
Abstract
Dopaminergic systems have been known to be involved in the regulation of locomotor activity and development of psychosis. However, the observations that some Parkinson’s disease patients can move effectively under appropriate conditions despite low dopamine levels (eg, kinesia paradoxia) and that several psychotic symptoms are typical antipsychotic resistant and atypical antipsychotic sensitive indicate that other systems beyond the dopaminergic system may also affect locomotor activity and psychosis. The present study showed that dopamine-deficient (DD) mice, which had received daily L-DOPA injections, could move effectively and even be hyperactive 72 h after the last L-DOPA injection when dopamine was almost completely depleted. Such hyperactivity was ameliorated by clozapine but not haloperidol or ziprasidone. Among multiple actions of clozapine, muscarinic acetylcholine (ACh) activation markedly reduced locomotor activity in DD mice. Furthermore, the expression of choline acetyltransferase, an ACh synthase, was reduced and extracellular ACh levels were significantly reduced in DD mice. These results suggest that the cholinergic system, in addition to the dopaminergic system, may be involved in motor control, including hyperactivity and psychosis. The present findings provide additional evidence that the cholinergic system may be targeted for the treatment of Parkinson’s disease and psychosis.
Similar content being viewed by others
Introduction
Dopamine (DA) plays an essential role in brain functions, including motor control, reward, and psychosis (Calabresi and Di Filippo, 2008; Ziauddeen and Murray, 2010). Low levels of DA and the blockade of DA neurotransmission generally cause hypolocomotion. The progressive degeneration of nigrostriatal DA neurons is well known to be the major pathological characteristic of Parkinson’s disease (PD). Medications for PD include the DA precursor L-dihydroxyphenylalanine (L-DOPA) that increases DA levels and enhances DA neurotransmission. In contrast, high levels of DA and enhanced DA neurotransmission generally cause hyperlocomotion that is considered to be relevant to psychosis. Some disorders, such as drug addiction and schizophrenia, have been reportedly associated with DA neuron activation in the mesocorticolimbic system (Lester et al, 2010). Almost all antipsychotic drugs block DA neurotransmission. However, several phenomena that are not consistent with this theory have been clinically reported. Effective movement in PD in certain situations (ie, kinesia paradoxia) indicates locomotor ability even at low DA levels. Typical antipsychotic drug-resistant and atypical antipsychotic drug-sensitive positive symptoms in schizophrenia indicate that at least a subgroup of positive symptoms is not treatable by the blockade of DA neurotransmission (Meltzer, 2013). To resolve this discrepancy, we investigated mice with extremely low levels of DA.
Dopamine-deficient (DD) mice were generated using a transgenic rescue approach, in which tyrosine hydroxylase (TH) expression in noradrenergic and adrenergic cells in mice that lacked TH expression was complemented by a specific DA β-hydroxylase gene promoter (Nishii et al, 1998). These mice exhibited a restoration of norepinephrine and epinephrine synthesis and prevention of the usual perinatal lethality and cardiac dysfunction observed in TH knockout mice. DD mice require daily administration of L-DOPA, the precursor of DA, to maintain feeding (Supplementary Figure S2; Szczypka et al, 1999; Zhou and Palmiter, 1995).
In the present study, we found hyperactivity in DD mice that could be ameliorated by the prototypical, atypical antipsychotic, clozapine.
Materials and Methods
Animals
DD mice were created as described previously (Nishii et al, 1998). Wild-type and DD mouse littermates from crosses of heterozygous/heterozygous DD mice on a C57BL/6J genetic background served as subjects. The experimental procedures and housing conditions were approved by the Institutional Animal Care and Use Committee (Animal Experimentation Ethics Committee of Tokyo Metropolitan Institute of Medical Science, Approval ID: 12–43), and all of the animals were cared for and treated humanely in accordance with our institutional animal experimentation guidelines. All of the mice were housed in an animal facility maintained at 23±1 °C and 55±5% relative humidity under a 12 h/12 h light/dark cycle with lights on at 0800 h and off at 2000 h. Food and water were available ad libitum. As routine maintenance of the mutants, 50 mg/kg L-DOPA was intraperitoneally (i.p.) administered to DD mice daily. Before this study, we confirmed that daily L-DOPA injections did not affect locomotor activity in wild-type mice (Supplementary Figure S1). Thus, wild-type mice were not treated with L-DOPA in the following experiments. In the experiments, male and female mice, 10 to 27 weeks old, were examined.
Surgery and Microdialysis Procedures
The mice were anesthetized with sodium pentobarbital (50 mg/kg, i.p.) and stereotaxically implanted with microdialysis probes in the striatum (anterior, +0.6 mm; lateral, +1.8 mm; ventral, −4.0 mm from bregma) (Franklin and Paxinos, 1997). At 24 h after implantation, the dialysis experiments were performed in freely moving animals. Ringer’s solution (145 mM NaCl, 3 mM KCl, 1.26 mM CaCl2, and 1 mM MgCl2, pH 6.5) was perfused at a constant flow rate of 1 μl/min. Perfusates were directly injected into the high-performance liquid chromatography system every 10 min using an autoinjector (EAS-20, Eicom, Kyoto, Japan). DA and serotonin (5-HT) in the dialysate were separated using a reverse-phase ODS column (PP-ODS, Eicom) and detected with a graphite electrode (HTEC-500, Eicom). The mobile phase consisted of 0.1 M phosphate buffer (pH 5.5) that contained sodium decanesulfonate (500 mg/l), ethylenediaminetetraacetic acid (EDTA; 50 mg/l), and 1% methanol. Norepinephrine (NE) in the dialysate was separated using a reverse-phase ODS column (CA-5ODS, Eicom) and detected with a graphite electrode (ECD-300, Eicom). The mobile phase consisted of 0.1 M phosphate buffer (pH 6.0) that contained sodium octanesulfonate (400 mg/l), EDTA (50 mg/l), and 5% methanol. Acetylcholine (ACh) in the dialysate was separated using an analytical column (Eicompak AC-GEL, Eicom) before being entered into an enzyme column (AC-Enzympak, Eicom) that contained immobilized Ach esterase and choline oxidase that converts ACh to hydrogen peroxide. Hydrogen peroxide was detected by a platinum electrode (HTEC-500, Eicom). The mobile phase consisted of 0.05 M potassium bicarbonate buffer that contained sodium decanesulfonate (300 mg/l) and EDTA (50 mg/l). Microdialysis samples were collected every 10 min for the DA, 5-HT, and NE assays or 20 min for the ACh assay. Usually, perfusion was initiated 180 min before the collection of baseline samples. The basal levels of extracellular DA (DAex), 5-HT (5-HTex), NE (NEex), and ACh (AChex) were obtained from average concentrations of three consecutive samples when they were stable. DAex, 5-HTex, and NEex responses to drugs are expressed as a percentage of basal levels. The areas under the curve (AUCs) of DAex, 5-HTex, and NEex during the 180 min period after drug administration were calculated as the effects of the drugs.
Locomotor Activity Measurements
Each mouse was exposed to an illuminated chamber (30 × 40 × 25 cm), and locomotor activity was measured with Supermex (Muromachi Kikai, Tokyo, Japan) and a sensor monitor mounted above the chamber. All counts were automatically summed and recorded every 10 min. After a 180-min habituation period, saline or drug was administered subcutaneously (s.c.), and locomotor activity was monitored continuously for 180 min. Locomotor tests with DD mice were performed 24 or 72 h after the last L-DOPA (50 mg/kg) injection. Each drug-treatment day was followed by at least a 2-week washout period, during which DD mice were treated only with L-DOPA (50 mg/kg), and wild-type mice remained untreated.
Catalepsy Test
The degree of catalepsy was measured by placing both forepaws on a horizontal bar, 4 cm above the tabletop. The time in seconds until the mouse took both forepaws off the bar was recorded, with a maximum cutoff time of 5 min.
Hindlimb Movement Recording
Before recording locomotion patterns, the mice were habituated to walk on a custom-made runway apparatus (length, 40 cm; width, 5 cm). The placement of all four paws was monitored by a mirror underneath the runway. The mirror was set a 45° angle from the vertical axis. To enable the observation of hindlimb movements, the fur on the hindlimb of each animal was shaved under isoflurane gas anesthesia (3% for induction, 1–2% for maintenance). Circular reflective markers (2 mm diameter) were precisely placed on the shaved skin of the right hindlimb at the iliac crest, greater trochanter (hip), knee, lateral malleolus (ankle), fifth metatarsophalangeal joint (MTP), and toe. After the mice were allowed to recover completely from anesthesia, they walked freely on the runway, and their locomotor movements were recorded at 200 frames/s using a high-speed digital image camera system (HAS-220, DITECT, Tokyo, Japan). The captured images were stored directly on a computer for later analysis.
Hindlimb Movement Analyses
Movement analyses were limited to the sagittal plane parallel to the direction of walking. For further details of analysis see Supplementary Information.
Whole-Genome Expression Analyses
The expression profiles of the genes related to ACh metabolism and signaling transduction pathways were analyzed using the Illumina iScan system with MouseRef-8 Expression BeadChips (Illumina, San Diego, CA, USA) that contain probes that detect over 24 000 transcripts. Total RNAs were isolated by Sepasol-RNA I super solution (Nacalai Tesque, Kyoto, Japan) from the striatum and globus pallidus in wild-type and DD mice. We used the area that includes the neostriatum, ventral striatum, and globus pallidus but not thalamus or hypothalamus in the present study. Total RNA (50 ng) was then applied to RNA amplification that was performed with the TargetAmp Nano-g Biotin-aRNA Labeling Kit (EPICENTRE Biotechnologies, Madison, WI, USA) according to the instructions of the manufacturer. Briefly, first-strand cDNA was synthesized with T7-oligo (dT) primer from total RNAs, and second-strand cDNA synthesis was then performed with synthesized first-strand cDNA. The in vitro transcription reaction was performed with the product of second-strand cDNA synthesis as the template for producing biotin-cRNAs by incorporating biotin-UTP into the RNA transcripts. Finally, 750 ng of biotin-cRNA was hybridized to the MouseRef-8 Expression BeadChips and then reacted with streptavidin-Cy3. The expression intensity of the transcripts on the BeadChips was detected with an Illumina iScan reader.
Mouse Brain Tissue Processing
Mouse brains were removed under deep anesthesia and divided sagittally. One hemisphere was snap-frozen in liquid nitrogen and stored at −80 °C for biochemical studies. The other hemisphere was fixed in Bouin’s solution and embedded in paraffin for the preparation of 4 μm serial sections for analyses.
Immunoblot Analyses
Mouse brains were homogenized in lysis buffer that contained 1% Triton X-100 (Fujita et al, 2010) and centrifuged at 100 000 g for 30 min. The supernatants (10 μg) were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and electroblotted onto nitrocellulose membranes. The membranes were blocked with 3% bovine serum albumin (BSA) in Tris-buffered saline (TBS) plus 0.2% Tween-20, followed by incubation with primary antibodies in TBS that contained 3% BSA. After washing, the membranes were further incubated with second antibody conjugated with horseradish peroxidase in TBS (1 : 10 000). Finally, the target proteins were visualized with ECL prime (GE Healthcare, Piscataway, NJ, USA), followed by quantification using MultiGauge software (Fuji Film, Tokyo, Japan).
Immunohistochemistry
Immunoblot analysis and immunohistochemistry were performed as previously described (Fujita et al, 2010). For further details of immunoblot analysis and immunohistochemistry see Supplementary Information.
Drugs
The following drugs were used: L-DOPA (50 mg/kg; Sigma-Aldrich, St Louis, MO, USA), methamphetamine (METH; 1 mg/kg; Dainippon Sumitomo Pharma, Osaka, Japan), haloperidol (1 mg/kg; Sigma-Aldrich), clozapine (5 and 10 mg/kg; Toronto Research Chemicals, Toronto, ON, Canada), 8-hydroxy-2-(di-n-propylamino)tetralin (8-OH-DPAT; 1 mg/kg; Sigma-Aldrich), ritanserin (3 mg/kg; Sigma-Aldrich), prazosin hydrochloride (1 mg/kg; Sigma-Aldrich), pyrilamine maleate salt (20 mg/kg; Sigma-Aldrich), clonidine (0.1 mg/kg; Sigma-Aldrich), oxotremorine-M (0.1 mg/kg; Sigma-Aldrich), donepezil hydrochloride (3 mg/kg, Tokyo Chemical Industry, Tokyo, Japan), and ziprasidone hydrochloride monohydrate (3 mg/kg; Toronto Research Chemicals). L-DOPA was prepared as 1.4 mg/ml in a 2.5 mg/ml ascorbic acid solution dissolved in saline. Methamphetamine, 8-OH-DPAT, pyrilamine, clonidine, and oxotremorine-M were dissolved in saline. Haloperidol and ritanserin were dissolved in 0.8% lactic acid. Clozapine was dissolved in a minimum amount of 0.1 N HCl and diluted to the required doses with saline. Prazosin and donepezil were dissolved in water. Ziprasidone was prepared as a suspension in aqueous Tween-80 (1% v/v in distilled water). L-DOPA was administered i.p. in a volume of 35 ml/kg, and the other drugs were administered s.c. in a volume of 10 ml/kg.
Statistical Analyses
The analyses were performed using two-way analysis of variance (ANOVA). Individual post hoc comparisons were performed using Fisher’s protected least significant difference (PLSD) test. Hindlimb movements, ACh level, and the expression of choline acetyltransferase (ChAT) were analyzed by Student’s t-test or Welch’s t-test. Values of P<0.05 were considered statistically significant. The data were analyzed using Statview J5.0 software (SAS Institute, Cary, NC, USA).
Results
Remaining DAex in DD Mice 24 h after the Last L-DOPA Injection
To confirm the depletion of DA in DD mice, we first measured DAex levels in the striatum using in vivo microdialysis. Baseline DAex levels in the striatum were detected in DD mice 24 h after the last intraperitoneal (i.p.) injection of L-DOPA (50 mg/kg), although they were significantly lower than the levels in wild-type mice (Table 1). A s.c. injection of the psychostimulant methamphetamine (METH; 1 mg/kg), which induces DA outflow from DA nerve terminals via the DA transporter, markedly increased DAex levels in both DD mice and wild-type mice (Figure 1a). Two-way ANOVA revealed a significant effect of drug (F1, 18=55.745, P<0.001) but not genotype (F1, 18=0.032, P=0.859) and no drug × genotype interaction (F1, 18=0.076, P=0.785). L-DOPA significantly increased DAex levels, with a 459±75% maximal increase in DD mice (Supplementary Figure S3a). The peak level of DAex was 5% of the level in wild-type mice after L-DOPA injection (Supplementary Figure S3b). L-DOPA did not apparently increase DAex levels in wild-type mice. These results suggest that DA systems may still partially function in DD mice 24 h after the last L-DOPA injection.

DAex, 5-HTex, and NEex in the striatum in DD mice. (a) METH-induced elevation of DAex levels in DD mice at 24 h after the last L-DOPA injection. ***P<0.001, compared with saline (SAL) group of the same genotype (two-way ANOVA followed by Fisher’s protected least significant difference (PLSD) post hoc test). (b) METH-induced elevation of DAex levels in DD mice at 72 h after the last L-DOPA injection. Baseline DAex could not be detected in DD mice. One-way ANOVA revealed a significant effect of drug (F1, 9=27.2431, P<0.001). ***P<0.001; NA, not applicable. (c) METH-induced elevation of 5-HTex levels in DD mice at 72 h after the last L-DOPA injection. ***P<0.001, compared with saline group within the same genotype (two-way ANOVA followed by Fisher’s PLSD post hoc test). (d) METH-induced elevation of NEex levels in DD mice at 72 h after the last L-DOPA injection. **P<0.01, compared with saline group within the same genotype (two-way ANOVA followed by Fisher’s PLSD post hoc test). The figure shows the time course of DAex, 5-HTex, or NEex before and after a subcutaneous injection of saline (SAL) or METH (1 mg/kg). The arrow indicates the drug injection time. Each point represents the mean±SEM of the percentage of baseline DAex, 5-HTex, or NEex. The histogram represents the mean AUC±SEM of DAex,5- HTex, or NEex during the 180 min period after the saline or METH injection (n=4–7).
Depletion of DAex and Altered 5-HTex Level in DD Mice at 72 h after the Last L-DOPA Injection
To deplete DA, L-DOPA was withdrawn for 72 h. DAex levels in DD mice 72 h after the last L-DOPA injection were below the detection limit for DA (ie, 0.07 fmol/sample with a signal-to-noise ratio of 2 under our measurement conditions; Figure 1b). Baseline 5-HTex levels were significantly higher in DD mice than in wild-type mice (Table 2), consistent with the increases in 5-HTex levels observed in mice whose DA neurons were neonatally lesioned with 6-hydroxydopamine (Avale et al, 2004). Baseline NEex levels were not different between wild-type and DD mice (Table 2). METH (1 mg/kg) markedly increased 5-HTex and NEex levels in wild-type and DD mice (Figure 1c and d). Two-way ANOVA (drug × genotype) of 5-HTex revealed a significant effect of drug (F1, 15=46.529, P<0.001) but not genotype (F1, 15=0.007, P=0.936) and no drug × genotype interaction (F1, 15=0.627, P=0.441). Two-way ANOVA (drug × genotype) of NEex revealed a significant effect of drug (F1, 17=22.154, P<0.001) but not genotype (F1, 17=1.423, P=0.249) and no drug × genotype interaction (F1, 17=0.248, P=0.625). Baseline and METH-induced increases in NEex levels were not altered in DD mice. These results suggest that the DA system may lose functionality in DD mice 72 h after the last L-DOPA injection.
Locomotor Activity in DD Mice
We next examined locomotor activity in DD mice 24 and 72 h after the last L-DOPA injection (Figure 2a). Previous studies reported that DD mice were hyperactive immediately after an L-DOPA injection and hypoactive after 24 h withdrawal (Szczypka et al, 1999; Zhou and Palmiter, 1995). In the present study, locomotor activity in DD mice 24 h after the last L-DOPA injection was lower than in wild-type mice during the initial 3 h but higher than in wild-type mice during the subsequent 3 h period (Figure 2a). Furthermore, DD mice at 72 h after the last L-DOPA injection were slow to initiate movement but gradually increased their locomotor activity for 30 min. The markedly increased locomotor activity was maintained throughout the following 5 h monitoring period (Figure 2a and Supplementary Movie S1). Wild-type mice exhibited a gradual reduction of locomotor activity during the 6 h period, suggesting habituation. Previous studies showed that DD mice displayed cataleptic behavior at 24 h after the last L-DOPA injection or in the juvenile stage without L-DOPA treatment (Nishii et al, 1998; Szczypka et al, 1999). In the present study, DD mice at 72 h after the last L-DOPA injection also displayed evident cataleptic behavior in the parallel bar test (Table 3 and Supplementary Movie S1). Although DD mice at 72 h after the last L-DOPA injection exhibited enhanced locomotor activity in the novel environment, they displayed a reduction of activity in the home cage (Supplementary Figure S4). In the juvenile stage, DD mice that were not treated with L-DOPA exhibited a reduction of spontaneous locomotor activity in the test cage (Nishii et al, 1998). Although locomotor activity in DD mice depended on experimental conditions, they first exhibited catalepsy and then hyperactivity in the novel environment.

Locomotor activity and gait features in DD mice. (a) Locomotor activity in DD mice at 24 or 72 h after the last L-DOPA injection (n=10–29). The arrow indicates the saline injection time. *P<0.05, **P<0.01, ***P<0.001, compared with wild-type mice (Student’s t-test). The data are expressed as mean±SEM. (b) Interjoint coordination patterns during the step cycle period in wild-type mice (n=8) and DD mice (n=9). The angle–angle plots illustrate the averaged data for coordination between joints of the hindlimb. (A) Knee vs hip. (B) Ankle vs knee. (C) Ankle vs hip. White star, foot contact at the beginning of the stance phase. Gray star, foot lift at the beginning of the swing phase. (c) Range of angular joint excursions during the step cycle period in wild-type mice (n=8) and DD mice (n=9). (A) Hip. (B) Knee. (C) Ankle. The data are expressed as mean±SEM. **P<0.01, ***P<0.001, compared with wild-type mice (Student’s t-test). (d, e) Temporal and spatial parameters of stride length (d) and walking speed (e) in wild-type mice (n=8) and DD mice (n=9) during locomotion. The data are expressed as mean±SEM. **P<0.01, ***P<0.001, compared with wild-type mice (Student’s t-test).
Hindlimb Movement in DD Mice at 72 h after the Last L-DOPA Injection
As DA is considered to play a key role in gait, we performed a kinematic analysis of hindlimb movements in DD mice at 72 h after the last L-DOPA injection. Compared with wild-type mice, DD mice exhibited marked hyperflexion of the knee and reduced movement magnitude at not only the knee but also the ankle joints during the step cycle period (Figure 2b and c and Supplementary Figure S5). Stride length and speed in DD mice were significantly shorter and lower, respectively, than in wild-type mice, and the step cycle was not different between wild-type mice and DD mice (Figure 2d and e and Supplementary Figure S6). No differences were observed in iliac crest heights and greater trochanter heights in the step cycle between DD mice and wild-type mice (Supplementary Figure S5). The scatterplot of stride length vs walking speed in wild-type and DD mice indicated that for any given speed, stride length was shorter in DD mice than in wild-type mice (Supplementary Figure S6). Altogether, DD mice exhibited a slight kinematic abnormality of hindlimb movements, indicating that gait was characterized by reduced joint movements, consistent with previous studies in subjects with PD (Morris et al, 2005). Gait disturbances in the advanced stage of PD include festination, start hesitation, and freezing of gait (Devos et al, 2010; Giladi, 2001). In PD patients, the spatial and temporal parameters of stable gait are characterized by a reduction of gait speed, stride length, and swing phase duration, together with an increase in cadence (Devos et al, 2010; Morris et al, 1994). Start hesitation, hyperactivity, and gait hypokinesia in DD mice at 72 h after the last L-DOPA injection resembled the key features of PD patients, although the phenotypes in DD mice were relatively mild compared with PD patients.
Stimulant and Antipsychotic Effects on Hyperactivity in DD Mice at 72 h after the Last L-DOPA Injection
To examine whether pharmacological treatment ameliorates abnormal behavior in DD mice, the psychostimulant METH, the typical antipsychotic drug haloperidol, and the prototypical, atypical antipsychotic drug clozapine were administered s.c. at 3 h after placing the mice in a novel environment. Methamphetamine (1 mg/kg) increased locomotor activity in wild-type mice but did not produce alterations in DD mice (Figure 3a). Haloperidol (1 mg/kg) and ziprasidone (3 mg/kg) markedly decreased locomotor activity in wild-type mice but not in DD mice (Figure 3b and Supplementary Figure S7). In contrast, clozapine (5 and 10 mg/kg) decreased locomotor activity in wild-type and DD mice (Figure 3c). Two-way ANOVA (genotype × drug) of locomotor activity revealed significant effects of genotype (F1, 72=92.543, P<0.001) and drug (F4, 72=59.581, P<0.001) and a genotype × drug interaction (F4, 72=16.598, P<0.001; Figure 3d). Methamphetamine and haloperidol did not affect hyperactivity in DD mice, suggesting that this hyperactivity was DA independent.

Stimulant and antipsychotic effects on hyperactivity in DD mice. (a–c) Locomotor responses to (a) METH (1 mg/kg) in wild-type mice (n=10) and DD mice (n=11), (b) haloperidol (1 mg/kg) in wild-type mice (n=9) and DD mice (n=8), and (c) clozapine (5 and 10 mg/kg) in wild-type mice (5 mg/kg, n=7; 10 mg/kg, n=5) and DD mice (5 mg/kg, n=6; 10 mg/kg, n=5). The arrows indicate the saline and drug injection time. (d) Locomotor activity after saline, METH, haloperidol, and clozapine administration. ***P<0.001, compared with saline treatment within the same genotype (two-way ANOVA followed by Fisher’s PLSD post hoc test). (e) Locomotor activity after 8-OH-DPAT, ritanserin, prazosin, clonidine, pyrilamine, oxotremorine-M, and donepezil administration. **P<0.01, ***P<0.001, compared with saline treatment within the same genotype (two-way ANOVA followed by Fisher’s PLSD post hoc test; n=4–11).
Clozapine has high affinity for the 5-HT2A, 5-HT2C, D4, muscarinic 1, and α1-adrenergic receptors but weak affinity for the D2 receptor (Arnt and Skarsfeldt, 1998). We examined which receptors are involved in the effects of clozapine in DD mice. The doses of the 5-HT1A receptor agonist 8-OH-DPAT (1 mg/kg), 5-HT2A/2C receptor antagonist ritanserin (3 mg/kg), α1 receptor antagonist prazosin (1 mg/kg), α2 receptor agonist clonidine (0.1 mg/kg), histamine H1 receptor antagonist pyrilamine (20 mg/kg), muscarinic receptor agonist oxotremorine-M (0.1 mg/kg), and cholinesterase inhibitor donepezil (3 mg/kg) have been shown to decrease locomotor activity in wild-type mice (Berendsen and Broekkamp, 1990; Heal and Philpot, 1987; Ninan and Kulkarni, 1998; Salomon et al, 2007). Locomotor activity was markedly decreased by oxotremorine-M and donepezil, whereas it was partially decreased by ritanserin, clonidine, and pyrilamine and not at all by 8-OH-DPAT or prazosin (Figure 3e). Two-way ANOVA (genotype × drug) of locomotor activity revealed significant effects of genotype (F1, 74=243.674, P<0.001) and drug (F6, 74=50.035, P<0.001) and a significant genotype × drug interaction (F6, 74=19.677, P<0.001). These results suggest that muscarinic receptor activation is mainly involved in the effects of clozapine on hyperactivity in DD mice.
AChex Levels, ChAT Gene Expression, and ChAT Protein Levels in DD Mice at 72 h after the Last L-DOPA Injection
To examine biochemical alterations in the cholinergic system in DD mice, we measured the gene expression of ACh synthase ChAT in the striatum and globus pallidus, cerebral cortex, and brainstem and found that the intensity of ChAT gene expression in the striatum and globus pallidus was significantly lower in DD mice than in wild-type mice at 72 h after the last L-DOPA injection (Figure 4a). Furthermore, baseline AChex levels in the striatum and ChAT protein expression in the striatum were significantly lower in DD mice than in wild-type mice at 72 h after the last L-DOPA injection (Figure 4b and c). ChAT-positive cells were reduced in DD mice compared with wild-type mice (Figure 4d).

Reduced AChex levels, ChAT gene expression, and ChAT protein levels in DD mice. (a) ChAT gene expression intensity in the area of the neostriatum, ventral striatum, and globus pallidus. The data are expressed as mean±SEM. *P<0.05, compared with wild-type mice (Student’s t-test). (b) Baseline AChex levels in the striatum in DD mice. The data are expressed as mean±SEM. *P<0.05, compared with wild-type mice (Student’s t-test). (c) Immunoblot analysis of ChAT protein expression in the striatum. Representative blots and data quantified against β-actin bands are shown as mean±SEM. *P<0.05, compared with wild-type mice (Student’s t-test). (d) Immunohistochemical analysis of ChAT. The two lower panels (C, D) are insets from the two upper panels (A, B). (A, C) Wild-type mice. (B, D) DD mice. Scale bars=200 μm (A, B) and 100 μm (C, D). Arrows indicate ChAT-positive cells.
Discussion
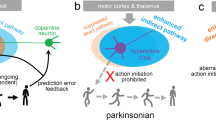
Locomotor activity and risk for psychosis were previously considered to decrease when DA levels decrease. In the present study, we found that animals with extremely low levels of DA could move effectively and were hyperactive (Figure 5). The hyperactivity may be relevant to kinesia paradoxia, a condition in which individuals with PD who typically experience severe difficulties with even simple movements may perform complex movements easily, although the mechanism is unknown (Au et al, 2010). Although DD mice are not a precise model of PD, they may partially reflect late-stage PD. Interestingly, hyperactivity in DD mice was ameliorated by clozapine. Because clozapine also suppressed locomotor activity in wild-type mice, the effect of the drug on motor activity may not be specific to DD mice. Remarkably, however, clozapine but not haloperidol or ziprasidone ameliorated hyperactivity in DD mice.

Illustrations showing possible relationships between locomotor activity and DA levels. Hyperlocomotion that occurs at high DA levels is ameliorated by antipsychotic drugs. Hypolocomotion that occurs at low DA levels is ameliorated by L-DOPA. Hyperlocomotion that occurs at extremely low DA levels is ameliorated by clozapine. Hyperlocomotion would be related to psychosis, and hypolocomotion would be related to PD.
Among the various effects of clozapine, 5-HT2A antagonism and 5-HT1A agonism are known to be critical elements of the antipsychotic and motor-sparing effects (Meltzer and Huang, 2008). In the present study, the 5-HT2A/2C receptor antagonist ritanserin partially ameliorated hyperactivity in DD mice. In light of the enhanced serotonergic activity in the DD mice reported herein (Tables 1 and 2), 5-HT2A inverse agonists or 5-HT1A partial agonists, both of which have shown promising results in treating behavioral and motor abnormalities in PD, should be further investigated in kinesia paradoxia (Huot et al, 2011; Meltzer et al, 2010).
Clozapine is well known to show improvements in schizophrenia patients with typical antipsychotic resistance. Several lines of evidence suggest that the effect of clozapine on muscarinic receptors may be important for the treatment of psychosis in patients with schizophrenia (Barak, 2009; Raedler et al, 2007, McKinzie and Bymaster, 2012). Hyperactivity that is induced by muscarinic receptor antagonists has been suggested to model antimuscarinic psychosis and cholinergic-related psychosis in schizophrenia (Yeomans, 1995). Similarly, hyperactivity in DD mice was ameliorated by a muscarinic receptor agonist and cholinesterase inhibitor, suggesting that such hyperactivity might be induced by similar mechanisms that underlie cholinergic-related psychosis in schizophrenia.
DD mice exhibited a decrease in ChAT expression, with a reduction of extracellular ACh levels in the striatum, demonstrating that the cholinergic system was suppressed in DD mice. DA depletion that is induced by α-methyl-p-tyrosine and reserpine was shown to decrease basal ACh release in the striatum (Bertorelli et al, 1992). In addition, ChAT activity has been reported to be normal or reduced in the striatum in PD patients (Hornykiewicz and Kish, 1987). Striatal cholinergic interneuron activity and ACh release were also reported to be modulated by DA neurons (Pisani et al, 2007). These previous results support the present finding that the ACh neurotransmission system is suppressed when the DA system becomes dysfunctional.
Dysfunctions in the interactions between DA and ACh may have important implications for various neurological and neuropsychiatric disorders (Lester et al, 2010). A reduction of dopaminergic input to the striatum causes relative cholinergic overactivity that in turn increases inhibitory output from the basal ganglia to thalamus, resulting in impaired motor function in PD (Graybiel, 2005). Motor function in PD patients is improved by anticholinergic drugs. In DD mice, hyperactivity was ameliorated by stimulation of the cholinergic system. Therefore, ACh may consistently reduce motor activity in PD patients and DD mice. Furthermore, the basal ganglia are interconnected with the pedunculopontine nucleus (PPN; Mena-Segovia et al, 2004). The PPN is thought to be involved in the initiation and modulation of gait and other stereotyped movements (Pahapill and Lozano, 2000). The loss of cholinergic neurons in the PPN has been reported in PD patients (Hirsch et al, 1987; Rinne et al, 2008). Cholinergic neurons in the PPN might also be altered because of low DA in DD mice as well as in PD patients.
DD mice lack DA in all brain regions, and determining the specific brain region responsible for hyperactivity may be difficult only by pharmacological approach. Injection of dopaminergic and cholinergic agents into specific brain regions, molecular and optogenetic approach, or viral rescue experiments may lead to a better understanding of the mechanisms that underlie this hyperactivity.
In conclusion, we investigated locomotion in mice with extremely low levels of DA and observed hyperactivity that is inconsistent in prior theories but consistent with kinesia paradoxia and typical antipsychotic drug-resistant symptoms in schizophrenia. We also pharmacologically found that reduced ACh levels may be involved in this hyperactivity.
FUNDING AND DISCLOSURE
The authors declare no conflict of interest. Over the past 3 years, KI has received support from Eisai, Taisho Pharmaceutical, and Nobelpharma for a project unrelated to this research and speaker’s fees from Shionogi, Taisho Pharmaceutical, and Japan Tobacco. Over the past 3 years, KK has received support from Novartis Pharma and Ono Pharmaceutical for a project unrelated to this research. Herbert Y Meltzer is a stockholder of ACADIA and SureGene. He has received grant support in the past 3 years from BioLine Rx, Cephalon, Dainippon Sumitomo, Eli Lilly, EnVivo, Janssen, Otsuka, Pfizer, and Sunovion. He is, or has been, a consultant to ACADIA, Alkemes, Astellas, Boehringer Mannheim, Bristol Myers Squibb, Cypress, Janssen, Lundbeck, Ovation, Merck, Novartis, Pfizer, Teva, and Valeant (BioVail).
References
Arnt J, Skarsfeldt T (1998). Do novel antipsychotics have similar pharmacological characteristics? A review of the evidence. Neuropsychopharmacology 18: 63–101.
Au WL, Lei N, Oishi MM, McKeown MJ (2010). L-dopa induces under-damped visually guided motor responses in Parkinson’s disease. Exp Brain Res 202: 553–559.
Avale ME, Nemirovsky SI, Raisman-Vozari R, Rubinstein M (2004). Elevated serotonin is involved in hyperactivity but not in the paradoxical effect of amphetamine in mice neonatally lesioned with 6-hydroxydopamine. J Neurosci Res 78: 289–296.
Barak S (2009). Modeling cholinergic aspects of schizophrenia: focus on the antimuscarinic syndrome. Behav Brain Res 204: 335–351.
Berendsen HH, Broekkamp CL (1990). Behavioural evidence for functional interactions between 5-HT-receptor subtypes in rats and mice. Br J Pharmacol 101: 667–673.
Bertorelli R, Zambelli M, Di Chiara G, Consolo S (1992). Dopamine depletion preferentially impairs D1- over D2-receptor regulation of striatal in vivo acetylcholine release. J Neurochem 59: 353–357.
Calabresi P, Di Filippo M (2008). ACh/dopamine crosstalk in motor control and reward: a crucial role for alpha 6-containing nicotinic receptors? Neuron 60: 4–7.
Devos D, Defebvre L, Bordet R (2010). Dopaminergic and non-dopaminergic pharmacological hypotheses for gait disorders in Parkinson’s disease. Fundam Clin Pharmacol 24: 407–4421.
Franklin KBJ, Paxinos G (1997) The Mouse Brain in Stereotaxic Coordinates. Academic Press: San Diego, CA, USA.
Fujita M, Sugama S, Sekiyama K, Sekigawa A, Tsukui T, Nakai M et al (2010). A beta-synuclein mutation linked to dementia produces neurodegeneration when expressed in mouse brain. Nat Commun 1: 110.
Giladi N (2001). Gait disturbances in advanced stages of Parkinson’s disease. Adv Neurol 86: 273–278.
Graybiel AM (2005). The basal ganglia: learning new tricks and loving it. Curr Opin Neurobiol 15: 638–644.
Heal DJ, Philpot J (1987). A study of the possible influence of central 5-HT function on clonidine-induced hypoactivity responses in mice. Psychopharmacology (Berl) 92: 219–223.
Hirsch EC, Graybiel AM, Duyckaerts C, Javoy-Agid F (1987). Neuronal loss in the pedunculopontine tegmental nucleus in Parkinson disease and in progressive supranuclear palsy. Proc Natl Acad Sci USA 84: 5976–5980.
Hornykiewicz O, Kish SJ (1987). Biochemical pathophysiology of Parkinson’s disease. Adv Neurol 45: 19–34.
Huot P, Fox SH, Newman-Tancredi A, Brotchie JM (2011). Anatomically selective serotonergic type 1A and serotonergic type 2A therapies for Parkinson’s disease: an approach to reducing dyskinesia without exacerbating parkinsonism? J Pharmacol Exp Ther 339: 2–8.
Lester DB, Rogers TD, Blaha CD (2010). Acetylcholine-dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci Ther 16: 137–162.
McKinzie DL, Bymaster FP (2012). Muscarinic mechanisms in psychotic disorders. Handb Exp Pharmacol 213: 233–265.
Meltzer HY (2013). Update on typical and atypical antipsychotic drugs. Annu Rev Med 64: 393–406.
Meltzer HY, Huang M (2008). In vivo actions of atypical antipsychotic drug on serotonergic and dopaminergic systems. Prog Brain Res 172: 177–197.
Meltzer HY, Mills R, Revell S, Williams H, Johnson A, Bahr D et al (2010). Pimavanserin, a serotonin(2A) receptor inverse agonist, for the treatment of Parkinson’s disease psychosis. Neuropsychopharmacology 35: 881–892.
Mena-Segovia J, Bolam JP, Magill PJ (2004). Pedunculopontine nucleus and basal ganglia: distant relatives or part of the same family? Trends Neurosci 27: 585–588.
Morris M, Iansek R, McGinley J, Matyas T, Huxham F (2005). Three-dimensional gait biomechanics in Parkinson’s disease: evidence for a centrally mediated amplitude regulation disorder. Mov Disord 20: 40–50.
Morris ME, Iansek R, Matyas TA, Summers JJ (1994). The pathogenesis of gait hypokinesia in Parkinson’s disease. Brain 117: 1169–1181.
Ninan I, Kulkarni SK (1998). 5-HT2A receptor antagonists block MK-801-induced stereotypy and hyperlocomotion. Eur J Pharmacol 358: 111–116.
Nishii K, Matsushita N, Sawada H, Sano H, Noda Y, Mamiya T et al (1998). Motor and learning dysfunction during postnatal development in mice defective in dopamine neuronal transmission. J Neurosci Res 54: 450–464.
Pahapill PA, Lozano AM (2000). The pedunculopontine nucleus and Parkinson’s disease. Brain 123: 1767–1783.
Pisani A, Bernardi G, Ding J, Surmeier DJ (2007). Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci 30: 545–553.
Raedler TJ, Bymaster FP, Tandon R, Copolov D, Dean B (2007). Towards a muscarinic hypothesis of schizophrenia. Mol Psychiatry 12: 232–246.
Rinne JO, Ma SY, Lee MS, Collan Y, Roytta M (2008). Loss of cholinergic neurons in the pedunculopontine nucleus in Parkinson’s disease is related to disability of the patients. Parkinsonism Relat Disord 14: 553–557.
Salomon L, Lanteri C, Godeheu G, Blanc G, Gingrich J, Tassin JP (2007). Paradoxical constitutive behavioral sensitization to amphetamine in mice lacking 5-HT2A receptors. Psychopharmacology (Berl) 194: 11–20.
Szczypka MS, Rainey MA, Kim DS, Alaynick WA, Marck BT, Matsumoto AM et al (1999). Feeding behavior in dopamine-deficient mice. Proc Natl Acad Sci USA 96: 12138–12143.
Yeomans JS (1995). Role of tegmental cholinergic neurons in dopaminergic activation, antimuscarinic psychosis and schizophrenia. Neuropsychopharmacology 12: 3–16.
Zhou QY, Palmiter RD (1995). Dopamine-deficient mice are severely hypoactive, adipsic, and aphagic. Cell 83: 1197–1209.
Ziauddeen H, Murray GK (2010). The relevance of reward pathways for schizophrenia. Curr Opin Psychiatry 23: 91–96.
Acknowledgements
We acknowledge Drs Minoru Saitoe and Tomoyuki Furuyashiki for critically reading the manuscript, Mr Michael Arends for assistance with editing the manuscript, and Ms Satomi Soma, Ms Junko Wakamatsu, Ms Yurie Nakamoto, and Ms Etsuko Kamegaya for assistance with breeding the DD mice. This work was supported by grants from the Ministry of Health, Labour and Welfare of Japan (H22-Iyaku-015 and H25-Iyaku-020), JSPS KAKENHI Grants 24659549, 24659490, and 24650205, MEXT KAKENHI Grant 25116532, the Smoking Research Foundation, the Naito Foundation, and the Astellas Foundation for Research on Metabolic Disorders. The funding agencies had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
YH, KK, HYM, and KI conceived and designed the experiments and wrote the paper. YH performed the microdialysis experiments and the behavioral experiments. SS, HY, and DY performed the kinematic analysis of hindlimb movements. SK performed the gene expression experiments. MF and MH performed immunoblot analysis and immunohistochemistry. KK provided the DD mice.
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article

Cite this article
Hagino, Y., Kasai, S., Fujita, M. et al. Involvement of Cholinergic System in Hyperactivity in Dopamine-Deficient Mice. Neuropsychopharmacol 40, 1141–1150 (2015). https://doi.org/10.1038/npp.2014.295
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2014.295
This article is cited by
-
Iron Supplementation and Exercise During Pregnancy: Effects on Behavior and the Dopaminergic System
Biological Trace Element Research (2023)
-
Involvement of muscarinic receptors in psychomotor hyperactivity in dopamine-deficient mice
Molecular Brain (2022)
-
Impaired wakefulness and rapid eye movement sleep in dopamine-deficient mice
Molecular Brain (2021)
-
Increase in excitability of hippocampal neurons during novelty-induced hyperlocomotion in dopamine-deficient mice
Molecular Brain (2020)
-
Hippocampal metabolism of amino acids by L-amino acid oxidase is involved in fear learning and memory
Scientific Reports (2018)
