
Abstract
Neurofibromatosis type 1 (NF1) is a hereditary tumour syndrome that predisposes to benign and malignant tumours originating from neural crest cells. Biallelic inactivation of the tumour-suppressor gene NF1 in glial cells in the skin, along a nerve plexus or in the brain results in the development of benign tumours: cutaneous neurofibroma, plexiform neurofibroma and glioma, respectively. Despite more than 40 years of research, only one medication was recently approved for treatment of plexiform neurofibroma and no drugs have been specifically approved for the management of other tumours. Work carried out over the past several years indicates that inhibiting different cellular signalling pathways (such as Hippo, Janus kinase/signal transducer and activator of transcription, mitogen-activated protein kinase and those mediated by sex hormones) in tumour cells or targeting cells in the microenvironment (nerve cells, macrophages, mast cells and T cells) might benefit NF1 patients. In this review, we outline previous strategies aimed at targeting these signalling pathways or cells in the microenvironment, agents that are currently in clinical trials, and the latest advances in basic research that could culminate in the development of novel therapeutics for patients with NF1.
Similar content being viewed by others
Background
Neurofibromatosis type 1 (NF1), first characterised in detail by the German pathologist Friedrich von Recklinghausen in 1882, is an autosomal-dominant genetic disorder that results from biallelic inactivation of the tumour-suppressor gene NF1. The NF1 gene encodes neurofibromin, a GTPase-activating protein (GAP) that negatively regulates the oncoprotein RAS. About half of all cases of the disorder involve de novo NF1 mutations, which can be passed on to successive generations. The penetrance is almost 100%, but the expressivity varies greatly, even between twins. Indeed, even though 99% of NF1 patients meet the diagnostic criteria (Box 1)1 by the age of 20 years,2 the manifestations are unpredictable even within the same family harbouring an identical mutation. This observation has given rise to the spatiotemporal hypothesis, which proposes that, in addition to the nature of the NF1 mutation and additional genetic/epigenetic differences, the timing and the exact cell population that is undergoing loss of the second functional copy of NF1 are key factors that explain the overall phenotype.3
Clinically, NF1 affects 1 in every 3000 new-borns regardless of gender or race. This multisystem disorder has diverse manifestations including neurocognitive, skeletal and cardiovascular abnormalities, and is also classified as a neurocutaneous syndrome to highlight its manifestations primarily in the nervous system and the skin as a result of lesions affecting neural-crest-derived tissues such as Schwann cells and melanocytes.4
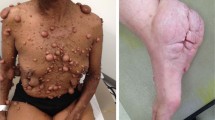
NF1 patients are predisposed to develop multiple types of benign and malignant tumour, including neurofibromas, malignant peripheral nerve sheath tumours (MPNSTs) and optic pathway gliomas (OPGs). Gastrointestinal stromal tumours, breast cancer, juvenile myelomonocytic leukaemia, rhabdomyosarcoma and pheochromocytoma5 are also associated with NF1; however, these neoplasms occur at a much lower frequency and will not be discussed in this review. Neurofibromas are benign tumours of the nerve sheath that are composed of neoplastic Schwann cells and non-neoplastic fibroblasts, mast cells, macrophages, endothelial cells, nerves and other cell types as well as abundant collagen deposition. Neurofibromas develop in the peripheral nervous system, and can be generally grouped into two main subtypes—cutaneous neurofibromas (cNFs) and plexiform neurofibromas (pNFs)—by their locations. cNFs, which occur in nearly all NF1 patients and are one of the defining features of the disorder, are discrete masses associated with cutaneous nerves, the development of which coincides with puberty.6 However, the clinical manifestations in cNF can be highly variable depending on individuals; some patients develop only a few visible tumours, whereas the entire body is covered in other cases. By contrast, pNFs arise from multiple nerve fascicles internally and infiltrate the surrounding soft tissue. Most pNFs are believed to be congenital and are more likely to grow during childhood. In the cases of cNF and pNF, no additional pathogenic mutation is recurrently found in neurofibroma except the one located in NF1.7,8 Clinical management largely comprises surgical resection, and no effective pharmacological agent is yet available for effective treatment or cure, although some agents have shown promising results in preclinical or clinical trials, and will be discussed in detail in this review.
A wide range of animal models (Box 2) has been developed to study the intracellular signalling pathways as well as extracellular signals that regulate tumour development in NF1. Tumorigenesis is a multi-step process that progressively transforms a normal tissue into a benign one (e.g. cNF, pNF, OPG) and potentially subsequently into a malignant one (e.g. MPNST). In the context of NF1-related tumours, it is unclear whether all NF1-associated MPNSTs originate from a pre-existing pNF. However, a pNF can progress into an atypical neurofibromatous neoplasm of uncertain biological potential (ANNUBP)9 typically as a result of a subsequent genetic mutation in CDKN2A10,11 and, upon further genetic insult to additional tumour-suppressor genes, such as TP53, PTEN or members of the PRC2 chromatin-remodelling complex,12 an ANNUBP or a pNF can transform into a MPNST. The lifetime risk of developing MPNSTs is 8–13% in all patients with NF1,13 although this risk is higher in patients with pNFs. MPNSTs are life-threatening, highly morbid, and show a high propensity for metastasising. Surgical resection of MPNSTs is challenging because these tumours are commonly inaccessible within the body or they infiltrate vital nerves—hence the urgent need for effective therapeutic treatment.
NF1 patients can also develop OPGs. These low-grade brain tumours affect young children, with most being diagnosed before the age of 5 years old. Although OPGs rarely cause death, they can lead to vision loss and even blindness in many patients.14 However, the mechanism of vision loss in OPGs is still unclear. First-line chemotherapy, using vincristine and carboplatin, for example, is usually effective at stabilising the disease15 but it is still challenging to predict who will benefit from this treatment as well as to decide who actually needs treatment. The microenvironment of OPGs is very heterogeneous and plays an essential role in tumorigenesis as in neurofibromas. OPGs are composed of astrocytes, oligodendrocytes, neurons and microglia (the nervous system equivalent of macrophages).16 Several studies have demonstrated that OPG tumour cell proliferation is largely dependent on the microglia,17,18,19 which secrete chemokine ligands to recruit more microglia and to induce the proliferation of astroglial tumour cells.20 In addition, microglia can cause neurotoxicity (e.g. by releasing reactive oxygen species), which can damage optic nerve axons.21
The focus of this review is to outline therapeutic strategies that have previously been developed and tested, agents that are currently being tested in clinical trials and future therapies that extrapolate from the latest published basic research in the laboratory for the treatment of the most common NF1-related tumours, including cNFs, pNFs, MPNSTs and OPGs.
Therapeutic strategies developed and tested in the past
Significant progress has been made by the neurofibromatosis research community, including the genetic cause of NF1, the cellular function of the neurofibromin protein, and the identification of cellular components that contribute to the tumours developed in NF1. This knowledge has been translated into clinical trials of putative therapeutic targets in an attempt to develop new treatments for NF1 (Fig. 1).

Overview of previously trialled therapies for the most common types of tumour associated with NF1. Therapeutic agents include the farnesyltransferase inhibitor ipifarnib, inhibitors of receptor tyrosine kinases (RTKs) upstream of RAS (imatinib, sunitinib, sorafenib), inhibitors of components of the pathway downstream of RAS (mTOR inhibitors sirolimus, everolimus) and anti-fibrotic agents such as pirfenidone.
Upstream of Ras: receptor tyrosine kinase inhibitors
The best characterised biochemical function of NF1 is the inhibition of Ras signalling mediated by its GAP domain. Ras (KRAS, NRAS and HRAS) is the most frequently mutated oncogene in human cancer22 but is notoriously ‘undruggable’.23 Indeed, directly targeting Ras using, among other agents, the farnesyltransferase inhibitor tipifarnib has been extremely challenging in general24 and was also demonstrated to be ineffective in the treatment of pNF25 (NCT00021541). As Ras itself is not mutated in NF1, an alternative strategy has been to target the upstream activators of Ras—receptor tyrosine kinases (RTKs).
Epidermal growth factor receptor (EGFR) is an RTK that is abundantly expressed in neurofibroma and MPNST cell lines, despite not normally being expressed in Schwann cells.26 Indeed, expressing EGFR in Schwann cells in transgenic mice leads to the formation of neurofibroma-like lesions (although at a very low penetrance), whereas an inactivating EGFR mutation decreases its tumorigenic potential in a mouse model of MPNST.27 However, in humans, the EGFR inhibitor erlotinib largely failed to inhibit tumour growth in a Phase 2 trial of patients with advanced stage MPNST28 (NCT00068367). It remains to be determined if erlotinib or another potent EGFR inhibitor would be clinically effective in the context of pNF, cNF or OPG, and whether or not patient stratification based on high level of EGFR positivity would be a better approach.
Platelet-derived growth factor receptor (PDGFR) is another RTK that is overexpressed in Schwann cells derived from neurofibromas29 and MPNSTs,30 and the expression levels of the c-met RTK proto-oncogene product and its ligand hepatocyte growth factor (HGF) correlate with disease progression.31 However, despite promising results from preclinical studies, including the targeting of mast cells through inhibition of the c-Kit RTK,32 when inhibitors targeting various relevant RTKs were tested in clinical trials using sorafenib for patients with OPG33 (NCT01338857) or imatinib for pNF34 (NCT01673009) and MPNST35 (NCT00031915), disappointingly, they largely lacked efficacy at reducing tumour burden.
Neurofibroma is highly vascularised,36 and the vascular endothelial growth factor (VEGF) ligand is known to be expressed in cNF37 and at even higher levels in MPNST, where it correlates with poor patient prognosis.38 A small molecule inhibitor targeting the RTK VEGF receptor 2 (VEGFR2) decreased angiogenesis and proliferation by over 50% in a tumour explant xenograft model39 but, unfortunately, in clinical trials, targeting VEGF-A using the monoclonal antibody ranibizumab was not effective for cNF (NCT00657202), and targeting VEGFR through the RTK inhibitor sorafenib was not effective for MPNST40 (NCT00245102). However, as sorafenib has multiple targets, it is not appropriate to simply imply that failure to inhibit VEGFR is the main reason for this result. On a positive note, cabozantinib, a small molecule inhibitor of the RTKs VEGFR2, c-MET, AXL and RET, was reported at the 2018 Joint Global Neurofibromatosis Conference to have some efficacy against pNF (NCT02101736). Furthermore, when treated with bevacizumab, a monoclonal antibody targeting VEGF-A, some children with OPG have been reported to respond positively, as indirectly judged by their improved vision.41
Downstream of Ras: mammalian target of rapamycin inhibitor
The mammalian target of rapamycin (mTOR) pathway is a central survival pathway downstream of Ras. Inhibition of mTOR is effective in the context of tuberous sclerosis, another neurocutaneous disease, prompting investigation of pharmacological inhibition of the mTOR pathway for NF1. However, although this approach decreased tumour cell proliferation and, consequently, tumour volume in an NF1 mouse model,42,43 mTOR inhibitors were ineffective against pNF in one clinical trial44 but prolonged progression-free survival, albeit modestly, in another trial45 (NCT00634270). A combination trial of mTOR inhibition plus bevacizumab was also ineffective for MPNST46 (NCT01661283). The mTOR inhibitor everolimus did, however, show promising early results in a pilot trial for cNF47 (NCT02332902) and is currently being tested in a clinical trial in patients with OPG (NCT01158651).
Anti-fibrotic agents
Von Recklinghausen coined the term ‘neurofibromatosis’ to highlight the abundance of nerve cells and fibrosis in neurofibromas. A hallmark of these tumours is the presence of collagen, which can comprise up to 50% of the dry weight of a tumour.48 As fibroblasts are collagen-producing cells and, by definition, fibrosis is an excess of collagen deposition, it has been hypothesised that neurofibroma constitutes ‘nerve fibrosis’. In support of this model, transforming growth factor β (TGF-β), one of the key signalling molecules involved in fibrosis, is secreted by mast cells upon stimulation by neoplastic Nf1–/– Schwann cells.49 This hypothesis led to the testing of the broad anti-fibrotic pirfenidone, which has proven efficacy for idiopathic pulmonary fibrosis, in Phase 2 trials in NF1 patients with advanced pNF but, unfortunately, no benefit of pirfenidone was reported50 (NCT00076102). However, it is not clear whether the trial was unsuccessful because pirfenidone was not effective at reducing collagen deposition or because collagen type I is not an effective drug target in pNF. Further investigation is needed to definitively address the contribution of collagen to NF1-related tumour development.
Current therapeutics strategies under development
The existence of mouse models in the NF1 arena (Box 2) has provided unprecedented opportunities to derive important insights into the biology of the disease as well as preclinical models that guide the development of effective therapies for NF1 (Fig. 2)

Overview of current therapies under investigation for the most common types of tumour associated with NF1, including inhibitors of receptor tyrosine kinases (RTKs) upstream of RAS (capmatinib), components of the pathway downstream of RAS (MEK inhibitors), immune-checkpoint inhibitors (ipilimumab, nivolumab, pembrolizumab) and BET inhibitors.
Downstream of Ras: mitogen-activated protein kinase kinase inhibitors
Downstream of Ras activation, the extracellular-signal regulated kinase/mitogen-activated protein kinase (ERK/MAPK) signalling pathway relays proliferation and cell-cycle entry signals. This pathway is hyperactivated in NF1-related tumours, and inhibition of MAPK kinase (MEK) was effective at controlling tumour size in a mouse model of NF1.51,52 Unlike other therapeutic strategies tested so far, MEK inhibition (using selumetinib) has yielded unprecedented results in the clinic for pNF.53 In fact, almost all selumetinib-treated patients showed no tumour progression while being treated with the drug, with 72% showing a partial response and 24% showing stable disease in the SPRINT trial (NCT01362803). Based on these promising results, the FDA recently approved selumetinib for children with symptomatic, progressive and inoperable pNFs. A small cohort of children with OPG were similarly responsive to MEK inhibition (NCT01089101)54 but equivalent strategies in the malignant context (in MPNST) have so far shown only moderate efficacy in mouse models.55 Of note, it has been reported that the RTK MET is activated in some MPNSTs and that a mouse model recapitulating MET activation in MPNST is sensitive to MET inhibition;56 it would therefore be interesting to test a variety of MEK combination therapies such as dual MET–MEK inhibition in the context of MPNSTs.56
Inhibitors of bromodomain-containing protein 4
A comparative transcriptome analysis of mouse MPNSTs and their cells of origin revealed that the gene encoding bromodomain-containing protein 4 (Brd4) is one of the most highly upregulated genes in MPNSTs.57 BRD4 is a chromatin regulator that contains two bromodomains and an extended terminal domain characteristic of the BET family of bromodomain proteins. It binds to acetylated histone (H3K27Ac) and facilitates the expression of mitotic genes. Its downregulation or inhibition through the selective small inhibitor JQ-1 drastically decreases the growth of many types of cancer cell,58 including MPNST cells in vitro and in vivo.57 As many MPNSTs in NF1 patients have mutations in components of the PRC2 complex, which negatively regulates gene expression through histone methylation (H3K27Me) resulting in histone acetylation (H3K27Ac), inhibiting BRD4 might be particularly effective.59 This observation prompted the launch of a Phase 2 clinical trial of a small molecule inhibitor of BET domain proteins for MPNST, but the study was discontinued due to a lack of participants (NCT02986919). However, another study of a BET inhibitor plus a MEK inhibitor plus an inhibitor of programmed death-ligand (PD-L1) (see below) for MPNST is being carried out by the NF Clinical Trials Consortium.
T cells
The discovery that cancer can escape the cytotoxic capacity of T cells through checkpoint strategies has great potential to revolutionise the field of cancer therapeutics but, despite the clinical success of immune checkpoint inhibitors,60,61 T cells have received little attention in the context of NF1. Immune profiling of neurofibroma and MPNSTs indicates considerable variation in the extent of infiltration of CD8+ T cells as well as in the expression level of the T-cell inhibitory ligand PD-L1;62,63 50% of gliomas contain a high number of T cells.64 Although the influence of T cells on neurofibroma formation has not been directly evaluated in NF1-related tumours, these cells are emerging as essential components of the stroma that support tumour growth.65,66 Accordingly, clinical trials targeting T cells using a combination of antibodies against the receptor for PD-L1 (PD-1) and cytotoxic T-lymphocyte-associated protein 4 (CTLA4, another immune checkpoint protein) are currently ongoing (e.g. NCT02834013) in a Phase 2 trial for patients with rare tumours (including NF1-related tumours), although reliable markers predicting the response to immunotherapies remain to be identified in NF1 preclinical models.67
Immunocompromised (T-cell-deficient) mouse models are commonly used in cancer biology to perform xenograft studies. Intriguingly, malignant glioma cells successfully graft in immunocompromised mice whereas benign glioma cells fail to engraft.65 Pan and colleagues65 discovered that the cause of the failure of benign cells to engraft is due to impaired microglia function, including reduced expression of the chemokine receptor Ccr2 and its ligand Ccl5 in the immunocompromised mice; accordingly, mice deficient in Ccl5 phenocopy T-cell-deficiency, as gliomas also fail to engraft in Ccl5 knockout mice. Importantly, T cell exposure restores Ccr2 and Ccl5 expression in athymic microglia.65 These observations suggest that T cells support benign glioma growth but impair malignant transformation. It is tempting to speculate that a similar mechanism occurs in humans, because human MPNST cells can be grown in athymic mice57 but a xenograft model with human neurofibroma cells has not so far been reported, indicating that dampening T-cell function can restrain benign tumour development whereas enhancing T-cell function, on the other hand, can be used to prevent malignant transformation.
Advances in basic research that could lead to the generation of novel therapeutics
Recent advances in our understanding of tumour molecular and cellular pathogenesis will provide opportunities to develop effective treatments in the future for NF1-associated neoplasms (Fig. 3).

Overview of the potential future therapies for the most common types of tumour associated with NF1, including targeting the Hippo pathway, JAK/STAT signalling pathway and oestrogen signalling as well as components of the NF1+/− tumour microenvironment (fibroblasts, nerve cells, macrophages and other immune cells).
The Hippo pathway
As a central regulator of cell growth and size, it is not surprising that the Hippo pathway is involved in a number of tumour types, including NF2-associated tumours.68 The NF2 disease, characterised by highly penetrant tumours of the vestibular nerve called vestibular schwannomas, is caused by biallelic inactivation of the NF2 gene. The protein encoded by the NF2 gene, Merlin, functions as an upstream initiator of the core kinase signalling cascade involving MST1/2, LATS1/2 and the transcription factors YAP and TAZ. Interestingly, biallelic inactivation of the NF2 gene leads to some features of NF1, such as cNF.69,70 Indeed, in genetically engineered mouse models of cNF and MPNST, the Hippo pathway acts as a modifier3 and a driver,71 respectively, suggesting that selectively dampening the Hippo pathway using inhibitors could be an attractive therapeutic approach, and several novel small molecule YAP inhibitors are currently in preclinical development.72 However, the fraction of NF1 patients that have an aberrant Hippo pathway and that, hence, might be sensitive to inhibition of the Hippo pathway, remains to be established.3,73,74,75
The Janus kinase/signal transducer and activator of transcription pathway
Signalling through the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) pathway is known to contribute to tumorigenesis in numerous ways.76 Using a high-throughput chemical library screen, Banerjee and colleagues discovered that STAT3 is hyperactivated in human glioma and MPNST cell lines.77 As EGFR is one of the tyrosine kinases that mediates STAT3 phosphorylation78 and is expressed in most human MPNSTs,26 researchers investigated, both genetically and pharmacologically, the contribution of STAT3 signalling in mouse models of pNF and MPNST.79,80,81 In brief, interfering with the STAT3 pathway delayed neurofibroma initiation80 as well as progression to MPNST,81 but had far less impact on tumour maintenance. The results of these preclinical studies suggest that inhibiting STAT3 might be of benefit to NF1 patients by preventing/delaying the development of pNFs. The safety and overall effect(s) of long-term STAT3 inhibition in humans are currently unknown, but data from early clinical trials indicate that antisense oligonucleotides that target STAT3 mRNA are well tolerated in the short term.82
The tyrosine kinase TYK2, also a member of the JAK family (in addition to JAK1–3), is emerging as an attractive target in the context of MPNST. This protein associates with the cytoplasmic domain of type I and type II cytokine receptors to ultimately activate downstream oncogenic signals.76 Overexpression of TYK2 was found in approximately 60% of MPNST cases but not in normal peripheral nerve or benign neurofibromas.83 In vitro, knockdown of TYK2 in murine and human MPNST cells significantly increased cell death, while knockdown in vivo led to a decreased tumour volume in a subcutaneous mouse model, supporting a role for TYK2 in driving MPNSTs.84 Importantly, orally available TYK2 inhibitors have been developed for the treatment of autoimmune disorders, and have so far proven to be clinically safe.85,86 It remains to be determined, however, if interfering with TYK2 expression and/or downstream targets in vivo will impair MPNST initiation and/or maintenance.
Oestrogen signalling
The development of cNFs in NF1 patients coincides with puberty. Additionally, as discussed by Brosseau et al.6, sex hormones have been proposed to influence the growth of tumours in NF1 patients. However, direct evidence is still lacking. The results of a study in an autologous mouse model suggest that pregnancy might favour the development of cNF,87 while OPG-induced retinal ganglion loss and retinal nerve fibre layer thinning in mice were reported to depend on the presence of ovaries.88 In humans, female NF1 patients are reported to be at a higher risk than the general population of developing breast cancer.89 Results from several studies point to a correlation between the NF1 and oestrogen receptor status in multiple model systems. Oestrogen-receptor-positive breast tumours harbouring a sporadic NF1 mutation have a poor outcome,90 and depletion of oestrogen through ovariectomy in a NF1-loss-driven rat model of breast cancer leads to tumour regression.91 Breast cancer patients with NF1 sporadic mutations treated with the oestrogen-receptor antagonist fulvestrant showed a good outcome,92 suggesting the potential use of fulvestrant in the context of NF1 patients. Interestingly, it has been shown that neurofibromin directly regulates the oestrogen receptor.93 Thus, further elucidating how neurofibromin modulates oestrogen signalling and vice versa might shed light on the role of sex hormones in regulating tumour growth in NF1 patients, and ultimately provide a rationale for repurposing agents that target oestrogen in order to inhibit neurofibroma development.
Nerve cells
As demonstrated by Liao et al.94, the nerve microenvironment plays a key role in initiating neurofibroma formation but the identity of the factors responsible for this process are largely unknown. Neuregulin-1, a Schwann cell growth factor secreted by nerve cells,95 binds to its receptors—EGFR and HER2—on Schwann cells to mediate cell growth and migration; notably, overexpression of either neuregulin-196 or EGFR27 is sufficient to drive neurofibroma and MPNST tumorigenesis in mice.27 Therefore, a potential therapeutic strategy might be to target this neuregulin-1–EGFR axis,97 perhaps by repurposing the breast cancer drug trastuzumab.
Macrophages
pNF-associated macrophages have become a focus in the study of the neurofibroma tumour microenvironment. Indeed microglia were found to be critical to glioma development98 and macrophages are abundant in neurofibromas and MPNSTs.99,100 These macrophages are recruited by macrophage colony-stimulating factor (M-CSF).100 Pharmaceutical inhibition of macrophages has been shown to slow down pNF progression.100 Therefore, in addition to tumorous glial cells, these microenvironmental macrophages are also potential therapeutic targets for neurofibroma and glioma.
In the context of OPG, Daginakatte et al.17,18 have investigated the role of microglia pharmacologically, using a hyaluronidase inhibitor, and genetically, using established mouse models. Their results indicate that microglia promote the growth of OPG and therefore suggest that the development of strategies targeting stromal cells such as microglia might be pursued.17,18,98 This same research group discovered that the expression of numerous cytokines, chemokines and their receptors is altered in microglia in the context of OPG. Indeed, the chemokine ligand CX3CL1 is expressed at abundant levels in the optic nerve and retina, and the expression of CX3CR1 in microglia has been shown to be required for optic glioma formation in vivo.19 Monoallelic inactivation of Cx3cr1 is sufficient to significantly reduce the growth of OPG,19 but the use of available small molecule inhibitors against CX3CR1 has so far not been reported. Ccl5 is also highly expressed in microglia and a neutralising antibody against this chemokine reduced OPG growth and improved retinal dysfunction in vivo in mice;20 however, antagonising the CCL5 receptors CCR1 and CCR5, which are expressed on the surface of astrocytes, yielded conflicting results as it had no effect on OPG growth in a mouse model of NF1.20 Therefore, the mechanism by which CCL5 exerts its pro-tumorigenic role in the context of OPG remains to be demonstrated.
The absolute role of macrophages in tumorigenesis in the NF1 context is not clear despite their abundant presence in pNF and even more so in MPNSTs in both mice and humans.100 Inhibiting the receptor for macrophage M-CSF using PLX3397 successfully decreased macrophage density and promoted tumour regression in a fraction of the mice tested, but only once pNFs had been established. Earlier treatment, prior to neurofibroma formation, increased tumour volume compared with controls, indicating that macrophages are likely to play a defensive role during tumour development but a permissive role once tumours are established.100 However, PLX3397 does not selectively inhibit macrophage function: it also inhibits the kinase receptor c-kit, an important positive modulator of neurofibroma growth, on mast cells.32 Interestingly, the STAT3 pathway is also hyperactivated in neurofibroma-associated macrophages as well as in neoplastic Schwann cells.101 Therefore, the identity of which cells and pathways within the tumour that contribute to the reduced growth of pNFs in response to PLX3397 is unclear. To characterise the neurofibroma-associated macrophages, Choi et al. performed high-throughput gene expression analysis, which confirmed the high expression of ccl5 in these cells, reinforcing the need to better understand its downstream effectors.102 An elevated macrophage number in neurofibroma and nerve requires the express the Cxcl10 chemokine receptor Cxcr3 and, notably, Dhh-Cre; Nf1f/f mice failed to develop neurofibromas in a Cxcr3-null background,66 suggesting a critical role for the Cxcl10–Cxcr3 chemokine axis in macrophage recruitment for neurofibroma formation. Interestingly, Choi et al.102 also discovered a switch from interferon (IFN) type I to type II signalling during the development of neurofibromas, which is consistent with the subsequent modulation of downstream targets of IFN-γ such as colony-stimulating factor (CSF)1.100,103 Remarkably, pegylated IFN-α2b significantly slowed down neurofibroma growth in some NF1 patients, warranting further investigation.104
In summary, there is a need to better assess the definitive contribution of macrophages to the formation of neurofibromas and MPNSTs in vivo and to pursue the development of strategies that specifically target these cells in future clinical investigation.
The NF1+/− tumour microenvironment
On average, patients with hereditary cancer syndromes develop tumours at a much younger age than the general population, typically due to a predisposing mutation in a tumour-suppressor gene. Importantly, in addition to the intrinsic tumorigenic drive generated by the biallelic inactivation of this tumour-suppressor gene in the neoplastic cells, it has been suggested that a heterozygous tumour-suppressor microenvironment also modulates tumour progression.105,106 In the context of neurofibromatosis, mast cells32 and nerve cells94 have been shown to accelerate neurofibroma development in vivo. Mast-cell infiltration has been recognised as a histological feature for neurofibroma,10 as a high density of mast cells is uncommon in most other tumour tissues. Our recent study showed that pNF-associated mast cells are recruited by tumorous Schwann cell-derived stem cell factor; however, removal of this factor source only slightly affects neurofibroma progression.11 This observation suggests that either mast cells might not play an essential role in sustaining pNF tumorigenesis or other immune cells in the heterozygous tumour microenvironment compensate for the loss of mast cell function in this specific murine model. In fact, the roles of macrophages and T cells in promoting NF1-related tumour development have also been shown.66,100 Taken together, the coexistence of multiple types of immune cell suggests a complex inflammatory microenvironment in neurofibroma, which could be associated with nerve injury99 to promote tumorigenesis. These studies highlight a positive influence of the microenvironment on tumour formation. By contrast, however, Brosseau et al. demonstrated that the Nf1+/– microenvironment can impair malignant progression.107 This result is consistent with the long-standing and puzzling clinical observation of the low malignant potential of the characteristic benign lesions of NF1 patients (e.g. iris hamartomas, café-au-lait macules, cNF). Several mechanisms might account for the impaired malignant progression. One possibility is that heterozygous tumour-suppressor genes might lead to immune cell hyperproliferation and function and, thus, increased immune surveillance, to prevent cancer development.107 Consistent with this view, it has been reported that Nf1+/− T cells trigger an enhanced immune reaction107 and that Nf1+/− natural killer cells have a superior protective effect than their wild-type counterparts.108 This scenario could have serious clinical implications for organ transplant recipients who coincidentally have NF1, as these patients would have a drastically increased likelihood of developing malignant tumours on treatment with immunosuppressive drugs to prevent transplant rejection. As proof of principle, an association between some autoimmune diseases and NF1 has been reported,109 most likely as a result of enhanced immune function due to NF1 heterozygosity. Initially, this hypothetical model seems to contrast with reports from Dodd et al.110 and Hirbe et al.111 that a Nf1+/– microenvironment accelerates MPNST formation. However, the findings can be reconciled by taking into account the fact that the mouse models used by Dodd et al.110 and Hirbe et al.111 do not recapitulate the physiological stepwise progression from benign to malignant tumour in human NF1 patients, as is the case in Brosseau et al.107. Importantly, this novel working model suggests that identifying the cell types and signalling pathways that underlie the capacity of the microenvironment to suppress malignant progression could open the door to novel treatments for cancer beyond NF1.
NF1 gene therapy
Research in other monogenic diseases such as muscular dystrophy indicates that even a partial functional rescue of the mutated protein confers patient clinical benefit,112 but such a demonstration has so far only been carried out in vitro for NF1.113 This is due to the fact that NF1 is one of the largest genes and its manipulation into expression vectors has proved very challenging. However, given the 2019 report of the successful adeno-associated viral transduction of truncated forms of NF1 into human cells,114 the cloning of a full-length Nf1 cDNA that is able to modulate Ras signalling,115 and opening of a proposal for a proof-of-concept of NF1 gene therapy by the Children’s Tumor Foundation and the Gilbert Foundation, the NF1 community is optimistic about novel therapeutics aimed at restoring the function of neurofibromin to correct the disease at its root in order to ultimately reduce the tumour burden in NF1 patients.
Conclusions
NF1 is a hereditary tumour syndrome with poor genotype–phenotype correlation and, hence, unpredictable outcome. Although earlier work has focused on interfering with signalling pathways upstream and downstream of Ras (Fig. 1), current clinical trials are evaluating the efficacy of BRD4 inhibitors and immune checkpoint blockers (Fig. 2). So far, the best results have been obtained by inhibiting MEK (downstream of Ras), but these results are currently limited to pNF and low-grade glioma. Future work is aimed at deciphering the contribution of the Hippo pathway, the JAK/STAT pathway and oestrogen signalling, as well as interfering with cells in the microenvironment that modulate tumour progression, such as nerve cells, T cells, macrophages and Nf1+/– stromal cells (Fig. 3).
Combination therapies targeting both neoplastic cells (Schwann cells) and cells of the tumour microenvironment might be more effective than drugs targeting only the tumour cells. Indeed, the NF1 heterozygous cells of the microenvironment modulate the growth of pNFs,116 OPGs16 and MPNSTs.107 As the tumour mass can contain up to 50% collagen,48 strategies aimed at targeting the cells that synthesise collagen and extracellular matrix might be required to shrink the bulk of the tumour, in addition to directly targeting the neoplastic cells. Therefore, further elucidating of the molecular interactions between Schwann cells and their tumour microenvironment will provide new tools and knowledge to develop more effective treatments for one of the most debilitating human genetic disorders in the world today.
Review criteria
Published research papers, clinical trials, and literature reviews were selected using different keywords (e.g., NF1, neurofibroma, malignant peripheral nerve sheath tumour, optic glioma) and database sources (e.g., PubMed, Google Scholar, ISI Proceedings, Journal storage [JSTOR] Search, Medline, Scopus, Web of Science, Clinicaltrials.gov).
References
Friedman, J. M. Neurofibromatosis 1. in GeneReviews((R)) (eds Adam, M. P., Ardinger, H. H., Pagon, R. A., Wallace, S. E., Bean, L. J. H., Stephens, K. et al.). (University of Washington Seattle, 1993).
Boyd, K. P., Korf, B. R. & Theos, A. Neurofibromatosis type 1. J. Am. Acad. Dermatol 61, 1–14 (2009).
Chen, Z., Mo, J., Brosseau, J. P., Shipman, T., Wang, Y. Liao, C. P. et al. Spatiotemporal loss of NF1 in Schwann cell lineage leads to different types of cutaneous neurofibroma susceptible to modification by the hippo pathway. Cancer Disco. 9, 114–129. (2019).
Klar, N., Cohen, B. & Lin, D. D. M. Neurocutaneous syndromes. Handb. Clin. Neurol. 135, 565–589. (2016).
Hirbe, A. C. & Gutmann, D. H. Neurofibromatosis type 1: a multidisciplinary approach to care. Lancet Neurol. 13, 834–843 (2014).
Brosseau, J. P., Pichard, D. C., Legius, E. H., Wolkenstein, P., Lavker, R. M., Blakeley, J. O. et al. The biology of cutaneous neurofibromas: consensus recommendations for setting research priorities. Neurology 91(2 Suppl. 1), S14–S20 (2018).
Gosline, S. J., Weinberg, H., Knight, P., Yu, T., Guo, X., Prasad, N. et al. A high-throughput molecular data resource for cutaneous neurofibromas. Sci. Data. 4, 170045 (2017).
Pemov, A., Li, H., Patidar, R., Hansen, N. F., Sindiri, S. Hartley, S. W. et al. The primacy of NF1 loss as the driver of tumorigenesis in neurofibromatosis type 1-associated plexiform neurofibromas. Oncogene 36, 3168–3177. (2017).
Miettinen, M. M., Antonescu, C. R., Fletcher, C. D. M., Kim, A., Lazar, A. J., Quezado, M. M. et al. Histopathologic evaluation of atypical neurofibromatous tumors and their transformation into malignant peripheral nerve sheath tumor in patients with neurofibromatosis 1-a consensus overview. Hum. Pathol. 67, 1–10 (2017).
Carrio, M., Gel, B., Terribas, E., Zucchiatti, A. C., Moline, T., Rosas, I. et al. Analysis of intratumor heterogeneity in Neurofibromatosis type 1 plexiform neurofibromas and neurofibromas with atypical features: correlating histological and genomic findings. Hum. Mutat. 39, 1112–1125 (2018).
Rhodes S. D., He Y., Smith A., Jiang L., Lu Q., Mund J. et al. Cdkn2a (Arf) loss drives NF1-associated atypical neurofibroma and malignant transformation. Hum. Mol. Genet. 28, 2752–2762 (2019).
Lee, W., Teckie, S., Wiesner, T., Ran, L., Prieto Granada, C. N., Lin, M. et al. PRC2 is recurrently inactivated through EED or SUZ12 loss in malignant peripheral nerve sheath tumors. Nat. Genet. 46, 1227–1232 (2014).
Evans, D. G., Baser, M. E., McGaughran, J., Sharif, S., Howard, E. & Moran, A. Malignant peripheral nerve sheath tumours in neurofibromatosis 1. J. Med Genet 39, 311–314 (2002).
Dodgshun, A. J., Elder, J. E., Hansford, J. R. & Sullivan, M. J. Long-term visual outcome after chemotherapy for optic pathway glioma in children: Site and age are strongly predictive. Cancer 121, 4190–4196 (2015).
Campen, C. J. & Gutmann, D. H. Optic pathway gliomas in neurofibromatosis type 1. J. Child Neurol. 33, 73–81 (2018).
Bajenaru, M. L., Hernandez, M. R., Perry, A., Zhu, Y., Parada, L. F., Garbow, J. R. et al. Optic nerve glioma in mice requires astrocyte Nf1 gene inactivation and Nf1 brain heterozygosity. Cancer Res. 63, 8573–8577 (2003).
Daginakatte, G. C., Gianino, S. M., Zhao, N. W., Parsadanian, A. S. & Gutmann, D. H. Increased c-Jun-NH2-kinase signaling in neurofibromatosis-1 heterozygous microglia drives microglia activation and promotes optic glioma proliferation. Cancer Res. 68, 10358–10366 (2008).
Daginakatte, G. C. & Gutmann, D. H. Neurofibromatosis-1 (Nf1) heterozygous brain microglia elaborate paracrine factors that promote Nf1-deficient astrocyte and glioma growth. Hum. Mol. Genet 16, 1098–1112 (2007).
Pong, W. W., Higer, S. B., Gianino, S. M., Emnett, R. J. & Gutmann, D. H. Reduced microglial CX3CR1 expression delays neurofibromatosis-1 glioma formation. Ann. Neurol. 73, 303–308 (2013).
Solga, A. C., Pong, W. W., Kim, K. Y., Cimino, P. J., Toonen, J. A., Walker, J. et al. RNA Sequencing of tumor-associated microglia reveals Ccl5 as a stromal chemokine critical for neurofibromatosis-1 glioma growth. Neoplasia 17, 776–788 (2015).
Block, M. L., Zecca, L. & Hong, J. S. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat. Rev. Neurosci. 8, 57–69 (2007).
Prior, I. A., Lewis, P. D. & Mattos, C. A comprehensive survey of Ras mutations in cancer. Cancer Res. 72, 2457–2467 (2012).
Cox, A. D., Fesik, S. W., Kimmelman, A. C., Luo, J. & Der, C. J. Drugging the undruggable RAS: mission possible? Nat. Rev. Drug Disco. 13, 828–851 (2014).
O’Bryan, J. P. Pharmacological targeting of RAS: recent success with direct inhibitors. Pharm. Res. 139, 503–511 (2019).
Widemann, B. C., Dombi, E., Gillespie, A., Wolters, P. L., Belasco, J., Goldman, S. et al. Phase 2 randomized, flexible crossover, double-blinded, placebo-controlled trial of the farnesyltransferase inhibitor tipifarnib in children and young adults with neurofibromatosis type 1 and progressive plexiform neurofibromas. Neuro Oncol. 16, 707–718 (2014).
DeClue, J. E., Heffelfinger, S., Benvenuto, G., Ling, B., Li, S. Rui, W. et al. Epidermal growth factor receptor expression in neurofibromatosis type 1-related tumors and NF1 animal models. J. Clin. Invest. 105, 1233–1241 (2000).
Ling, B. C., Wu, J., Miller, S. J., Monk, K. R., Shamekh, R., Rizvi, T. A. et al. Role for the epidermal growth factor receptor in neurofibromatosis-related peripheral nerve tumorigenesis. Cancer Cell. 7, 65–75 (2005).
Albritton, K. H., Rankin, C., Coffin, C. M., Ratner, N., Budd, G. T., Schuetze, S. M. et al. Phase II study of erlotinib in metastatic or unresectable malignant peripheral nerve sheath tumors (MPNST). J. Clin. Oncol. 24, 9518 (2006).
Kadono, T., Kikuchi, K., Nakagawa, H. & Tamaki, K. Expressions of various growth factors and their receptors in tissues from neurofibroma. Dermatology 201, 10–14 (2000).
Badache, A. & De Vries, G. H. Neurofibrosarcoma-derived Schwann cells overexpress platelet-derived growth factor (PDGF) receptors and are induced to proliferate by PDGF BB. J. Cell Physiol. 177, 334–342 (1998).
Rao, U. N., Sonmez-Alpan, E. & Michalopoulos, G. K. Hepatocyte growth factor and c-MET in benign and malignant peripheral nerve sheath tumors. Hum. Pathol. 28, 1066–1070 (1997).
Yang, F. C., Ingram, D. A., Chen, S., Zhu, Y., Yuan, J., Li, X. et al. Nf1-dependent tumors require a microenvironment containing Nf1+/- and c-kit-dependent bone marrow. Cell 135, 437–448 (2008).
Karajannis, M. A., Legault, G., Fisher, M. J., Milla, S. S., Cohen, K. J., Wisoff, J. H. et al. Phase II study of sorafenib in children with recurrent or progressive low-grade astrocytomas. Neuro Oncol. 16, 1408–1416 (2014).
Robertson, K. A., Nalepa, G., Yang, F. C., Bowers, D. C., Ho, C. Y., Hutchins, G. D. et al. Imatinib mesylate for plexiform neurofibromas in patients with neurofibromatosis type 1: a phase 2 trial. Lancet Oncol. 13, 1218–1224 (2012).
Chugh, R., Wathen, J. K., Maki, R. G., Benjamin, R. S., Patel, S. R., Meyers, P. A. et al. Phase II multicenter trial of imatinib in 10 histologic subtypes of sarcoma using a bayesian hierarchical statistical model. J. Clin. Oncol. 27, 3148–3153 (2009).
Friedrich, R. E., Holstein, A. F., Middendorff, R. & Davidoff, M. S. Vascular wall cells contribute to tumourigenesis in cutaneous neurofibromas of patients with neurofibromatosis type 1. A comparative histological, ultrastructural and immunohistochemical study. Anticancer Res. 32, 2139–2158 (2012).
Kawachi, Y., Xu, X., Ichikawa, E., Imakado, S. & Otsuka, F. Expression of angiogenic factors in neurofibromas. Exp. Dermatol. 12, 412–417 (2003).
Wasa, J., Nishida, Y., Suzuki, Y., Tsukushi, S., Shido, Y., Hosono, K. et al. Differential expression of angiogenic factors in peripheral nerve sheath tumors. Clin. Exp. Metastasis 25, 819–825 (2008).
Angelov, L., Salhia, B., Roncari, L., McMahon, G. & Guha, A. Inhibition of angiogenesis by blocking activation of the vascular endothelial growth factor receptor 2 leads to decreased growth of neurogenic sarcomas. Cancer Res. 59, 5536–5541 (1999).
Maki, R. G., D’Adamo, D. R., Keohan, M. L., Saulle, M., Schuetze, S. M., Undevia, S. D. et al. Phase II study of sorafenib in patients with metastatic or recurrent sarcomas. J. Clin. Oncol. 27, 3133–3140 (2009).
Avery, R. A., Hwang, E. I., Jakacki, R. I. & Packer, R. J. Marked recovery of vision in children with optic pathway gliomas treated with bevacizumab. JAMA Ophthalmol. 132, 111–114 (2014).
Johannessen, C. M., Reczek, E. E., James, M. F., Brems, H., Legius, E. & Cichowski, K. The NF1 tumor suppressor critically regulates TSC2 and mTOR. Proc. Natl Acad. Sci. USA 102, 8573–8578 (2005).
Hegedus, B., Banerjee, D., Yeh, T. H., Rothermich, S., Perry, A., Rubin, J. B. et al. Preclinical cancer therapy in a mouse model of neurofibromatosis-1 optic glioma. Cancer Res. 68, 1520–1528 (2008).
Weiss, B., Widemann, B. C., Wolters, P., Dombi, E., Vinks, A. A., Cantor, A. et al. Sirolimus for non-progressive NF1-associated plexiform neurofibromas: an NF clinical trials consortium phase II study. Pediatr. Blood Cancer 61, 982–986 (2014).
Weiss, B., Widemann, B. C., Wolters, P., Dombi, E., Vinks, A., Cantor, A. et al. Sirolimus for progressive neurofibromatosis type 1-associated plexiform neurofibromas: a neurofibromatosis Clinical Trials Consortium phase II study. Neuro Oncol. 17, 596–603 (2015).
Widemann B. C., Meyer C. F., Cote G. M., Chugh R., Milhem M. M., Tine B. A. V. et al. SARC016: Phase II study of everolimus in combination with bevacizumab in sporadic and neurofibromatosis type 1 (NF1) related refractory malignant peripheral nerve sheath tumors (MPNST). J. Clin. Oncol. 34, 11053 (2016).
Slopis, J. M., Arevalo, O., Bell, C. S., Hebert, A. A., Northrup, H., Riascos, R. F. et al. Treatment of disfiguring cutaneous lesions in neurofibromatosis-1 with Everolimus: a phase II, open-label, single-arm trial. Drugs R. D. 18, 295–302 (2018).
Peltonen, J., Penttinen, R., Larjava, H. & Aho, H. J. Collagens in neurofibromas and neurofibroma cell cultures. Ann. N. Y. Acad. Sci. 486, 260–270 (1986).
Yang, F. C., Chen, S., Clegg, T., Li, X., Morgan, T., Estwick, S. A. et al. Nf1+/- mast cells induce neurofibroma like phenotypes through secreted TGF-beta signaling. Hum. Mol. Genet. 15, 2421–2437 (2006).
Widemann, B. C., Babovic-Vuksanovic, D., Dombi, E., Wolters, P. L., Goldman, S., Martin, S. et al. Phase II trial of pirfenidone in children and young adults with neurofibromatosis type 1 and progressive plexiform neurofibromas. Pediatr. Blood Cancer 61, 1598–602. (2014).
Chen, Z., Liu, C., Patel, A. J., Liao, C. P., Wang, Y. & Le, L. Q. Cells of origin in the embryonic nerve roots for NF1-associated plexiform neurofibroma. Cancer Cell. 26, 695–706 (2014).
Jousma, E., Rizvi, T. A., Wu, J., Janhofer, D., Dombi, E. Dunn, R. S. et al. Preclinical assessments of the MEK inhibitor PD-0325901 in a mouse model of Neurofibromatosis type 1. Pediatr. Blood Cancer 62, 1709–1716 (2015).
Dombi, E., Baldwin, A., Marcus, L. J., Fisher, M. J., Weiss, B. Kim, A. et al. Activity of Selumetinib in Neurofibromatosis Type 1-related plexiform neurofibromas. N. Engl. J. Med. 375, 2550–2560 (2016).
Kondyli, M., Larouche, V., Saint-Martin, C., Ellezam, B., Pouliot, L. Sinnett, D. et al. Trametinib for progressive pediatric low-grade gliomas. J. Neurooncol. 140, 435–444 (2018).
Malone, C. F., Fromm, J. A., Maertens, O., DeRaedt, T., Ingraham, R. & Cichowski, K. Defining key signaling nodes and therapeutic biomarkers in NF1-mutant cancers. Cancer Discov. 4, 1062–1073 (2014).
Peacock, J. D., Pridgeon, M. G., Tovar, E. A., Essenburg, C. J., Bowman, M. Madaj, Z. et al. Genomic status of MET potentiates sensitivity to MET and MEK inhibition in NF1-related malignant peripheral nerve sheath tumors. Cancer Res. 78, 3672–3687 (2018).
Patel, A. J., Liao, C. P., Chen, Z., Liu, C., Wang, Y. & Le, L. Q. BET bromodomain inhibition triggers apoptosis of NF1-associated malignant peripheral nerve sheath tumors through Bim induction. Cell Rep. 6, 81–92 (2014).
Xu, Y. & Vakoc, C. R. Targeting cancer cells with BET bromodomain inhibitors. Cold Spring Harb. Perspect. Med. 7, 1–10 (2017).
De Raedt, T., Beert, E., Pasmant, E., Luscan, A., Brems, H. Ortonne, N. et al. PRC2 loss amplifies Ras-driven transcription and confers sensitivity to BRD4-based therapies. Nature 514, 247–251 (2014).
Hamid, O., Robert, C., Daud, A., Hodi, F. S., Hwu, W. J., Kefford, R. et al. Safety and tumor responses with lambrolizumab (anti-PD-1) in melanoma. N. Engl. J. Med. 369, 134–144 (2013).
Le, D. T., Uram, J. N., Wang, H., Bartlett, B. R., Kemberling, H., Eyring, A. D. et al. PD-1 blockade in tumors with mismatch-repair deficiency. N. Engl. J. Med. 372, 2509–2520 (2015).
Haworth, K. B., Arnold, M. A., Pierson, C. R., Choi, K., Yeager, N. D. Ratner, N. et al. Immune profiling of NF1-associated tumors reveals histologic subtype distinctions and heterogeneity: implications for immunotherapy. Oncotarget 8, 82037–82048 (2017).
Shurell, E., Singh, A. S., Crompton, J. G., Jensen, S., Li, Y., Dry, S. et al. Characterizing the immune microenvironment of malignant peripheral nerve sheath tumor by PD-L1 expression and presence of CD8+ tumor infiltrating lymphocytes. Oncotarget 7, 64300–64308 (2016).
D’Angelo, F., Ceccarelli, M., Tala., Garofano, L., Zhang, J. Frattini, V. et al. The molecular landscape of glioma in patients with Neurofibromatosis 1. Nat. Med. 25, 176–187 (2019).
Pan, Y., Xiong, M., Chen, R., Ma, Y., Corman, C., Maricos, M. et al. Athymic mice reveal a requirement for T-cell-microglia interactions in establishing a microenvironment supportive of Nf1 low-grade glioma growth. Genes Dev. 32, 491–496 (2018).
Fletcher, J. S., Wu, J., Jessen, W. J., Pundavela, J., Miller, J. A., Dombi, E. et al. Cxcr3-expressing leukocytes are necessary for neurofibroma formation in mice. JCI Insight. 4, 1–16 (2019).
Havel, J. J., Chowell, D. & Chan, T. A. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat. Rev. Cancer. 19, 133–150 (2019).
Harvey, K. F., Zhang, X. & Thomas, D. M. The Hippo pathway and human cancer. Nat. Rev. Cancer 13, 246–257 (2013).
Parry, D. M., Eldridge, R., Kaiser-Kupfer, M. I., Bouzas, E. A., Pikus, A. & Patronas, N. Neurofibromatosis 2 (NF2): clinical characteristics of 63 affected individuals and clinical evidence for heterogeneity. Am. J. Med. Genet. 52, 450–461 (1994).
Evans, D. G. Neurofibromatosis type 2 (NF2): a clinical and molecular review. Orphanet J. Rare Dis. 4, 16 (2009).
Wu, L. M. N., Deng, Y., Wang, J., Zhao, C., Wang, J. Rao, R. et al. Programming of schwann cells by Lats1/2-TAZ/YAP signaling drives malignant peripheral nerve sheath tumorigenesis. Cancer Cell. 33, 292–308.e7 (2018).
Woodard, G. A., Yang, Y. L., You, L. & Jablons, D. M. Drug development against the hippo pathway in mesothelioma. Transl. Lung Cancer Res. 6, 335–342 (2017).
Kim, Y. H., Ohta, T., Oh, J. E., Le Calvez-Kelm, F., McKay, J., Voegele, C. et al. TP53, MSH4, and LATS1 germline mutations in a family with clustering of nervous system tumors. Am. J. Pathol. 184, 2374–2381 (2014).
Oh, J. E., Ohta, T., Satomi, K., Foll, M., Durand, G., McKay, J. et al. Alterations in the NF2/LATS1/LATS2/YAP pathway in schwannomas. J. Neuropathol. Exp. Neurol. 74, 952–959 (2015).
Faden, D. L., Asthana, S., Tihan, T., DeRisi, J. & Kliot, M. Whole exome sequencing of growing and non-growing cutaneous neurofibromas from a single patient with neurofibromatosis type 1. PLoS ONE 12, e0170348 (2017).
Woss K., Simonovic N., Strobl B., Macho-Maschler S., Muller M. TYK2: an upstream kinase of STATs in Cancer. Cancers (Basel). 11, 1–18 (2019).
Banerjee, S., Byrd, J. N., Gianino, S. M., Harpstrite, S. E., Rodriguez, F. J., Tuskan, R. G. et al. The neurofibromatosis type 1 tumor suppressor controls cell growth by regulating signal transducer and activator of transcription-3 activity in vitro and in vivo. Cancer Res. 70, 1356–1366 (2010).
Brantley, E. C. & Benveniste, E. N. Signal transducer and activator of transcription-3: a molecular hub for signaling pathways in gliomas. Mol. Cancer Res. 6, 675–684 (2008).
Wu, J., Keng, V. W., Patmore, D. M., Kendall, J. J., Patel, A. V., Jousma, E. et al. Insertional Mutagenesis Identifies a STAT3/Arid1b/beta-catenin Pathway Driving Neurofibroma Initiation. Cell Rep. 14, 1979–1990 (2016).
Wu, J., Liu, W., Williams, J. P. & Ratner, N. EGFR-Stat3 signalling in nerve glial cells modifies neurofibroma initiation. Oncogene 36, 1669–1677 (2017).
Wu, J., Patmore, D. M., Jousma, E., Eaves, D. W., Breving, K., Patel, A. V. et al. EGFR-STAT3 signaling promotes formation of malignant peripheral nerve sheath tumors. Oncogene 33, 173–180 (2014).
Reilley, M. J., McCoon, P., Cook, C., Lyne, P., Kurzrock, R., Kim, Y. et al. STAT3 antisense oligonucleotide AZD9150 in a subset of patients with heavily pretreated lymphoma: results of a phase 1b trial. J. Immunother. Cancer 6, 119 (2018).
Hirbe, A. C., Kaushal, M., Sharma, M. K., Dahiya, S., Pekmezci, M., Perry, A. et al. Clinical genomic profiling identifies TYK2 mutation and overexpression in patients with neurofibromatosis type 1-associated malignant peripheral nerve sheath tumors. Cancer 123, 1194–201. (2017).
Qin, W., Godec, A., Zhang, X., Zhu, C., Shao, J., Tao, Y. et al. TYK2 promotes malignant peripheral nerve sheath tumor progression through inhibition of cell death. Cancer Med. 8, 5232–5241 (2019).
Moslin, R., Gardner, D., Santella, J., Zhang, Y., Duncia, J. V. Liu, C. et al. Identification of imidazo[1,2-b]pyridazine TYK2 pseudokinase ligands as potent and selective allosteric inhibitors of TYK2 signalling. Medchemcomm 8, 700–712 (2017).
Papp, K., Gordon, K., Thaci, D., Morita, A., Gooderham, M. Foley, P. et al. Phase 2 trial of selective tyrosine kinase 2 inhibition in psoriasis. N. Engl. J. Med. 379, 1313–1321 (2018).
Le, L. Q., Shipman, T., Burns, D. K. & Parada, L. F. Cell of origin and microenvironment contribution for NF1-associated dermal neurofibromas. Cell Stem Cell 4, 453–463 (2009).
Toonen, J. A., Solga, A. C., Ma, Y. & Gutmann, D. H. Estrogen activation of microglia underlies the sexually dimorphic differences in Nf1 optic glioma-induced retinal pathology. J. Exp. Med. 214, 17–25 (2017).
Evans, D. G., O’Hara, C., Wilding, A., Ingham, S. L., Howard, E., Dawson, J. et al. Mortality in neurofibromatosis 1: in North West England: an assessment of actuarial survival in a region of the UK since 1989. Eur. J. Hum. Genet 19, 1187–1191 (2011).
Griffith, O. L., Spies, N. C., Anurag, M., Griffith, M., Luo, J., Tu, D. et al. Author Correction: The prognostic effects of somatic mutations in ER-positive breast cancer. Nat. Commun. 9, 4850 (2018).
Dischinger, P. S., Tovar, E. A., Essenburg, C. J., Madaj, Z. B., Gardner, E. E., Callaghan, M. E. et al. NF1 deficiency correlates with estrogen receptor signaling and diminished survival in breast cancer. NPJ Breast Cancer 4, 29 (2018).
Pearson A., Proszek P., Pascual J., Fribbens C., Shamsher M. K., Kingston B., et al. Inactivating NF1 mutations are enriched in advanced breast cancer and contribute to endocrine therapy resistance. Clin. Cancer Res. https://doi.org/10.1158/1078-0432.CCR-18-4044. (2019).
Zheng, Z. Y., Anurag, M., Lei, J. T., Cao, J., Singh, P., Peng J. et al. Neurofibromin is an estrogen receptor-alpha transcriptional co-repressor in breast cancer. Cancer Cell. 37, 387–402 (2020).
Liao, C. P., Pradhan, S., Chen, Z., Patel, A. J., Booker, R. C. & Le, L. Q. The role of nerve microenvironment for neurofibroma development. Oncotarget 7, 61500–61508 (2016).
Birchmeier, C. & Nave, K. A. Neuregulin-1. Neuregulin-1, a key axonal signal that drives Schwann cell growth and differentiation. Glia 56, 1491–1497 (2008).
Kazmi, S. J., Byer, S. J., Eckert, J. M., Turk, A. N., Huijbregts, R. P., Brossier, N. M. et al. Transgenic mice overexpressing neuregulin-1 model neurofibroma-malignant peripheral nerve sheath tumor progression and implicate specific chromosomal copy number variations in tumorigenesis. Am. J. Pathol. 182, 646–667 (2013).
Stonecypher, M. S., Byer, S. J., Grizzle, W. E. & Carroll, S. L. Activation of the neuregulin-1/ErbB signaling pathway promotes the proliferation of neoplastic Schwann cells in human malignant peripheral nerve sheath tumors. Oncogene 24, 5589–5605 (2005).
Simmons, G. W., Pong, W. W., Emnett, R. J., White, C. R., Gianino, S. M., Rodriguez, F. J. et al. Neurofibromatosis-1 heterozygosity increases microglia in a spatially and temporally restricted pattern relevant to mouse optic glioma formation and growth. J. Neuropathol. Exp. Neurol. 70, 51–62 (2011).
Liao, C. P., Booker, R. C., Brosseau, J. P., Chen, Z., Mo, J., Tchegnon, E. et al. Contributions of inflammation and tumor microenvironment to neurofibroma tumorigenesis. J. Clin. Invest. 128, 2848–2861 (2018).
Prada, C. E., Jousma, E., Rizvi, T. A., Wu, J., Dunn, R. S. Mayes, D. A. et al. Neurofibroma-associated macrophages play roles in tumor growth and response to pharmacological inhibition. Acta Neuropathol. 125, 159–168 (2013).
Fletcher, J. S., Springer, M. G., Choi, K., Jousma, E., Rizvi, T. A., Dombi, E. et al. STAT3 inhibition reduces macrophage number and tumor growth in neurofibroma. Oncogene. 38, 2876–2884 (2018).
Choi, K., Komurov, K., Fletcher, J. S., Jousma, E., Cancelas, J. A., Wu, J. et al. An inflammatory gene signature distinguishes neurofibroma Schwann cells and macrophages from cells in the normal peripheral nervous system. Sci. Rep. 7, 43315 (2017).
Park, S. J., Sawitzki, B., Kluwe, L., Mautner, V. F., Holtkamp, N. & Kurtz, A. Serum biomarkers for neurofibromatosis type 1 and early detection of malignant peripheral nerve-sheath tumors. BMC Med. 11, 109 (2013).
Jakacki, R. I., Cohen, K. J., Buxton, A., Krailo, M. D., Burger, P. C., Rosenblum, M. K. et al. Phase 2 study of concurrent radiotherapy and temozolomide followed by temozolomide and lomustine in the treatment of children with high-grade glioma: a report of the Children’s Oncology Group ACNS0423 study. Neuro. Oncol. 18, 1442–1450 (2016).
Paraskeva, C. & Williams, A. C. Are different events involved in the development of sporadic versus hereditary tumours? The possible importance of the microenvironment in hereditary cancer. Br. J. Cancer 61, 828–830 (1990).
Brosseau J. P., Le L. Q. Heterozygous tumor suppressor microenvironment in cancer development. Trends Cancer. 5, 541–546 (2019).
Brosseau, J. P., Liao, C. P., Wang, Y., Ramani, V., Vandergriff, T., Lee, M. et al. NF1 heterozygosity fosters de novo tumorigenesis but impairs malignant transformation. Nat. Commun. 9, 5014 (2018).
Liu, J., Gallo, R. M., Khan, M. A., Renukaradhya, G. J. & Brutkiewicz, R. R. Neurofibromin 1 impairs natural killer T-cell-dependent antitumor immunity against a T-Cell lymphoma. Front Immunol. 8, 1901 (2017).
Nanda, A. Autoimmune diseases associated with neurofibromatosis type 1. Pediatr. Dermatol. 25, 392–393 (2008).
Dodd, R. D., Lee, C. L., Overton, T., Huang, W., Eward, W. C. Luo, L. et al. NF1(+/−) hematopoietic cells accelerate malignant peripheral nerve sheath tumor development without altering chemotherapy response. Cancer Res. 77, 4486–4497 (2017).
Hirbe, A. C., Dahiya, S., Friedmann-Morvinski, D., Verma, I. M., Clapp, D. W. & Gutmann, D. H. Spatially- and temporally-controlled postnatal p53 knockdown cooperates with embryonic Schwann cell precursor Nf1 gene loss to promote malignant peripheral nerve sheath tumor formation. Oncotarget 7, 7403–7414 (2016).
Levin, A. A. Treating disease at the RNA Level with oligonucleotides. N. Engl. J. Med. 380, 57–70 (2019).
Mellert, K., Lechner, S., Ludeke, M., Lamla, M., Moller, P., Kemkemer, R. et al. Restoring functional neurofibromin by protein transduction. Sci. Rep. 8, 6171 (2018).
Bai, R. Y., Esposito, D., Tam, A. J., McCormick, F., Riggins, G. J. Wade Clapp, D. et al. Feasibility of using NF1-GRD and AAV for gene replacement therapy in NF1-associated tumors. Gene Ther. 26, 277–286 (2019).
Wallis, D., Li, K., Lui, H., Hu, K., Chen, M. J., Li, J. et al. Neurofibromin (NF1) genetic variant structure-function analyses using a full-length mouse cDNA. Hum. Mutat. 39, 816–821 (2018).
Zhu, Y., Ghosh, P., Charnay, P., Burns, D. K. & Parada, L. F. Neurofibromas in NF1: Schwann cell origin and role of tumor environment. Science 296, 920–922 (2002).
Wu, J., Williams, J. P., Rizvi, T. A., Kordich, J. J., Witte, D., Meijer, D. et al. Plexiform and dermal neurofibromas and pigmentation are caused by Nf1 loss in desert hedgehog-expressing cells. Cancer Cell. 13, 105–116 (2008).
Joseph, N. M., Mosher, J. T., Buchstaller, J., Snider, P., McKeever, P. E., Lim, M. et al. The loss of Nf1 transiently promotes self-renewal but not tumorigenesis by neural crest stem cells. Cancer Cell. 13, 129–140 (2008).
Radomska, K. J., Coulpier, F., Gresset, A., Schmitt, A., Debbiche, A. Lemoine, S. et al. Cellular origin, tumor progression, and pathogenic mechanisms of cutaneous neurofibromas revealed by mice with Nf1 knockout in boundary cap cells. Cancer Discov. 9, 130–147 (2019).
Isakson, S. H., Rizzardi, A. E., Coutts, A. W., Carlson, D. F., Kirstein, M. N., Fisher, J. et al. Genetically engineered minipigs model the major clinical features of human neurofibromatosis type 1. Commun. Biol. 1, 158 (2018).
White, K. A., Swier, V. J., Cain, J. T., Kohlmeyer, J. L., Meyerholz, D. K., Tanas, M. R. et al. A porcine model of neurofibromatosis type 1 that mimics the human disease. JCI Insight. 3, 1–18 (2018).
Cichowski, K., Shih, T. S., Schmitt, E., Santiago, S., Reilly, K., McLaughlin, M. E. et al. Mouse models of tumor development in neurofibromatosis type 1. Science 286, 2172–2176 (1999).
Vogel, K. S., Klesse, L. J., Velasco-Miguel, S., Meyers, K., Rushing, E. J. & Parada, L. F. Mouse tumor model for neurofibromatosis type 1. Science 286, 2176–2179 (1999).
King, D., Yang, G., Thompson, M. A. & Hiebert, S. W. Loss of neurofibromatosis-1 and p19(ARF) cooperate to induce a multiple tumor phenotype. Oncogene 21, 4978–4982 (2002).
Dodd, R. D., Mito, J. K., Eward, W. C., Chitalia, R., Sachdeva, M., Ma, Y. et al. NF1 deletion generates multiple subtypes of soft-tissue sarcoma that respond to MEK inhibition. Mol. Cancer Ther. 12, 1906–1917 (2013).
Kim, A., Stewart, D. R., Reilly, K. M., Viskochil, D., Miettinen, M. M. & Widemann, B. C. Malignant peripheral nerve sheath tumors state of the science: leveraging clinical and biological insights into effective therapies. Sarcoma 2017, 7429697 (2017).
Gutmann, D. H., McLellan, M. D., Hussain, I., Wallis, J. W., Fulton, L. L., Fulton, R. S. et al. Somatic neurofibromatosis type 1 (NF1) inactivation characterizes NF1-associated pilocytic astrocytoma. Genome Res. 23, 431–439 (2013).
Zhu, Y., Harada, T., Liu, L., Lush, M. E., Guignard, F. Harada, C. et al. Inactivation of NF1 in CNS causes increased glial progenitor proliferation and optic glioma formation. Development 132, 5577–5588 (2005).
Becher, O. J. & Holland, E. C. Evidence for and against regional differences in neural stem and progenitor cells of the CNS. Genes Dev. 24, 2233–2238 (2010).
Acknowledgements
We apologise to all colleagues whose relevant work we could not discuss due to space limitations.
Author information
Authors and Affiliations
Contributions
J.P.B. and C.P.L. drafted the paper and figures with scientific guidance from L.Q.L. J.P.B., C.P.L. and L.Q.L. revised and approved the final paper.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent to publish
Not applicable.
Data availability
Not applicable.
Competing interests
The authors declare no competing interests.
Funding information
L.Q.L. is supported by funding from the National Cancer Institute of the National Institutes of Health (grant number R01 CA166593 and U54 CA196519), the US Department of Defense, the Neurofibromatosis Therapeutic Acceleration Program, the NF1 Research Consortium Fund, and the Giorgio Foundation. J.P.B. and C.P.L. are recipient of the Young Investigator Award from the Children’s Tumor Foundation. J.P.B. is also an Early Investigator Research Award from the US Department of Defense. C.P.L. is also a recipient of the Career Development Award from Dermatology Foundation. L.Q.L. holds a Career Award for Medical Scientists from the Burroughs Wellcome Fund and the Thomas L. Shields, M.D. Professorship in Dermatology.
Additional information
Note This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution 4.0 International (CC BY 4.0).
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brosseau, JP., Liao, CP. & Le, L.Q. Translating current basic research into future therapies for neurofibromatosis type 1. Br J Cancer 123, 178–186 (2020). https://doi.org/10.1038/s41416-020-0903-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41416-020-0903-x
This article is cited by
-
Schwann cell derived pleiotrophin stimulates fibroblast for proliferation and excessive collagen deposition in plexiform neurofibroma
Cancer Gene Therapy (2024)
-
The therapeutic potential of neurofibromin signaling pathways and binding partners
Communications Biology (2023)
-
Neurofibromin and suppression of tumorigenesis: beyond the GAP
Oncogene (2022)
-
Human cutaneous neurofibroma matrisome revealed by single-cell RNA sequencing
Acta Neuropathologica Communications (2021)
-
Neurofibromatosis in the Era of Precision Medicine: Development of MEK Inhibitors and Recent Successes with Selumetinib
Current Oncology Reports (2021)
