most of their distribution, O. bonariensis and O. argentinensis are found to coexist in brackish waters found in river estuaries and coastal lagoons. Phenotypically intermediate individuals between the freshwater O. bonariensis and marine O. argentinensis are found in the MChL, and were hypothesized to be hybrids as a result of secondary contact between these two species (González-Castro et al. 2016). These intermediate morphotypes were clearly separated from either species by meristic and morphometric traits. González-Castro et al. (2016) also reported that the most abundant pejerrey in MChL can be identified as a “resident population” of O. argentinensis, readily distinguished by morphometric analysis from the purely marine populations of this species. Individuals of O. argentinensis from the adjacent oceanic coast enter the lagoon only under certain environmental conditions, and are restricted to the mouth of the lagoon where salinity is high. A similar pattern was described for this species in other estuaries in Southern Brazil (Patos and Cassino Lagoons) based on allozyme, mtDNA, and microsatellite data (Beheregaray and Levy 2000; Beheregaray and Sunnucks 2001). The distinction between marine and resident estuarine populations also has been shown to involve reproductive behavior (spawning site selection) and egg morphology (Phonlor and Cousin 1997; Moresco and Bemvenuti 2006), and some authors went as far as to suggest that the estuarine forms of O. argentinesis may represent incipient species that are reproductively isolated from the marine stock through a process of “divergence with gene flow” (Beheregaray and Sunnucks 2001; González-Castro et al. 2019). In contrast to O. argentinenis, the abundance of O. bonariensis in MChL is quite low and restricted to the innermost, mixo-oligohaline zone of MChL and its freshwater tributaries, where the salinity is low (González-Castro et al. 2009). The extent of genetic differentiation between marine and estuarine populations of O. argentinensis in MChL and the extent of presumed hybridization between this species and O. bonariensis are not known. Besides description of intermediate morphotypes (González-Castro et al. 2016), the nature of a putative hybrid zone, including the fitness of hybrids and environmental factors affecting them, still needs to be characterized.
In this study, we compile and analyze RAD sequence data together with morphological and spatial data (abundance, locality, and salinity) in order to characterize the presumed Odontesthes hybrid zone in the MChL, with the goal of determining potential gene flow between species and between marine and resident populations of O. argentinensis. Based on the environmental data, we investigate the effect of fluctuating salinity within MChL on the distribution of species that determine the extent of secondary contact. We assess the genetic composition and distribution of intermediate individuals, and determine their reproductive status. We include in our comparisons putative pure parental O. argentinensis and O. bonariensis specimens collected in other localities. Finally, we characterize genetic and morphometric differentiation between O. argentinensis resident in MChL and those from adjacent coastal marine environments to better understand the evolutionary status of resident estuarine populations.
Material and methods
Taxonomic sampling
Specimens sampled for this study are listed in Table S1 and their sampling localities are shown in Fig. 1. Fish collected inside the MChL, and in other coastal and freshwater environments were identified as species using meristic counts and other morphological features (González-Castro et al. 2016) and placed into one of five groups: (i) Odontesthes bonariensis from the MChL and from other freshwater localities; (ii) Odontesthes argentinensis from the MChL (considered “residents” of the lagoon12); (iii) Odontesthes argentinensis from adjacent and more distant marine localities; (iv) Putative hybrids, referred to as MChL “morphotypes” in (González-Castro et al. 2016); (v) Odontesthes platensis: from the MChL and from an adjacent coastal locality. The number of individuals assigned to each group for the molecular and/or morphometrics analyses is shown in Table S1.
RAD sequencing and analysis
Total DNA was extracted using a 96-well plate format on a Gene Prep (Autogen), following manufacturer’s instructions, at the Laboratory of Analytic Biology at the National Museum of Natural History (Smithsonian Institution, Washington DC). Genomic libraries were generated following the original double digest RAD (ddRAD) sequencing protocol (Peterson et al. 2012) at the University of Virginia Sequencing and Genomics Facility, modified to use MseI and PstI restriction enzymes and a 350–550 bp size selection to be compatible with previous ddRADseq data generated for Odontesthes (Hughes et al. 2020). Paired-end libraries were sequenced on two lanes of an Illumina HiSeq 4000 at the University of Chicago Genomics Facility. Sequences were demultiplexed, quality filtered, and assembled into ddRAD loci using ipyrad v. 0.7.30 (Eaton 2014) using the settings shown in the supplementary information. We compiled five different datasets (Table S2). Dataset 1 includes 110 specimens from all five groups (O. bonariensis, MChL resident and marine O. argentinensis, putative hybrids, and O. platensis), plus published data (Hughes et al. 2020) for outgroup taxa O. incisa, O. smitti, and O. hatcheri (Table S1 and Fig. 1). This dataset was used to infer a phylogeny and assess the genetic affinities of putative hybrids. Dataset 2 includes O. bonariensis, O. argentinensis (MchL residents and marine), and putative hybrids; Dataset 3 only O. argentinensis (MchL residents and marine) and putative hybrids, and both were used to test hybridization and population structure. Dataset 4 included only specimens of O. argentinensis (MchL residents and marine) and dataset 5 only specimens of O. bonariensis and were used to test population structure within each species.
Concatenated ddRAD loci in Dataset 1 were analyzed under Maximum Likelihood using IQ-Tree (Nguyen et al. 2015). We selected the best-fitting model with ModelFinder (Kalyaanamoorthy et al. 2017) and generated 1000 UFBoot replicates to assess branch support (Hoang et al. 2018). Concatenated ddRAD loci of datasets 2–5 also were analyzed using a phylogenetic network approach in SplitsTree4 v.4.17.1 (Huson and Bryant 2006). We estimated the networks using the best-fitting model, Neighbour-Net algorithm, and statistical support for branches were calculated using 1000 replicates (Table S2).
To detect potential admixture and population structure in our samples, the software fastStructure v1.0 (Raj et al. 2014) wrapped in Structure_threader v0.4.3 (Pina-Martins et al. 2017) was used with K values ranging from 1–7 using a logistic prior. The optimal K value was found using the chooseK.py script bundled in Structure_threader. A Principal Components Analysis (PCA) on SNPs was conducted using the ALStructure (Cabreros and Storey 2019) after excluding non-biallelic loci using the snpgds PCA function in Structure_threader. The results were visualized using the distruct.py package (Raj et al. 2014). This analysis was performed using Datasets 2–5 in order to examine structure and admixture in the species that may contribute to the hybridization process in the MChL.
Analysis of hybridization
Dataset 2 was used to test the extent of hybridization between O. bonariensis and O. argentinensis within the MChL. We used the R package gghybrid to estimate the hybrid index (HI) using 10,000 MCMC generations and estimated posterior probability values after a 5000-generation burn-in (Bailey 2018). We selected the samples of O. argentinensis and O. bonariensis from outside the MChL as pure parental references. Hybridization was also assessed using the program NewHybrids (Anderson and Thompson 2002). Posterior probabilities were calculated for six genotype classes: pure O. argentinensis, pure O. bonariensis, F1 hybrid, F2 hybrid, or backcrosses with either O. argentinensis or O. bonariensis. Two independent runs were conducted with 1,000,000 sweeps following a 100,000 burn-in period, using Jeffreys-like priors and mixing proportions. We then run the R package HybridDetective (Wringe et al. 2017) to estimate the statistical confidence assignments made with NewHybrid. To examine convergence, simulations were run across three replicates, each with three independent MCMC chains performed with the function freqbasedsim_AlleleSample. Final runs were used to categorize individuals to genealogical class, using a posterior probability threshold of 0.90. Also, for each individual, we calculated a maximum likelihood estimate of the genomic hybrid index and the average heterozygosity across ancestry-informative loci using R package Introgress (Gompert and Buerkle 2010). F1 hybrids have an expected hybrid index of 0.5 and expected heterozygosity of 1.0 for loci fixed in parental individuals. Heterozygosity is reduced in later-generation hybrids and backcrosses.
Morphometric analyses based on interlandmark distances (ilDs)
The goal of this analysis was to characterize body shape differences among parental species and the putative hybrids to test the predictive value of this technique to identify the genetic groupings (excluding O. platensis). Almost all individuals of O. bonariensis, O. argentinensis, and putative hybrids used in the molecular analyses were used here. Additional individuals were taken from (González-Castro et al. 2016) (see Table S1 to compare the new individuals analyzed here and in that study). Twenty-six morphometric variables were measured as ilDs on a truss network (Bookstein et al. 1985) based on 12 anatomically defined landmarks (González-Castro et al. 2016). All measurements were taken on the left side of specimens, using a digital caliper (0.05-mm precision). Data analysis followed approaches described by González-Castro et al. (2016). Briefly, a normalization technique to scale the data that exhibits an allometric growth was used according to Lleonart et al. (2000). After transformation, a new matrix was constructed containing the corrected matrices for each group and a PCA was performed using MULTIVARIADO software (Salomon et al. 2004). Principal components scores (PCs) were subsequently used for Discriminant Analysis (DA) using SPSS v.13.0, in order to build a predictive model of group membership based on the original measurements. This procedure generates a set of discriminant functions based on linear combinations of the predictor variables that provides the best discrimination among groups. The functions are generated from a sample of cases for which group membership is known; the functions can then be applied to new cases with measurements for the predictor variables but unknown group membership.
Distribution and abundance of taxonomic groups in relation to water salinity
For this analysis, we used archived data of sampling surveys performed since 2005 by the first author in the MChL and the adjacent coast (see Table S3). The data include more than 60 sampling points over 14 years of work in the MChL region. Water salinity (ppt) was measured in the field after collecting fish samples using a Hydrobios refractometer. Subsequently, each fish was assigned to one of the five groups (O. bonariensis, MChL resident O. argentinensis, marine O. argentinensis, putative hybrids, and O. platensis). Sampling localities were grouped according to zone (I–III) within the MChL, and samples from the adjacent coast areas were analyzed separately. The objective of this analysis is to estimate the relative abundances among groups inside and outside the lagoon and to estimate the relationship between the abundance of each group and salinity.
Gonadal maturation of putative hybrid individuals
To examine if Odontesthes hybrids are sterile or capable of sexual reproduction, individuals identified as intermediate morphs (putative hybrids) were sexed and scored on a macroscopic maturity scale to assess the development of the gonads. Five categories were used following (Brown-Peterson et al. 2011): 1- immature, 2- developing, 3- spawning capable (which include a sub-phase of active spawning), 4- regressing, and 5- regenerating.
Results
Table S2 shows the different data sets used for the following analyses, showing number of individuals, number of groups or species, SNPs, number of loci, missing data, and software used for analyses.
Phylogenomics and population structure analyses
The phylogeny among individuals (Fig. 2) based on 9876 loci shows that all specimens (n = 110) assigned to each of the species included in this study form reciprocally monophyletic groups, in agreement with previous results (Hughes et al. 2020). Odonthestes platensis, not included in that study, is shown here to be distantly related to the other two species present in MChL. All relationships among different species obtained very high support values. In contrast, intermediate morphotypes (putative hybrids) from MChL do not form a monophyletic group and are placed along the branch between O. bonariensis and O. argentinensis, suggesting that they are likely hybrids with varying genetic affinities with these two species.

Odontesthes species in bold were collected for this study inside and outside the Mar Chiquita Coastal Lagoon.
Population genetic structure assessed with FastStructure and ALStructure for all specimens of O. bonariensis, O. argentinensis, and the intermediate morphs (Dataset 2, n = 82, 2936 loci) revealed that the optimal number of populations (K) was two (Fig. 3a, Table S2). In agreement with the phylogenetic analysis, all individuals from O. bonariensis and O. argentinensis sorted into their respective groups, while intermediate morphotypes were found to be hybrids between the two (see hybridization section). Analyses of Dataset 3 (all O. argentinensis individuals and intermediates morphs, n = 55, 9822 loci) also resulted in K = 2 groups, with hybrids clearly separating into their own group (Fig. 3c). Analysis of “pure” O. argentinensis individuals alone (Dataset 4, n = 41, 9390 loci) also resulted in a preferred K = 2 (Fig. 3d), revealing different genetic identities for individuals of marine populations and those that are MChL residents. Finally, results using Dataset 5 with O. bonariensis specimens from inside MChL and from other localities (n = 28, 12456 loci) indicated lack of genetic distinction among populations in this species (Fig. 3b). Fig. S1 shows K plots for all these analyses.

a Results based on O. bonariensis, hybrids and O. argentinensis. b Results based on different localities of O. bonariensis. c Results based on hybrids and O. argentinensis. d Results based on different localities of O. argentinensis (inside and outside the MChL).
Hybridization
The 14 individuals with intermediate phenotype were assigned to a hybrid class by gghybrids with HI ranging from 0.49 to 0.51 (Fig. 4a), while individuals of O. argentinensis and O. bonariensis from MChL were assigned HIs ranging from 0.01 to more than 0.87, respectively. This analysis was calibrated with pure parental species represented by individuals of both species collected from populations outside MChL. Compared to these reference populations (with HI = 0 or HI = 1), HIs of MChL-resident O. bonariensis individuals were slightly shifted toward the value of hybrids (0.5 < 0.87 < 1) but this shift was less marked for MChL-resident O. argentinensis (Fig. 4a). NewHybrid and Hybriddetective identified seven individuals with intermediate morphology as F1 hybrids and another seven individuals as F2 generation hybrids (posterior probabilities > 0.98) (Fig. 4c), but the probability of back-crossed individuals with parental types was negligible. All the O. argentinensis and O. bonariensis from MChL not assigned as parental types a priori were detected as pure species by this analysis. All 14 hybrids had lower than expected inter-taxon heterozygosities (~0.2) at ancestry-informative SNPs given their respective hybrid indices (0.4–0.6), with no obvious disparities among F1 and F2 individuals (Fig. 4b).

a Hybrid index estimated for each individual in gghybrids, individuals are ordered along the x-axis. b A triangle plot of hybrid index and observed heterozygosity calculated for all putative hybrids. Colors correspond to the Hybriddetective assignment class. c Plot produced by Hybriddetective. Individuals are ordered along the x-axis while the y-axis shows the individual-specific cumulative posterior probability of assignment to each of the six possible hybrid classes.
Morphometric analysis based on ilDs
The 26 normalized interlandmark distances (ilDs), which were analyzed by PCA of the correlation matrix, produced six eigenvalues greater than one. The first four PCs explained more than 71% of the variance in the data. Only correlations (between variables and components) higher than 0.50 were taken as significant. PCA based on ilDs allowed almost a total graphic segregation of the four groups analyzed, excluding O. platensis (Fig. 5). Intermediate morphs were completely located in the second quadrant, with higher loadings for the 1-3, 1-4, 2-3, 2-4, 3-4 ilDs that represent the length and height of the head (i.e, bigger head); also variables 5-6, 6-7, 7-8, and 8-9 had high loadings that denote pronounced robustness of the body at Box-Trusses III and IV (Fig. 5a). In addition, they showed lower values for the 4-6 variable, which represents the distance between the end of the head and the origin of the first dorsal fin, thus indicating that this fin originates nearer to the head for intermediate morphs when compared with the other Odontesthes groups analyzed. The specimens of O. bonariensis were basically located in the third and, to a lesser extent, second quadrants. This species was characterized by a higher loading for the ilD 5-7, which indicates their individuals possess a larger distance between the origins of pelvic and anal fins, when compared to the other morpho-groups (Fig. 5). O. bonariensis specimens displayed higher loadings for the ilDs 1-3, 1-4, 2-4 (i.e, a longer head) and, to a lesser degree, 2-3, 3-4 (head height), and 5-6 (body height) (Fig. 5). Also, they showed lower values for the 9-10, 9-11, 9-12, 10-11, 10-12, 12-11, 7-8, and 8-9 variables, all ilDs of the fourth and fifth Box-Trusses that denote a small caudal peduncle. In contrast, the MChL-resident O. argentinensis located in the first quadrant and was characterized by the highest loadings for the 7-8, 8-9 9-10, 9-11, 9-12, 10–11, 10-12, and 12-11 IlDs which denote a robust Box-Truss IV and robust and elongated caudal peduncle (Box-Truss V) (Fig. 5). Also, these specimens showed lower loadings for the IlDs 4-6 and 5-7. Finally, the marine O. argentinensis was located in the fourth quadrant and showed higher loadings for the 4–6 IlD, but also 9-10, 9-11, 9-12, 10–11, 10-12, and 12-11 variables, which denote a long caudal peduncle. They also showed lower loadings for 1-4, 2-3, 2-4, 3-4 (indicating a small size head) and 6-7, 7-8, and 8-9 IlDs (Fig. 5). The data corresponding to 26 PCs of the PCA were employed to perform the DA for the 95 individuals of Odontesthes and produced three significant canonical discrimination functions. The first two discriminant functions explained 92.7% of the total variance in the data, (Wilks’-lambda test of functions 1–3 = 0.001, P < 0.0001). The DA correctly classified 100% of the Odontesthes individuals according to the four groups defined a priori, whereas the cross-validated analysis correctly classified 97.9% of the fish according to their body shape (Table 1). Accordingly, group misclassifications were only recorded in intermediates silversides, where 14.3% of its individuals were misclassified as O. argentinensis from MChL (Table 1). Four groups were defined, according to those defined a priori in the PCA, and their centroids and individuals were separated both on the first and second discriminant functions (Fig. S1).

a Box truss and interlandmark distances based on 12 anatomical landmarks in a hybrid individual. b Projection of individuals’ scores onto the first factorial plane of PCA. c the correlation of the variables labeled as a.
Relationship between species-group abundance and salinity
The relative abundance of each group differs considerably between zones within MChL (Table 1, Figs. 6, 7, and Table S3). Sampling surveys over 14 years show that the most abundant species by far is O. argentinensis (>90%). The resident MChL morph of this species is predominant (85.5%) compared to purely marine forms (5%) which enter the lagoon frequently but have been recorded mostly in Zone I where salinity is highest (average 25.8 ppt). Odontesthes bonariensis, in contrast, make up only 1.8% of all individuals collected in MChL and they can be found in Zone III, but mostly in tributaries to MChL where salinity is lowest (between 1 and 7 ppt). Putative hybrids represented 7.5% of all samples and were collected from all zones of MChL, including freshwater tributaries where they reach their highest frequency (up to 20%). Salinity preference for hybrids seems to be intermediate (around 9 ppt) and similar to the resident MChL O. argentinensis, but both groups were recorded from waters with a wide range of salinities. Finally, O. plantensis was sampled only once inside the MChL in Zone I when the salinity was very high (34 ppt).

a abundance of freshwater input, b intermediate conditions and typical distribution of species, c abundance of marine water input, d pictures of O. bonariensis, hybrids, and O. argentinensis from inside and outside the Mar Chiquita Coastal Lagoon.

Abundance of silverside specimens (y-axis) collected in Mar Chiquita Lagoon between 2005 and 2018 in the different zones shown in Fig. 6a in relation to water salinity at sampling locations.
Gonadal maturation of hybrid individuals
Of the 19 hybrid individuals analyzed, four were classified as immature. The remaining 15 specimens were adults, which exhibited different macroscopic gonadal maturation stages showing some degree of gonadal development, in concordance with the annual life cycle described for O. argentinensis of MChL by (González-Castro et al. 2009). Sex ratio was heavily female-biased since a majority of mature specimens were identified as female (86% of the total sample; n = 13/15). Four of the five maturation phases were found (Table 2). This evidence rejects the hypothesis that hybrid individuals are sterile, given the presence of multiple sexual maturity phases and that almost 60% of the individuals analyzed were females in the spawning capable phase, carrying vitellogenic oocytes in their ovaries (Table 2).
Discussion
Hybridization between marine and freshwater silversides
Marine Odontesthes argentinensis and freshwater O. bonariensis have partially overlapping ranges in countless brackish environments along the Atlantic coast of South America. Hypotheses of hybridization among these species had been proposed based on the observation of intermediate forms (González-Castro et al. 2016) and the lack of differentiation in their mitochondrial genomes (Hughes et al. 2020). This study provides unambiguous genetic evidence from thousands of nuclear loci to characterize hybrid individuals between these two species in a coastal lagoon of Buenos Aires Province in Argentina (the MChL). Our findings may represent a common condition along the vast geographic space where these two species come into secondary contact, and more generally for other species of Odontesthes that display similar patterns. In taxa where hybridization is widespread, detailed knowledge of the dynamics of hybrid zones is fundamental for understanding their evolutionary history and the stability of species boundaries. Our results also confirm that O. platensis is a more distantly related species and that, although it is present in the MChL as an occasional visitor, it does not hybridize with other silverside species (Fig. 2c).
The frequency of hybrids detected in MChL is relatively low (7.5%) when compared to the abundance of other Odontesthes species in the same habitat (González-Castro et al. 2009, 2016) (Table 1). Environmental conditions in this coastal lagoon can vary greatly in a short period of time, modifying the ecological factors that modulate the hybrid zone and the distribution of marine and freshwater species. Odontesthes bonariensis is restricted to fresh water or slightly brackish water habitats (salinity 1–2 ppt) in Zone III and tributaries. Variation in population density (Berasain et al. 2015), embryo survival rate (Noguez Piedras et al. 2009; Colautti et al. 2020), and sperm motility (Chalde et al. 2016) have been shown to be affected by salinity for this species. In contrast, MChL-resident O. argentinesis are abundant throughout the lagoon and show tolerance to highly variable salinity conditions (from 1–34 ppt, but seem to prefer salinities ~10 ppt, Fig. 6, Table 1). As a consequence, the narrow contact zone seems to be limited to low salinity conditions in Zone III and tributaries, limiting opportunities for hybridization and, as a consequence, the number of hybrid individuals available for study. Genetic analyses of intermediate morphs reveal that they are F1 or F2 hybrids, with no evidence of backcrossing to either parental species which, in turn, show no signs of introgression. These results are consistent with the observation of advanced gonadal maturation stages seen in most hybrid individuals (~60%), indicating that they are able to reproduce. Hybrids are found throughout MChL and seem to tolerate a wide range of salinities, but their low frequency overall suggests that hybrid fitness within MChL must be considerably lower than fitness of the most abundant resident species, O. argentinensis. The same is true for freshwater tributaries where O. bonariensis is the most abundant species. Given this disparity in abundance among hybrids and parental species, it is surprising to find only weak evidence of backcrossing to O. bonariensis, as indicated by gghybrids. The triangle plot displaying observed heterozygosity and hybrid index in hybrid individuals (Fig. 4b) is consistent with no back-crosses, without hybrids in off-diagonal positions. But the empirical triangle plot also revealed that, against expectations, F1 and F2 hybrids have very low observed heterozygosity (<0.4), possibly due to an artifact of the method. It has been shown that RADseq data processing may result in under calling heterozygotes due to polymorphism in restriction sites at one allele, or due to low or uneven counts of reads at heterozygous sites (Huang et al. 2009; Fragoso et al. 2016; Zheng et al. 2018). Larger sample sizes, deeper sequencing efforts, or application of error correction tools may be necessary to increase the power of these tests to define hybrid classes, especially backcrossed individuals, with confidence.
In addition to environmental constraints, intrinsic processes such as lower fecundity or lower larval survival, or other maladaptive traits putatively present in hybrids due to genetic incompatibilities may be considered to explain the observed pattern. Although small, the sample of hybrid individuals shows a highly biased sex ratio (86% female, Table 2), suggesting that known differences in sex determination mechanisms among Odontesthes species (Strüssmann et al. 1997; Yamamoto et al. 2014; Hattori et al. 2019) also may affect overall hybrid fitness. Both parental species are known to carry a master sex-determining gene (Strüssmann et al. 1997; Yamamoto et al. 2014; Hattori et al. 2019) that is the dominant factor establishing genetic sex determination (GSD) in O. argentinenis. In contrast, O. bonariensis exhibits strong temperature-dependent sex determination (TSD). Developing larvae of this species undergo a thermo-sensitive period in the first weeks after hatching during which the water temperature will determine the phenotypic sex, frequently overriding the genotypic sex. Hatchlings reared at water temperatures of 17 °C or 29 °C during post-hatching weeks 1–5 develop population sex ratios of 100% females or males, respectively (Strüssmann et al. 1997). The reproductive physiology of O. bonariensis, including TSD, has been extensively studied but remains far from being fully understood (Miranda et al. 2013; Yamamoto et al. 2014; Fernandino et al. 2015; Hattori et al. 2019). Finely tuned interaction between GSD and TSD in parental species may be disrupted in hybrids, leading to the observed biased sex ratio and implied negative fitness consequences. If the sex ratio observed in our sample is representative of the hybrid population as a whole, it is more likely that backcrossing will occur with either parental species (the most abundant at a particular locality) than reproduction among hybrids.
The two closely-related parental species are well differentiated morphologically (Fig. 5) and all individuals analyzed in this study were unambiguously assigned to either species based on analyses of genetic data (Figs. 2 and 3a). The reciprocal monophyly of O. bonariensis and O. argentinensis individuals in the MChL with other members of their species from more distant localities (Fig. 2) suggests that the rate of hybridization between these species is not sufficient to disrupt their genomic identity. This is confirmed by previous analyses of SNPs obtained from transcriptomes (Hughes et al. 2017). Analysis of gene expression in gills of these two species also revealed consistent differences in expression and coding sequences that could be predicted on the basis of gill osmoregulatory function. That study (Hughes et al. 2017) shows that, among many others, the gene coding for the Na/K transporting ATPase subunit alpha 1—one of the best known osmoregulatory genes in fish that has been linked to marine-to-freshwater transitions in the stickleback system (Jones et al. 2012)—is both differentially expressed and also contains a nonsynonymous SNP between O. argentinensis and O. bonariensis. Other genes showing differential expression and non-synonymous SNPs between the parental species included heat-shock proteins and the claudin family of tight-junction proteins (Hughes et al. 2017). The latter gene family has been suggested to be critical for species adapted to variable salinities such as the European sea bass (Tine et al. 2014). Furthermore, that study also reports differential expression of other genes associated with the immune response, such as those coding for MHC molecules (major histocompatibility complex) and lectins that have been linked to habitat diversity and speciation in fishes (Malmstrøm et al. 2016) or shown to be differentially expressed between anadromous and resident populations of trout (Boulet et al. 2012), respectively. In other words, genomic differences between parental species reflect adaptive responses to their native habitats that differ not only in salinity but also in their bacterial communities and other environmental factors. Some degree of plasticity, especially in O. argentinensis, provides the opportunity for secondary contact (see below), but both species maintain their distinctive adaptations in the face of hybridization. Hybridization across a strong environmental gradient or ecological transition leads to conditions of “genetic tension” among loci coding for important adaptive traits (Abbott et al. 2013), suggesting that this is an unstable hybrid zone. Opportunities for secondary contact between closely related species that lead to generation of hybrids seem contingent on the pronounced tendency of O. argentinensis to enter and colonize estuaries and coastal lagoons.
Whereas genomic loci under strong selection remain differentiated among parental species, mitochondrial genes have been shown to flow freely between these two species (e.g., García et al. 2014; González-Castro et al. 2016; Hughes et al. 2020). This pattern of mito-nuclear discordance has contributed to historical confusion about the relationships among Odontesthes argentinensis and the freshwater species in its sister clade that includes O. bonariensis and many others in the La Plata basin, throughout southern Brazil and Uruguay—the “O. argentinensis species group,” sensu (Hughes et al. 2020). This freshwater clade, in fact, represents one of the three separate freshwater colonization events inferred for ancestral marine Odontesthes that originated freshwater species in South America. Another instance of mitochondrial introgression from marine-to-freshwater species of Odontesthes (and consequent mito-nuclear discordance) has been documented in some populations in southern Chile of the freshwater species O. mauleanum that carry mitochondrial haplotypes that are common in the marine ancestor O. regia, widespread in south Pacific coastal waters (Hughes et al. 2020). The third freshwater species with marine ancestors is the Patagonian silverside, O. hatcheri, widely distributed in rivers and lakes of Patagonia (east of the Andes). Unlike the other two cases, this species does not form natural hybrids with other silverside species and its mitochondrial and nuclear genomes are well differentiated. However, human-mediated introductions of O. bonariensis into some Patagonian lakes and reservoirs for stocking had drastic effects on the native Patagonian silverside (Rueda et al. 2017). In the most extreme cases, introgressive hybridization triggered by these introductions demographically overwhelmed the native freshwater O. hatcheri, apparently a common outcome for human-mediated introductions. Similarly, the Colorado River Delta endemic silverside Colpichthys hubbsi is threatened by genetic swamping from hybridization with its sister species C. regis, after the salinity in the delta has increased from the effects of damming (Lau and Jacobs 2017). In most cases, anthropogenic actions or alterations of the environment lead to disruptions of genetic barriers and loss of biodiversity (Ottenburghs 2021).
Population structure and plasticity in O. argentinensis
Several studies have documented the pervasive tendency of O. argentinensis to establish local populations in estuaries and lagoons along the Atlantic coastline from northern Argentina to southern Brazil. Many large estuaries and floodplains, such as the La Plata estuary, Mar Chiquita, Mirim, and Dos Patos Lagoons host estuarine ecotypes that can be readily distinguished from their marine counterparts based on life history and reproductive traits. Marine populations lay eggs along the beach in dynamic surf zones (Phonlor and Vinagre 1989), while estuarine silversides prefer submerged vegetation sites for spawning that are abundant in most estuarine environments but almost absent along the marine coast (Moresco and Bemvenuti 2006). Eggs of estuarine ecotypes are slender and have a thin chorion supporting a reduced number of adhesive filaments compared to marine forms (Phonlor and Cousin 1997). An early genetic study of O. argentinensis along the Brazilian coast based on allozymes revealed three loci with significant differences in allelic frequencies in the estuarine population of the Patos Lagoon (Beheregaray and Levy 2000). Subsequent surveys of this system and other coastal lagoons in Brazil based on mtDNA and six microsatellite loci arguably reported genetic differences (population genetic structure) between marine and estuarine populations (Beheregaray and Sunnucks 2001). These authors interpreted the available evidence in light of a model assuming that phenotypic plasticity would pre-adapt estuarine silversides to invade, colonize, and form new species in vacant niches in freshwater habitats (Bamber and Henderson 1988), calling the estuarine populations “incipient species.” More recently, morphometric and meristic data collected for O. argentinensis in MChL in this and previous studies (González-Castro et al. 2016), show that the estuarine resident forms can be distinguished from marine populations (Fig. 5). Another example was found in a landlocked freshwater population near the coast (Salada de Pedro Luro Lake; south-east of Buenos Aires Province, Argentina) where the resident silverside population has unique phenotypic characteristics due to local adaptation, phenotypic plasticity, or both (Colautti et al. 2020).
The new genetic data analyzed in this study suggest some degree of distinction (K = 2, Fig. 3d) among individuals collected inside and outside MChL when these samples were analyzed separately from the rest. The observation that MChL-residents are able to complete their life cycle inside MChL is consistent with this result. A recent analysis of the structure and composition of parasite assemblages of O. argentinensis from inside and outside MChL also showed a high degree of differentiation (Levy et al. 2021). Phenotypic differences can result in local adaptations when, among other necessary conditions, populations live in unstable environments, exchange few or no migrants, and they are subject to differential selective pressures (Ghalambor et al. 2007). Although marine individuals regularly enter MChL, they tend to remain close to the mouth of the lagoon where salinity ranges are always high (Fig. 6). But the genetic cohesion of all O. argentinesis samples, including individuals from distant marine locations (Figs. 2, 3a, and 4b), suggests that high levels of gene flow occur across the vast geographic range of this species in spite of some degree of local differentiation. Among other taxa, the evidence for local adaptations in Atlantic salmon has been well documented (Garcia de Leaniz et al. 2007). Phenotypic plasticity due to local adaptation is a ubiquitous aspect of organisms (Price et al. 2003). In the future, it would be interesting to see how these morphological differences may be correlated with different ecological conditions.
Conclusions
Hybridization appears to be common in the Odontesthes genus, and the hybrid zones are narrow and geographically localized due to the salinity environment. Nonetheless, other hybrid zones in this group could exist in the absence of any obvious physical barriers, suggesting that other ecological or intrinsic factors may be playing a role in maintaining species as distinct entities. The study of hybrid zones advances understanding of the speciation process and approaches incorporating genomic data are increasingly used to draw significant conclusions about the impact of hybridization (Shipham et al. 2019). Our study showed two different levels of differentiation among silverside species in a coastal estuarine habitat that is strongly influenced by salinity. The first between O. bonariensis and O. argentinensis where hybridization persists, but does not appear to blur the species boundaries. The second within populations of O. argentinensis where genetic, morphological and ecological differences were reported.
Data archiving
Raw reads for ddRAD sequences are archived under NCBI BioProject PRJNA796799.
References
Abbott R, Albach D, Ansell S, Arntzen JW, Baird SJE, Bierne N et al. (2013) Hybridization and speciation. J Evol Biol 26:229–246
Anderson EC, Thompson EA (2002) A model-based method for identifying species hybrids using multilocus genetic data. Genetics 160:1217–1229
Bailey RI (2018) gghybrid: evolutionary analysis of hybrids and hybrid zones. R package version 0009000
Bamber RN, Henderson PA (1988) Pre-adaptive plasticity in atherinids and the estuarine seat of teleost evolution. J Fish Biol 33:17–23
Barton NH, Hewitt GM (1985) Analysis of hybrid zones. Annu Rev Ecol Syst 16:113–148
Beheregaray LB, Levy JA (2000) Population genetics of the silverside Odontesthes argentinensis (Teleostei, Atherinopsidae): evidence for speciation in an estuary of Southern Brazil. Copeia 2000:441–447
Beheregaray LB, Sunnucks P (2001) Fine-scale genetic structure, estuarine colonization and incipient speciation in the marine silverside fish Odontesthes argentinensis. Mol Ecol 10:2849–2866
Berasain GE, Colautti DC, Lenicov MR, Argemi F, Bohn VY, Miranda LA (2015) Impact of water salinity on Odontesthes bonariensis (Actinopterygii, Atherinopsidae) fisheries in Chasicó Lake (Argentina). Hydrobiologia 752:167–174
Bookstein FB, Chernoff R, Elder RL, Humpries JM, Smith GR, Strauss RE (1985) Morphometrics in evolutionary biology, 1st edn. Academy of Natural Sciences of Philadelphia, Ed., Philadelphia
Boulet M, Normandeau É, Bougas B, Audet C, Bernatchez L (2012) Comparative transcriptomics of anadromous and resident brook charr Salvelinus fontinalis before their first salt water transition. Curr Zool 58:158–170
Brown-Peterson NJ, Wyanski DM, Saborido-Rey F, Macewicz BJ, Lowerre-Barbieri SK (2011) A standardized terminology for describing reproductive development in fishes. Mar Coast Fish 3:52–70
Cabreros I, Storey JD (2019) A likelihood-free estimator of population structure bridging admixture models and principal components analysis. Genetics 212:1009–1029
Calvelo J, D’Anatro A (2021) Mitochondrial genome architecture and phylogenetic relationships of Odontesthes argentinensis within Atherinomorpha. Genetica 149:129–141
Chalde T, Gárriz Á, Sanches EA, Miranda LA (2016) Influence of pejerreyOdontesthes bonariensis (Valenciennes, 1835) broodstock age on gamete quality, reproductive performance and plasma sex steroid levels during the spawning season. Aquac Res 47:969–982
Colautti DC, Miranda L, Gonzalez-Castro M, Villanova V, Strüssmann CA, Mancini M et al. (2020) Evidence of a landlocked reproducing population of the marine pejerrey Odontesthes argentinensis (Actinopterygii; Atherinopsidae). J Fish Biol 96:202–216
DiVittorio CT, Singhal S, Roddy AB, Zapata F, Ackerly DD, Baldwin BG et al. (2020) Natural selection maintains species despite frequent hybridization in the desert shrub Encelia. Proc Natl Acad Sci USA 117:33373–33383
Dyer BS (2006) Systematic revision of the South American silversides (Teleostei, Atheriniformes). Biocell 30:69–88
Eaton DAR (2014) PyRAD: assembly of de novo RADseq loci for phylogenetic analyses. Bioinformatics 30:1844–1849
Fernandino JI, Hattori RS, Strüssmann CA, Yamamoto Y, Somoza G (2015) Sex determination in fish: Odontesthes spp. (Atherinopsidae) as experimental models. Anim Reprod 12:24–27
Fragoso CA, Heffelfinger C, Zhao H, Dellaporta SL (2016) Imputing genotypes in biallelic populations from low-coverage sequence data. Genetics 202:487–495
García G, Ríos N, Gutiérrez V, Varela JG, Bouza Fernández C, Pardo BG et al. (2014) Promiscuous speciation with gene flow in silverside fish genus Odontesthes (Atheriniformes, Atherinopsidae) from south western Atlantic Ocean basins. PLoS ONE 9:e104659
Garcia de Leaniz C, Fleming IA, Einum S, Verspoor E, Jordan WC, Consuegra S et al. (2007) A critical review of adaptive genetic variation in Atlantic salmon: implications for conservation. Biol Rev Camb Philos Soc 82:173–211
Ghalambor CK, McKAY JK, Carroll SP, Reznick DN (2007) Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct Ecol 21:394–407
Gompert Z, Buerkle CA (2010) introgress: a software package for mapping components of isolation in hybrids. Mol Ecol Resour 10:378–384
González-Castro M, Díaz de Astarloa JM, Cousseau MB, Figueroa DE, Delpiani SM, Bruno DO et al. (2009) Fish composition in a south-western Atlantic temperate coastal lagoon: spatial–temporal variation and relationships with environmental variables. J Mar Biol Ass 89:593–604
González-Castro M, Rosso JJ, Delpiani SM, Mabragaña E, Díaz de Astarloa JM (2019) Inferring boundaries among fish species of the new world silversides (Atherinopsidae; genus Odontesthes): new evidences of incipient speciation between marine and brackish populations of Odontesthes argentinensis. Genetica 147:217–229
González-Castro M, Rosso JJ, Mabragaña E, Díaz de Astarloa JM (2016) Surfing among species, populations and morphotypes: Inferring boundaries between two species of new world silversides (Atherinopsidae). C R Biol 339:10–23
Hattori RS, Somoza GM, Fernandino JI, Colautti DC, Miyoshi K, Gong Z, et al. (2019) The duplicated Y-specific amhy gene is conserved and linked to maleness in silversides of the Genus Odontesthes. Genes 10:679
Hoang DT, Chernomor O, von Haeseler A, Minh BQ, Vinh LS (2018) Ufboot2: improving the ultrafast bootstrap approximation. Mol Biol Evol 35:518–522
Huang X, Feng Q, Qian Q, Zhao Q, Wang L, Wang A et al. (2009) High-throughput genotyping by whole-genome resequencing. Genome Res 19:1068–1076
Hughes LC, Cardoso YP, Sommer JA, Cifuentes R, Cuello M, Somoza GM et al. (2020) Biogeography, habitat transitions and hybridization in a radiation of South American silverside fishes revealed by mitochondrial and genomic RAD data. Mol Ecol 29:738–751
Hughes LC, Somoza GM, Nguyen BN, Bernot JP, González-Castro M, Díaz de Astarloa JM et al. (2017) Transcriptomic differentiation underlying marine-to-freshwater transitions in the South American silversides Odontesthes argentinensis and O. bonariensis (Atheriniformes). Ecol Evol 7:5258–5268
Huson DH, Bryant D (2006) Application of phylogenetic networks in evolutionary studies. Mol Biol Evol 23:254–267
Jones FC, Chan YF, Schmutz J, Grimwood J, Brady SD, Southwick AM et al. (2012) A genome-wide SNP genotyping array reveals patterns of global and repeated species-pair divergence in sticklebacks. Curr Biol 22:83–90
Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS (2017) ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods 14:587–589
Lau CLF, Jacobs DK (2017) Introgression between ecologically distinct species following increased salinity in the Colorado Delta- Worldwide implications for impacted estuary diversity. PeerJ 5:e4056
Levy E, Canel D, Rossin MA, González-Castro M, Timi JT (2021) Parasite assemblages as indicators of an incipient speciation process of Odontesthes argentinensis in an estuarine environment. Estuar Coast Shelf Sci 250:107168
Lleonart J, Salat J, Torres GJ (2000) Removing allometric effects of body size in morphological analysis. J Theor Biol 205:85–93
Malmstrøm M, Matschiner M, Tørresen OK, Star B, Snipen LG, Hansen TF et al. (2016) Evolution of the immune system influences speciation rates in teleost fishes. Nat Genet 48:1204–1210
Miranda LA, Chalde T, Elisio M, Strüssmann CA (2013) Effects of global warming on fish reproductive endocrine axis, with special emphasis in pejerrey Odontesthes bonariensis. Gen Comp Endocrinol 192:45–54
Montanari SR, Hobbs J-PA, Pratchett MS, van Herwerden L (2016) The importance of ecological and behavioural data in studies of hybridisation among marine fishes. Rev Fish Biol Fish 26:181–198
Moore WS, Buchanan DB (1985) Stability of the northern flicker hybrid zone in historical times: implications for adaptive speciation theory. Evolution 39:135
Moresco A, Bemvenuti MDA (2006) Biologia reprodutiva do peixe-rei Odontesthes argentinensis (Valenciennes)(Atherinopsidae) da região marinha costeira do sul do Brasil. Rev Bras Zool 23:1168–1174
Nguyen L-T, Schmidt HA, von Haeseler A, Minh BQ (2015) IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol 32:268–274
Noguez Piedras S, Osório Fernandes J, Motoyama I, Martins G (2009) Efeito de diferentes concentracoes de salinas (NaCl) na sobrevivencia de embrioes de peixe—rei Odontesthes bonariensis e Odontesthes humensis. Biotemas 22:235–238
Ottenburghs J (2021) The genic view of hybridization in the anthropocene. Evol Appl 14:2342–2360
Peterson BK, Weber JN, Kay EH, Fisher HS, Hoekstra HE (2012) Double digest RADseq: an inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS One 7:e37135
Phonlor G, Cousin JC (1997) Early life history of silverside fishes. In: Seeliger U, Odebrecht C, Castello JP (eds) Subtropical convergence environments: the coast and sea in Southwestern Atlantic. Springer-Verlag, Berlin, p 136–141
Phonlor G, Vinagre LEC (1989) Efeito do retarde da primeira alimentação sobre o crescimento ea sobrevivência da larva de Odonthestes argentinensis. Atlântica 11:63–75
Pina-Martins F, Silva DN, Fino J, Paulo OS (2017) Structure_threader: an improved method for automation and parallelization of programs structure, fastStructure and MavericK on multicore CPU systems. Mol Ecol Resour 17:e268–e274
Price TD, Qvarnström A, Irwin DE (2003) The role of phenotypic plasticity in driving genetic evolution. Proc Biol Sci 270:1433–1440
de Queiroz K (2007) Species concepts and species delimitation. Syst Biol 56:879–886
Raj A, Stephens M, Pritchard JK (2014) fastSTRUCTURE: variational inference of population structure in large SNP data sets. Genetics 197:573–589
Rhymer JM, Simberloff D (1996) Extinction by hybridization and introgression. Annu Rev Ecol Syst 27:83–109
Rueda EC, Mullaney KA, Conte-Grand C, Habit EM, Cussac V, Ortí G (2017) Displacement of native Patagonian freshwater silverside populations (Odontesthes hatcheri, Atherinopsidae) by introgressive hybridization with introduced O. bonariensis. Biol Invasions 19:971–988
Salomon R, Melo S, Winzer N (2004) MULTIVARIADO software. Departamento de Matemática
Sheaves M, Baker R, Nagelkerken I, Connolly RM (2015) True value of estuarine and coastal nurseries for fish: incorporating complexity and dynamics. Estuaries Coast 38:401–414
Shipham A, Joseph L, Schmidt DJ, Drew A, Mason I, Hughes JM (2019) Dissection by genomic and plumage variation of a geographically complex hybrid zone between two Australian non-sister parrot species, Platycercus adscitus and Platycercus eximius. Heredity 122:402–416
Somoza GM, Miranda LA, Berasain GE, Colautti D, Remes Lenicov M, Strüssmann CA (2008) Historical aspects, current status and prospects of pejerrey aquaculture in South America. Aquac Res 39:784–793
Strüssmann CA, Saito T, Usui M, Yamada H, Takashima F (1997) Thermal thresholds and critical period of thermolabile sex determination in two atherinid fishes,Odontesthes bonariensis andPatagonina hatcheri. J Exp Zool 278:167–177
Taylor EB, Boughman JW, Groenenboom M, Sniatynski M, Schluter D, Gow JL (2006) Speciation in reverse: morphological and genetic evidence of the collapse of a three-spined stickleback (Gasterosteus aculeatus) species pair. Mol Ecol 15:343–355
Taylor SA, Larson EL (2019) Insights from genomes into the evolutionary importance and prevalence of hybridization in nature. Nat Ecol Evol 3:170–177
Tine M, Kuhl H, Gagnaire P-A, Louro B, Desmarais E, Martins RST et al. (2014) European sea bass genome and its variation provide insights into adaptation to euryhalinity and speciation. Nat Commun 5:5770
Todesco M, Pascual MA, Owens GL, Ostevik KL, Moyers BT, Hübner S et al. (2016) Hybridization and extinction. Evol Appl 9:892–908
Wringe BF, Stanley RRE, Jeffery NW, Anderson EC, Bradbury IR (2017) hybriddetective: a workflow and package to facilitate the detection of hybridization using genomic data in R. Mol Ecol Resour 17:e275–e284
Yamamoto Y, Zhang Y, Sarida M, Hattori RS, Strüssmann CA (2014) Coexistence of genotypic and temperature-dependent sex determination in pejerrey Odontesthes bonariensis. PLoS One 9:e102574
Zhang L, Thibert-Plante X, Ripa J, Svanbäck R, Brännström Å (2019) Biodiversity loss through speciation collapse: mechanisms, warning signals, and possible rescue. Evolution 73:1504–1516
Zheng C, Boer MP, van Eeuwijk FA (2018) Accurate genotype imputation in multiparental populations from low-coverage sequence. Genetics 210:71–82
Acknowledgements
The authors would like to thank Julio Mangiarotti (Mar Chiquita Biosphere Reserve); Pablo Rizzo, Cristian Di Paolo, Nadir Dupuy, and Marcelo Pons (sport fishing guides of Mar Chiquita Coastal Lagoon); Daniel Blanco, Santiago Gaudioso, Juan Pablo Gaudioso (San Gabriel and Juan y Juan fishing-points in Mar Chiquita Coastal Lagoon) and Daniel Giménez (silversides sport fishing expert) and Mar Chiquita Town Hall authorities. Rose Peterson (George Washington University) assisted with DNA extractions. Three anonymous reviewers provided insightful comments to improve the manuscript. During this project, YPC was supported by the Fulbright-CONICET fellowships to visit the Department of Biological Sciences at George Washington University. This work was funded in part by the PICT2017-1330 and PICT2019-1344 (YPC/MGC); PIP0339 (MGC) and a National Science Foundation grant DEB1457426 (GO).
Author information
Authors and Affiliations
Contributions
MGC and YPC performed fieldwork to obtain samples, participated in DNA data collection, analyses, and prepared the first draft of the manuscript. LCH participated in data analysis and GO edited the manuscript and provided financial support. All authors designed the research.
Corresponding author
Ethics declarations
Competing interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Associate editor: Ben Evans.
Supplementary information
Rights and permissions
About this article

Cite this article
González-Castro, M., Cardoso, Y.P., Hughes, L.C. et al. Hybridization is strongly constrained by salinity during secondary contact between silverside fishes (Odontesthes, Atheriniformes). Heredity 129, 233–243 (2022). https://doi.org/10.1038/s41437-022-00555-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41437-022-00555-9
