
Abstract
In a biphasic, ovalbumin (OVA)-induced murine asthma model where allergic airway disease is followed by resolution and the development of local inhalational tolerance (LIT), transforming growth factor (TGF)-β-expressing CD5+ B cells were selectively expanded locally in hilar lymph nodes (HLN) of LIT mice. LIT HLN CD5+ B cells, but not LIT HLN CD5– B cells, induced expression of Foxp3 in CD4+CD25– T cells in vitro. These CD5+ regulatory B cells (Breg) and CD4+Foxp3+ T cells demonstrated similar increases in expression of chemokine receptors (CXCR4 and CXCR5) and co-localized in HLN B cell zones of LIT mice. The adoptive transfer of LIT HLN CD5+ B cells, but not LIT HLN CD5– B cells, increased the number of CD4+Foxp3+ T cells in the lung and inhibited airway eosinophilia in this OVA model. Thus, Breg in HLNs of LIT mice reside in a CD5+ TGF-β-producing subpopulation and co-localize with CD4+Foxp3+ T cells.
Similar content being viewed by others


Introduction
Research over the past three decades has provided overwhelming evidence that asthma is an inflammatory disease of the airways orchestrated by T lymphocytes.1, 2 Activation of T helper (Th) type 2 effector lymphocytes (Teff) causes elaboration of cytokines such as interleukin (IL)-4, IL-5, and IL-13, which in turn results in eosinophil activation and immunoglobulin (Ig)E production intrinsic to allergic inflammation.3, 4 The identification of regulatory T lymphocytes (Tregs) that can modulate Teff function has led to intensive research on their role in allergic asthma.5, 6, 7 B lymphocytes have traditionally been viewed as target cells for Th2 cytokines, with IL-4 and IL-13 responsible for the initiation and maintenance of B-cell IgE production.8 However, emerging evidence suggests that there are also regulatory types of B cells (Bregs), which are specifically induced under inflammatory conditions9, 10, 11, 12, 13, 14 and which are capable of suppressing inflammation, enhancing recovery, or inducing tolerance.15, 16, 17 Such Bregs have been implicated in the development of nasal tolerance to aeroallergens.18 Like Tregs, Bregs may comprise several subpopulations that exhibit different mechanisms of immunomodulation, including production of anti-inflammatory cytokines (IL-10 or TGF-β1), function as inhibitory antigen-presenting cells, and induction or recruitment of Tregs.19, 20
We have recently identified a novel putative Breg that may be involved in the regulation of airway inflammation.21 Acute aerosolized ovalbumin (OVA) exposure in sensitized mice induces typical allergic airway disease (AAD), but chronic inhalational antigen exposure results in resolution of the airway eosinophilia, Th2-cytokine secretion, and airway hyperreactivity22, 23, 24, 25, 26, 27 regardless of the persistence of OVA-specific IgE in serum and OVA-specific subcutaneous allergic reactions.23, 28 Reflecting the focal resolution of inflammation, we have termed this process local inhalational tolerance (LIT). Adoptive transfer of regional hilar lymph node (HLN) B cells from LIT mice to OVA-sensitized recipients increases the number of CD4+CD25+Foxp3+ Tregs in bronchoalveolar lavage (BAL) and attenuates AAD following subsequent OVA aerosol exposure.21 In contrast to AAD HLN B cells or LIT splenic B cells, LIT HLN B cells produce TGF-β and are capable of converting naive CD4+CD25– T cells into functionally suppressive CD4+CD25+Foxp3+ regulatory T cells (Foxp3+ Treg) in vitro.21 These findings discovered a regulatory role for regional B cells in the attenuation of inflammation and re-establishment of homeostasis in the airway despite ongoing antigen challenge. The present study was undertaken to further establish the phenotype of these TGF-β-producing Bregs and to investigate their relationship to Tregs with respect to location in the HLN and in chemokine receptor expression.
Results
CD5+ B cells were increased in the HLN at AAD and LIT
B cells isolated from inguinal lymph nodes and spleens were found to have an identical phenotype in both AAD and LIT mice: CD21intermIgDhi IgMintermCD5lo consistent with mature follicular B cells. AAD HLN B cells had a similar phenotype to the cells in these systemic tissues, whereas LIT HLN had increased percentage of IgDlo and CD5hi cells as compared with AAD (see Supplementary Figure S1 online). In HLN, there was a significant increase (P<0.005) in the total number of CD5+ B cells in mice at AAD and LIT as compared with mice in the Naive and Sensitized groups and to any of the groups in inguinal lymph nodes (Figure 1a). In the inguinal nodes, there were no significant differences in the total number of CD5+ B cells in mice at any of the stages (Figure 1a). Also in HLN, there was a significant increase (P<0.005) in the percentage CD5+ B cells in mice at LIT as compared with mice in the Naive, Sensitized, and AAD groups and to any of the groups in inguinal nodes (Figure 1b). By contrast, in the inguinal lymph nodes, there were no significant differences in the percentage of CD5+ B cells in mice at any of the stages (Figure 1b).

CD5+ B cells were increased in the hilar lymph node (HLN) at allergic airway disease (AAD) and local inhalational tolerance (LIT). Cells were isolated from hilar (black bars) and inguinal lymph nodes (ILN; grey bars) at different stages of the ovalbumin model (Naive, Sensitized (Sens), AAD, and LIT). Panel a depicts flow cytometry dot plots from representative tissues, and panels b and c show mean±s.e.m. values for total number (b) and percentage of CD5+ B cells (c). Numbers of CD5+ B cells were significantly expanded in hilar nodes at AAD and LIT, and they remained elevated as a percentage of total B cells during LIT. By contrast, no changes were seen in the inguinal nodes. n=8–12 in each group. **P<0.005 as compared with the naive and sensitized groups in hilar nodes and to all the groups in inguinal nodes (b), or P<0.005 as compared with all other groups (c).
LIT HLN CD5+ B cells predominantly expressed TGF-β
The selective expansion of CD5+ B cells in HLNs from LIT mice suggested that these cells could constitute the suppressive Breg phenotype identified previously, where B lymphocytes isolated from HLNs of LIT mice induced CD4+CD25+Foxp3+ T cells via expression of TGF-β.21 Thus, we examined whether the LIT HLN TGF-β-producing B cells resided in the CD5+ or CD5– cell population. Minimal anti-latency-associated protein (LAP) expression was seen in CD5+ or CD5– cells isolated from spleens of Naive or LIT mice (0.08–0.2% of these B cells were LAP+; Figure 2a and c). LAP was found at low levels in both types of B cells from HLN of Naive mice. Its expression increased in both CD5+ and CD5– B cells in HLN from LIT mice, but the enhanced expression was much more dramatic in the CD5+ LIT HLN B-cell population. In LIT HLNs, 37.9±9% of CD5+ B cells were LAP+, as compared with 13.3±4% of CD5– B cells (P<0.05; Figure 2c). As anticipated, little IL-10 staining was seen in unstimulated B cells, but IL-10 production could be induced by 5-h stimulation with phorbol 12-myristate 13-acetate and ionomycin.29 As shown in Figure 2b and d, IL-10 was also primarily produced by CD5+ B cells (8.2±1.3% in LIT HLN and 5.2±0.7% in LIT spleen) as compared with CD5− B cells (1.0±0.3% in LIT HLN and 0.6±0.1% in LIT spleen; P<0.0005 each). Unfortunately, the LAP staining was poor in these studies, possibly because of interference from the IL-10 staining conditions (e.g., permeabilization, low temperature), and so co-expression of IL-10 and TGF-β could not be directly assessed. However, in contrast to TGF-β, IL-10 production did not differ between LIT HLN and LIT spleen CD5+ B cells (P>0.05), and in the HLNs, CD5+ B-cell expression of IL-10 was significantly less than their expression of TGF-β (P<0.005; Figure 2d).

Anti-latency-associated protein (LAP) and interleukin (IL)-10 expression in CD5+ and CD5– B cells. Lymphocytes isolated from Spleens and hilar lymph node (HLN) of Naive and local inhalational tolerance (LIT) mice were stained for B220, CD5, anti-latency-associated protein (LAP), and IL-10, as described in the text. Panels a and b depict flow cytometry dot blots from representative tissues, and panels c and d show mean±s.e.m. values for LAP expression (c) and IL-10 expression (d) in CD5+ (black bars) and CD5– B cells (gray bars). (c) Relative to Naive animals, LAP expression was unchanged in spleen CD5+ and CD5– B cells of LIT mice, but LAP expression increased in HLN CD5+ and CD5– B cells at LIT. (d) IL-10 levels were increased in CD5+ B cells from both spleen and HLN compared with CD5– B cells, but there was no difference in expression between LIT spleen and LIT HLN. n=4–5 mice per group for LAP and 6 mice per group for IL-10 staining. **P<0.001 from other groups by ANOVA; †P<0.05 for CD5+ cells vs. CD5– cells.
LIT HLN CD5+ B cells induced formation of Foxp3+ T cells in vitro, and adoptive transfer of LIT HLN CD5+ B cells increased airway Foxp3+ Tregs in sensitized recipient mice
We have previously demonstrated that LIT HLN B cells can induce Foxp3 expression in naive T cells in vitro through a mechanism dependent on TGF-β and independent of IL-10.21 To determine whether the differential expression of LAP/TGF-β in CD5+ vs. CD5– HLN B cells resulted in functional distinctions in the two subsets, CD19+ B cells that were isolated from HLN of LIT mice were divided into CD5+ and CD5– populations, irradiated, and then co-cultured with naive splenic CD4+CD25– T cells and anti-CD3/CD28. Foxp3+ expression by the T cells was increased threefold by LIT HLN CD5+ B cells relative to control-stimulated conditions, but the LIT HLN CD5– B cells were without effect (Figure 3a). B cells were also required for the in vivo expansion of Foxp3+ Treg cells during LIT. In a first series of experiments, the progression of AAD to LIT was compared with wild-type mice and in B-cell–deficient JhD–/– mice (Jackson Laboratory, Bar Harbor, ME). Wild-type mice showed regional expansion of Foxp3+ T cells during LIT, occurring in hilar but not inguinal lymph nodes (Figure 3b). This regional expansion did not occur in the JhD–/– mice (Figure 3b; P<0.005 vs. wild-type mice). Next, adoptive transfer studies were performed with LIT HLN CD5+ B cells, LIT HLN CD5– B cells, and LIT Spleen CD5+ B cells. The number of airway Foxp3+ Tregs present in BAL of AAD mice increased by 43% in mice receiving LIT HLN CD5+ B cells (0.70±0.1×105 cells) as compared with saline control mice (0.5±0.1×105 cells) but was not affected by the adoptive transfer of LIT HLN CD5– B cells or LIT splenic CD5+ B cells (Figure 3c). Moreover, there was a direct correlation between the number of airway Foxp3+ Tregs and airway CD5+ B cells (r=0.56; P<0.005; Figure 3d). This correlation did not hold between airway Foxp3– Teff cells and CD5+ B cells in (r=0.27; P=0.20) or between airway Foxp3+ Tregs and CD5– B cells (r=0.31; P=0.14; data not shown).

In vitro and in vivo dependence of Foxp3+ Treg (regulatory T lymphocytes) cells on CD5+ B cells. (a) CD4+CD25– T cells from spleens of naive mice were co-cultured for 5 days without or with stimulation by soluble anti-CD3 and anti-CD28. Stimulated (Stim) cells were also co-cultured with either irradiated CD19+CD5+ B cells or CD19+CD5– B cells from local inhalational tolerance (LIT) hilar lymph nodes (HLNs). T-cell Foxp3 expression was increased when cultured with LIT HLN CD5+ B cells vs. with LIT CD5– B cells or anti-CD3/CD28 alone. Data represent mean±s.e.m. values of 3–4 animals per group; **P<0.01 vs. other groups by analysis of variance (ANOVA). (b) Foxp3+ Treg expression was compared in LIT control and JhD–/– mice. Control (CRL) mice showed expansion of Foxp3+ Treg cells in hilar nodes (black bars) but not inguinal nodes (striped bars), but this local expansion did not occur in the JhD–/– mice. n=5 mice per group; *P<0.005 between CRL and JhD–/– mice. (c) Two days before a week of daily ovalbumin (OVA) aerosol exposures, OVA-sensitized mice received tail vein injections of saline or specific B-cell populations. Mice receiving LIT HLN CD5+ B cells demonstrated increased numbers of Foxp3+ T cells in their bronchoalveolar lavage (BAL), as compared with control (saline) mice or mice receiving LIT HLN CD5– B cells or LIT spleen (SPL) CD5+ B cells. n=5–11 mice per group; *P<0.05 vs. other groups by ANOVA. (d) For all groups of mice in panel c, there was a direct correlation between the number of CD4+Foxp3+ Treg cells and the number of CD19+CD5+ B cells in BAL (dashed line; r=0.56; P<0.005). This relationship also held for the LIT HLN CD5+ recipient animals alone (solid line; r=0.73; P<0.05). By contrast, there was no association between CD4+CD25+Foxp3– Teff (Th2 effector lymphocytes) cells and CD19+CD5+ B cells in BAL (r=0.28; P>0.10; data not shown).
Foxp3+ Tregs increased in the B-cell zone and T cell–B cell border of HLN via confocal microscopy in mice at LIT
The expansion of airway and HLN Foxp3+ cells from AAD to LIT was associated with a striking difference in the distribution of Foxp3+ Tregs in HLN, as demonstrated in representative confocal microscopy images in Figure 4a–c. Qualitative observations of these confocal images showed apparent increases of Foxp3+ Tregs in HLN of mice at LIT (Figure 4a, lower panel) as compared with sensitized and AAD stages (Figure 4a, upper and middle panels). These observations were supported by quantification of Foxp3+ Tregs and B cells in different compartments of HLN, which included T-cell zone, T cell–B cell border (T–B border), and B-cell zone (follicle). Representative images for the quantification strategy of Foxp3+ Tregs in the T–B border are shown in Figure 4b and c. In general, the B-cell zones (follicular area) were reduced in size and number of cells in the HLN at LIT as compared with AAD, corresponding with the resolution of lung inflammation (see Supplementary Table 1 online). There was a 78.3% increase (P<0.05) in the density of Tregs (follicular Tregs) in the follicular B-cell zone of HLN from mice at LIT as compared with AAD (Figure 4d), with no changes in Foxp3+ Treg densities in the T-cell zone and T–B border. Moreover, in the T–B border and B-cell zone, the ratios of Foxp3+ Tregs to B cells increased 184 and 146%, respectively, in HLN from mice at LIT as compared with AAD (Figure 4e; P<0.05 each). Thus, Foxp3+ Tregs preferentially localized in the follicular area of HLN at LIT. CD5 staining was not bright enough to permit specific identification of CD5+ B cells by confocal microscopy.

Increased Foxp3+ Tregs (regulatory T lymphocytes) in the follicular area of the hilar lymph node (HLN) in mice at local inhalational tolerance (LIT) via confocal microscopy. Confocal microscopy was performed on HLNs from mice after sensitization (white bars), at allergic airway disease (AAD; grey bars), and at LIT (black bars) to determine the locations of B cells and CD4+Foxp3+Treg cells. (a–c) Representative confocal images of HLNs from sensitized (Sens; upper panels), AAD (middle panels), and LIT mice (lower panels), with quantification assessed using Imaris software as described in the Methods section. Regions of appropriate dimensions were selected along the T–B border for quantification of Foxp3+ Tregs in that area (b). Foxp3+ Tregs were enhanced using the spots tool in (c) Imaris Suite and counted. Representative colors for confocal microscopy: green B220+ cells, blue CD4+ cells, and red Foxp3+ cells. Images of magnification ×20. (d and e) Mean±s.e.m. values for (d) Foxp3+ Treg cell density and (e) the ratio of the number of Tregs to B cells in the different HLN zones. n=6–8 specimens per group; *P<0.05 as compared with sensitized and AAD in T–B border and B-cell zone.
The number of CD5+ B cells and Foxp3+ Tregs expressing CXCR4 and CXCR5 were increased in the HLN at AAD and LIT
The co-localization of B cells and Foxp3+ T cells in HLNs suggested that the cells could be expressing similar chemokine receptors. Therefore, the expression of CXCR4 and CXCR5 on CD5+ B cells and Tregs was compared between hilar and inguinal lymph nodes and at various stages of the OVA model (Naive, Sensitized, AAD, and LIT). Significantly more CD5+ B cells expressed CXCR4 in hilar nodes vs. inguinal nodes at each stage of the model (P<0.05; Figure 5b), and this regional expression increased further at AAD and LIT as compared with the Naive and Sensitized stages (P<0.05). Moreover, within HLN, expression of CXCR4 was 2.3–3.5-fold higher on CD5+ B cells than on CD5– B cells at all stages of the model (see Supplementary Figure S3 online). No changes were observed in the percentage of CXCR4+ CD5+ B cells in the inguinal lymph nodes except for a significant decrease at the Sensitized stage. There was no difference in the percentage of CD5+ B cells expressing CXCR5 at any of the stages in hilar or inguinal lymph nodes (Figure 5c). Like the percentages, the total number CD5+ B cells expressing CXCR4 was significantly increased in the HLN of mice at AAD and LIT as compared with mice at the Naive and Sensitized stages (see Supplementary Figure S2A online), and the total number of CD5+ B cells expressing CXCR5 was also increased at AAD and LIT (see Supplementary Figure S2B online). The number of CD5+ B cells expressing CXCR4 and CXCR5 was not altered in the inguinal lymph node at any of the stages (see Supplementary Figure S2A and B online).

Hilar lymph node (HLN) CD5+ B cell expression of CXCR4 and CXCR5. CD5+ B cells were isolated from hilar nodes (black bars) and inguinal nodes (ILN; gray bars) at different stages (Naive, Sensitized (Sens), allergic airway disease (AAD), and local inhalational tolerance (LIT)) of the ovalbumin model. Panel a depicts representative flow cytometry dot plots of CXCR4 expression on CD19+CD5+ B cells. Panel b demonstrates increased CXCR4+CD5+ B cells in hilar compared with inguinal lymph nodes at all stages of the model, and expansion of hilar node CXCR4+CD5+ B cells during AAD and LIT. Panel c demonstrates similar CXCR5 expression by CD5+ B cells in both tissues and at all stages of the model. Data represent the mean±s.e.m.; n=8–12 in each group (a and b); *P<0.05 as compared with the Naive and Sensitized groups in the HLN and to all the groups in the ILN; !P<0.05 as compared with all groups in the ILN; †P<0.05 between HLN and ILN in the Naive and Sensitized groups; ††P<0.005 between HLN and ILN in the AAD and LIT groups.
The percentage (Figure 6b) and total number (see Supplementary Figure S2C online) of Foxp3+ Tregs expressing CXCR4 were also increased in HLN in mice at AAD and LIT, as compared with the Naive and Sensitized groups (P<0.005), with no changes seen in inguinal nodes. In contrast to CD5+ B cells in HLN, the percentage of Foxp3+ Tregs expressing CXCR5 was significantly increased in mice at AAD and LIT as compared with the Naive and Sensitized stages (P<0.005; (Figure 6c). In inguinal nodes, the percentage of CXCR5+ Foxp3+ Tregs was not altered at any of the stages (Figure 6c). Similarly, the total number of Tregs expressing CXCR5 was significantly increased in HLNs of mice at AAD and LIT as compared with mice in the Naive and Sensitized stages (see Supplementary Figure S2D online). These data show that both CD5+ B cells and Foxp3+ Tregs up-regulate the expression of CXCR4 and CXCR5 at AAD and LIT specifically in hilar, but not inguinal, lymph nodes.

Hilar lymph node (HLN) Foxp3+ T cell expression of CXCR4 and CXCR5. Foxp3+ T cells were isolated from hilar nodes (black bars) and inguinal nodes (ILN; gray bars) at different stages (Naive, Sensitized (Sens), allergic airway disease (AAD), and local inhalational tolerance (LIT)) of the ovalbumin model. Panel a depicts representative flow cytometry dot plots of CXCR4 and CXCR5 expression on CD4+Foxp3+ T cells. Panel b demonstrates increased CXCR4+Foxp3+ T cells in hilar compared with inguinal lymph nodes at AAD and LIT, and expansion of hilar node CXCR4+Foxp3+ T cells during AAD and LIT. Panel c demonstrates expansion of hilar node CXCR4+Foxp3+ B cells during AAD and LIT. Data represent the mean±s.e.m.; n=4–8 in each group (a and b); **P<0.005 as compared with the Naive and Sensitized groups in the HLN; ††P<0.005 between HLN and ILN in the AAD and LIT groups.
Adoptive transfer of LIT HLN CD5+ B cells attenuated airway eosinophilia in sensitized recipient mice
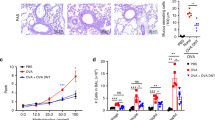
The association between CD5+ B cells and Foxp3+ Tregs suggested that the regional HLN B cells responsible for the previously reported attenuation of AAD21 resided in the CD5+ population. To test this, the degree of AAD was compared in animals receiving LIT HLN CD5+ B cells and animals receiving other cells. The development of airway eosinophilia was significantly reduced in mice that received LIT HLN CD5+ B cells (60.6±2.0% and 17.2±2.6×105 eosinophils) as compared with mice that received either vehicle alone, LIT HLN CD5– B cells, or LIT spleen CD5+ B cells (Figure 7). Mice receiving LIT spleen CD5– B cells, naive spleen CD5– B cells, or naive spleen CD5+ B cells also developed similar airway eosinophilia as the control recipient animals (data not shown).

Attenuation of allergic airway disease (AAD) by adoptive transfer of local inhalational tolerance (LIT) hilar lymph node (HLN) CD5+ B cells into sensitized recipient mice. Ovalbumin (OVA)-sensitized mice received tail vein injections of saline or specific B-cell populations (0.2×106 CD19+CD5+ or 1.0×106 CD19+CD5– cells) 2 days before a week of daily exposure to 1% OVA aerosols. Mice receiving the LIT HLN CD5+ B cells (solid bars) developed attenuated AAD, with less (a) relative and (b) absolute airway eosinophilia relative to control (saline) mice or mice receiving LIT HLN CD5– B cells or LIT spleen CD5+ B cells. Data represent mean±s.e.m. values of 5–11 mice per group; *P<0.05 vs. the other groups by analysis of variance. BAL, bronchoalveolar lavage.
Discussion
Previously, we have shown that regional B cells isolated from HLN of LIT mice are capable of suppressing AAD in recipient animals.21 B cells from LIT HLNs are unique in this function, as AAD suppression is not exerted by B cells isolated from AAD HLN or by splenic B cells of either AAD or LIT mice.21 These LIT HLN Bregs induce Foxp3+ T-cell generation through expression of TGF-β.21 The present study further characterized these Bregs as a specific local CD19+B220+CD1dhiCD5+ B-cell population expressing TGF-β and up-regulated the expression of the chemokine receptor CXCR4.
The finding of CD5+ cells in LIT HLNs was distinctive, as there are very few reports of CD5+ cells in draining lymph nodes.30 Their presence, in part, could be attributed to their expression of the chemokine receptors CXCR4 and CXCR5. The high level of CXCR5 expression by all the hilar and inguinal node B cells was consistent with its constitutive expression on circulating B cells, which directs their homing to B-cell follicles.31, 32 The up-regulation of CXCR4 on HLN CD5+ B cells at AAD and LIT would direct their localization to germinal center dark zones, which are located adjacent to T-cell areas.33 Further, the increased expression of CXCR4 and CXCR5 on HLN Foxp3+ Tregs during AAD and LIT would facilitate their interaction with follicular B cells by placing Bregs and Foxp3+ Tregs in close proximity in the hilar nodes. Foxp3+ Treg cells are known to express CXCR4,34 and so the increased numbers of CXCR4+Foxp3+ Treg cells in HLNs of AAD and LIT mice may have been due to enhanced migration of existing Foxp3+ Treg cells to the HLN. Alternately, the ability of LIT HLN CD5+ B cells to convert naive CD4+ T cells to suppressive Foxp3+ Treg cells in vitro raises the intriguing possibility that this conversion may be occurring in vivo in the HLN follicles, initially during AAD to help limit the inflammatory response and continued at LIT to sustain the re-established airway homeostasis. The confocal images support the close localization of these regulatory cells in HLN especially in the T–B border, both qualitatively and quantitatively.
Other investigators have implicated a regulatory role for B-1a CD19+CD5+ cells in moderating allergic airway disease. In a cockroach allergen-induced asthma model, mice that are deficient in B-1a cells (CBA/CaHN-Btkxid/J mice) develop higher serum IgE levels and more airway inflammation than wild-type mice.35 It should be noted, however, that Xid mice have several B-cell abnormalities, including impaired survival of follicular IgDhiIgMlo B cells and reduced numbers of marginal zone B cells.36 Thus, potential abnormalities in other Breg populations in Xid mice could have accounted for their increased allergic airway disease. The IgDloCD5hi phenotype also suggested a B-1 rather than B-2 cell morphology in our subpopulation of LIT HLN B cells; however, all the cells were also B220+ (e.g., CD45Rhi), which is associated with B-2 cells. Thus, the expanded population of CD19+CD5+ cells in LIT HLNs had characteristics of both B-1 (IgDlo) and B-2 (B220+) subsets, and therefore may reflect a previously undefined Breg-cell population that arose with chronic antigen exposure. Indeed, while CD5 expression is characteristic of B-1 cells, this surface marker can also be induced in B-2 cells by a variety of stimuli.37 CD5 expression can be induced in human peripheral blood B cells by co-cultivation with T cells via CD40-CD40L interaction, and these CD5+ B cells have T-cell suppressive activity.38 As our LIT mice are chronically exposed to OVA for 6 weeks, the CD5+ cells seen in their HLN may represent induced expression of the marker by non-B-1 cells.
To date, most studies of Breg have focused on IL-10-producing B10 cells that reside in a CD19hiCD1dhiCD5+ B-cell population.39, 40 It has been shown that B10 cells expand in response to Schistosoma mansoni infections, and that the adoptive transfer of these B10 cells into allergen-sensitized mice suppresses anaphylaxis and AAD through IL-10-dependent mechanisms.41, 42 These B10 cells have a CD19+CD21hiCD23+IgD+IgMhiCD1dhi phenotype,42 somewhat similar to our CD19+CD21intermIgDloIgMintermCD1dhi LIT HLN B cells. In the worm model, these suppressive B10 cells were found in the spleen. We found some expansion of CD5+ IL-10-producing B cells in the spleen and HLN in LIT mice (Figure 2d); however, these cells were ineffective at inhibiting AAD (Figure 7). In our LIT model, the suppressive Bregs arose only in regional HLNs, and no suppressive B cells were found in spleen or inguinal lymph node tissues. Moreover, the LIT hilar nodes contained fivefold more TGF-β+ cells than IL-10+ cells (Figure 2). Of interest, CD19+ B cells from regional, mesenteric lymph nodes of mice with chronic enteric helminth infections are also capable of suppressing AAD through an IL-10-independent mechanism.43 In addition, both IL-10-producing CD19+CD5+ B cells (B-10 cells) and TGF-β-producing CD19+CD5+ B cells have been shown to regulate cow's milk allergic responses in human subjects.44, 45 TGF-β-expressing B cells have also been shown to down-regulate pathogenic Th1 immunity in non-obese diabetic mice.14 As with T cells, it is likely that several populations of Breg exist, and that different subsets may be preferentially generated based on the types of stimuli and anatomical sites of antigen presentation.
Asthma is a complex disease. Its generation and resolution, pathogenesis and regulation are dependent upon intricate interactions between adaptive and innate immune cells. In animal models, antigen-induced AAD constitutes an unwanted and unnecessary Th2-driven inflammatory reaction against harmless inhaled antigens. LIT represents one process by which mice are able to suppress this detrimental inflammatory response and re-establish airway homeostasis despite ongoing systemic responses. It is apparent, both from basic science and clinical stand points, that no one cell type—effector or regulator—stands alone in this disease and is able to create or prevent asthma. Our studies have shown that LIT does not depend on the presence of either Bregs or Foxp3+ Tregs in knockout or antibody-depleted animals.21, 46 Although B-cell knockout animals develop LIT, they do so despite diminished levels of regional Foxp3+ Tregs. This finding supports the complex and interactive contributions of Bregs and Tregs in the development of tolerance in wild-type animals. Indeed, the contrasting pro- and anti-inflammatory roles of B and T cells are likely to overlap in asthma, with the balance of these opposing actions determining the natural course of the disease.
In summary, the present study identified a subpopulation of B cells that arose preferentially in regional lymphoid tissues in response to chronic antigen exposure and inflammation. These cells express a CD19+B220+CD21intermIgDloIgMinterm CD1dhiCD5+ phenotype, produce TGF-β, induce formation of regulatory Foxp3+ T cells in vitro, and suppress development of AAD in vivo via enhanced accumulation of Foxp3+ T cells in the airways. Further analysis of the genesis and function of these TGF-β-producing Bregs will help us understand how mice are capable of modulating their AAD and, thereby, yield potential insights into a cure for asthma.
Methods
OVA animal model protocol. Female C57BL/6J mice were obtained from The Jackson Laboratory (Bar Harbor, ME). Animals were housed in the animal facility at the University of Connecticut Health Center and were treated in accordance with the institutional and Office of Laboratory Animal Welfare guidelines. As previously described,21, 22, 23 mice were immunized with 3 weekly intraperitoneal injections of 25 μg OVA in alum. One week after the last immunization, the mice are exposed to 1% aerosolized OVA in physiological saline, 1 hour/day for either 7 days (AAD) or 42 days (LIT), with an estimated inhaled daily dose of 30–40 μg per mouse.
BAL/tissue analysis. At sacrifice, BAL fluid, HLN, inguinal lymph nodes, and spleens were harvested and processed for the isolation and enumeration of leukocytes. For collection of BAL, lungs were lavaged in situ with five 1.0 ml aliquots of sterile saline. Lymph nodes and spleens were harvested and mechanically disrupted into single-cell suspensions, with lysis of splenic erythrocytes by Tris-buffered ammonium chloride solution (9 parts 0.83% w/v NH4Cl; 1 part 2.57% w/v Tris, pH 7.0). For all tissue samples, total nucleated cell counts were obtained using a hemocytometer with nigrosin dye exclusion as a measure of viability.
Antibodies and immunofluorescence reagents. The following monoclonal antibodies, either phycoerythrin or fluorescein isothiocyanate conjugated or unlabeled, were purchased from BD Pharmingen (San Diego, CA) and eBiosciences (San Diego, CA): CD3 (145-2C11), CD4 (RM4-5), CD5 (53-7.3), CD8 (53-6.72), CD19 (1D3), CD25 (PC61), B220 (RA3-6B2), CXCR4 (2B11), CXCR5 (2G8), Foxp3 (FJR-16s), IL-10 (JES5-16E3), IgD (11–26), and IgM (11/41). In addition, phycoerythrin-labeled anti-CD4 and anti-CD8 were purchased from Coulter Immunology (Marietta, GA). A LAP of TGF-β was purchased from R & D Systems (Minneapolis, MN) and used to detect TGF-β bound to cells.
FACS analysis. Isolated lymphocytes were resuspended in phosphate-buffered saline (PBS) containing 0.2% bovine serum albumin and 0.1% NaN3 at a concentration of 1×105 to 1×106 white blood cells per ml, and 100 μl of the cells were incubated with 100 μl of properly diluted mAb at 4°C for 30 min. For the identification of Foxp3+ Treg cells, cells stained with anti-CD3, anti-CD4, and anti-CD25 were permeabilized using fixation/permeabilization buffer, following the manufacturer's protocol, and stained using anti-Foxp3-allophycocyanin (FJK-16s) with corresponding isotype controls (eBioscience, San Diego, CA). After staining, the cells were washed twice with PBS containing 0.2% bovine serum albumin and 0.1% NaN3 solution, and relative fluorescence intensities were measured on a 4-decade log scale by flow-cytometric analysis using a FACSCalibur or LSR II (Becton-Dickinson, San Diego, CA). For identification of CXCR4- and CXCR5-positive cells, cells were stained, incubated at 37°C for 45 min, washed in the above PBS solution, and samples were run on Becton Dickinson LSR II.
For intracellular IL-10 staining, 1×106 cells from the Spleen or HLN were plated in a 96-well plate and treated with BD Golgi Plug (BD Pharmigen, San Diego, CA). Cells were stimulated with phorbol 12-myristate 13-acetate (50 ng/ml; Sigma-Aldrich, St Louis, MO) plus ionomycin (1 μg/ml; Sigma-Aldrich) or left unstimulated for 5 h at 37°C.29 Cells were then stained for surface cell markers, treated with Cytofix/perm buffer (BD Pharmigen), and followed by intracellular cytokine staining with α-IL-10 at 4°C After staining, all samples were run on LSR II and analyzed with FlowJo (Tree Star Software, Ashland, OR).
Mice at the various stages were killed on different days and hence the gating strategy was kept consistent for all tissues at a particular stage. This is shown by different sized quadrants in the dot plots in Figures 1, 5 and 6. Selective representative dot plots are presented in each figure.
Confocal microscopy. HLN were cryopreserved in optimal cutting temperature, sectioned, and stained as described previously.47 In brief, frozen sections of thickness 20 μm were fixed in acetone or glutaraldehyde, washed in ice-cold PBS, and stained for 60 min at room temperature with a combination of fluorescently conjugated antibodies (anti-B220, anti-CD4, anti-Foxp3) in 2% normal goat serum and 2% FCS (fetal calf serum)/PBS solution. For consistency, the middle sections from each node were used for staining. To detect Foxp3 positive cells, the sections were prepared and permeabilized according to instructions in the Foxp3 staining kit (eBiosciences, CA). The sections were then Fc blocked (anti CD16/CD32), stained with Foxp3 overnight at 4°C, washed, mounted, and analyzed by confocal microscopy.
For quantification of B220+ B cells and Foxp3+ Tregs, the spots tool in Imaris suite was used. B-cell zones were selected using boxes along the entire area of B220+-cell clusters and T-cell zones were selected using boxes along the entire area based on the distribution of CD4+ cells. The T–B border of 100 μm thickness (50 μm into the B-cell zone and 50 μm into the T–cell zone) was selected using approximately four boxes of appropriate length and width along the border of B-cell zone. The four boxes represent the T–B cell border for one follicular image. For every mouse, 3–4 such follicular areas were imaged, and there were 2 mice per group. B220+ cells (green) and Foxp3+ cells (red) were enhanced, and the spots were counted to obtain the total number of each cell population. The ratios were then calculated from these numbers. To calculate the density values, X, Y, and Z dimensions were obtained from regions selected based on the distribution of T cells and B cells. The density was calculated using the formula=number of Tregs in a specific compartment/(X×Y×Z) of that compartment, with units=μm−3.
Treg-cell induction assay. Freshly isolated CD4+CD25– Teff cells were obtained from the spleens of Naive mice using magnetic bead isolation (Miltenyi Biotech, Auburn, CA), with >95% purity. These Teff cells (1×106) were cocultured for 5 days with viable irradiated (2,600 Rad) CD19+ CD5+ or CD5– B cells from HLNs of LIT mice (1×106; isolated as above), soluble anti-CD3 (1 μg/ml), and anti-CD28 (1 μg/ml). The cultures were in RPMI 1640 supplemented with 10% FCS and 50 μM 2-ME in 24-well plates. After 5 days in culture, the cells were taken out from the wells, and the intracellular expression of Foxp3 was assessed on gated CD4+ T cells. All studies were performed in triplicate.
B-cell isolation and adoptive transfers. B cells from HLNs and spleens of LIT mice were positively selected using a mouse CD19+ isolation kit (Miltenyi Biotech). The CD19+ cells were then separated by immunofluorescent labeling and subsequent isolation utilizing a FACS-Vantage sorter into CD5– and CD5+ subpopulations. Purified B-cell subpopulations (0.2×106 CD19+CD5+ or 1.0×106 CD19+CD5– cells) were injected by tail vein into sensitized animals 2 days before 7 days of OVA aerosol exposures. Different numbers of CD19+CD5+ and CD19+CD5– cells were chosen for transfer to represent their relative proportions in the whole lymph node adoptive transfer studies.21
Statistical analysis. Data were expressed as mean±standard error of mean values. Statistical comparisons between group means were made with analysis of variance followed by Fisher's protected least significant difference test between possible pair-wise combinations of means (StatView 4.5; Abacus Concepts, Berkeley, CA or JMP7; SAS Institute, Cary, NC). In all comparisons, P<0.05 was used to determine statistical significance.
References
Mosmann, T.R. & Coffman, R.L. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 7, 145–173 (1989).
Ricci, M., Ross, O., Bertoni, M. & Matucci, A. The importance of Th2-like cells in the pathogenesis of airway allergic inflammation. Clin. Exp. Allergy 23, 360–369 (1993).
Till, S. et al. IL-13 production by allergen-stimulated T cells is increased in allergic disease and associated with IL-5 but not IFN-gamma expression. Immunology 91, 53–57 (1997).
Robinson, D.S. et al. Predominant TH2-like bronchoalveolar T-lymphocyte population in atopic asthma. N. Engl. J. Med. 326, 298–304 (1992).
Romagnani, S. Immunologic influences on allergy and the TH1/TH2 balance. J. Allergy Clin. Immunol. 113, 395–400 (2004).
Xystrakis, E., Boswell, S.E. & Hawrylowicz, C.M. T regulatory cells and the control of allergic disease. Exp. Opin. Biol. Ther. 6, 121–133 (2006).
Larché, M. Regulatory T cells in allergy and asthma. Chest 132, 1007–1014 (2007).
Poulsen, L.K. & Hummelshoj, L. Triggers of IgE class switching and allergy development. Ann. Med. 39, 440–456 (2007).
Wolf, S.D., Dittel, B.N., Hardardottir, F. & Janeway, C.A. Experimental autoimmune encephalomyelitis induction in genetically B cell-deficient mice. J. Exp. Med. 184, 2271–2278 (1996).
Fillatreau, S., Sweenie, C.H., McGeachy, M.J., Gray, D. & Anderton, S.M. B cells regulate autoimmunity by provision of IL-10. Nat. Immunol. 3, 944–950 (2002).
Mizoguchi, A., Mizoguchi, E., Takedatsu, H., Blumberg, R.S. & Bhan, A.K. Chronic intestinal inflammatory condition generates IL-10-producing regulatory B cell subset characterized by CD1d up-regulation. Immunity 16, 219–230 (2002).
Wei, B. et al. Mesenteric B cells centrally inhibit CD4+ T cell colitis through interaction with regulatory T cell subsets. Proc. Natl. Acad. Sci. USA 102, 2010–2015 (2005).
Lenert, P., Brummel, R., Field, H. & Ashman, R.F. TLR-9 activation of marginal zone B cells in lupus mice regulates immunity through increased IL-10 production. J. Clin. Immunol. 25, 29–40 (2005).
Tian, J., Zekzer, D., Hanssen, L., Lu, Y., Olcott, A. & Kaufman, D.L. Lipopolysaccharide-activated B cells down-regulate Th1 immunity and prevent autoimmune diabetes in nonobese diabetic mice. J. Immunol. 167, 1081–1089 (2001).
Gonnella, P.A., Waldner, H.P. & Weiner, H.L. B cell-deficient (μMT) mice have alterations in the cytokine microenvironment of the gut-associated lymphoid tissue (GALT) and a defect in the low dose mechanism of oral tolerance. J. Immunol. 166, 4456–4464 (2001).
Parekh, V.V., Prasad, D.V., Banerjee, P.P., Joshi, B.N., Kumar, A. & Mishra, G.C. B cells activated by lipopolysaccharide, but not by anti-Ig and anti-CD40 antibody, induce anergy in CD8+ T cells: role of TGF-β1. J. Immunol. 170, 5897–5911 (2003).
D'Orazio, T.J., Mayhew, E. & Niederkorn, J.Y. Ocular immune privilege promoted by the presentation of peptide on tolerogenic B cells in the spleen. II. Evidence for presentation by Qa-1. J. Immunol. 166, 26–32 (2001).
Tsitoura, D.C., Yeung, V.P., DeKruyff, R.H. & Umetsu, D.T. Critical role of B cells in the development of T cell tolerance to aeroallergens. Int. Immunol. 14, 659–667 (2002).
Mizoguchi, A. & Bhan, A.K. A case for regulatory B cells. J. Immunol. 176, 705–710 (2006).
Mauri, C. & Ehrenstein, M.R. The ‘short’ history of regulatory B cells. Trends Immunol. 29, 34–40 (2008).
Singh, A. et al. Regulatory role of B cells in a murine model of allergic airway disease. J. Immunol. 180, 7318–7326 (2008).
Yiamouyiannis, C.A. et al. Shifts in lung lymphocyte profiles correlate with the sequential development of acute allergic and chronic tolerant stages in a murine asthma model. Am. J. Pathol. 154, 1911–1921 (1999).
Schramm, C.M. et al. Chronic inhaled ovalbumin exposure induces antigen-dependent but not antigen-specific inhalational tolerance in a murine model of allergic airway disease. Am. J. Pathol. 164, 295–304 (2004).
Swirski, F.K., Sajic, D., Robbins, C.S., Gajewska, B.U., Jordana, M. & Stampfli, M.R. Chronic exposure to innocuous antigen in sensitized mice leads to suppressed airway eosinophilia that is reversed by granulocyte macrophage colony-stimulating factor. J. Immunol. 169, 3499–3506 (2002).
Sakai, K., Yokoyama, A., Kohno, N., Hamada, H. & Hiwada, K. Prolonged antigen exposure ameliorates airway inflammation but not remodeling in a mouse model of bronchial asthma. Int. Arch. Allergy Immunol. 126, 126–134 (2001).
Van Hove, C.L., Maes, T., Joos, G.F. & Tournoy, K.G. Prolonged inhaled allergen exposure can induce persistent tolerance. Am. J. Respir. Cell Mol. Biol. 36, 573–584 (2007).
Jungsuwadee, P., Benkovszky, M., Dekan, G., Stingl, G. & Epstein, M.M. Repeated aerosol allergen exposure suppresses inflammation in B-cell-deficient mice with established allergic asthma. Int. Arch. Allergy Immunol. 133, 40–48 (2004).
Singh, A. et al. Subcutaneous late phase responses are augmented during local inhalational tolerance in a murine asthma model. Immunol. Cell Biol. 86, 535–538 (2008).
Matsushita, T. & Tedder, T.F. Identifying regulatory B cells (B10 cells) that produce IL-10 in mice. In Suppression and Regulation of Immune Responses, Methods in Molecular Biology, Vol 677 (Cuturi, M.C. & Anegon, I., eds) 99–111 ( Springer Science+Business Media, New York, 2011 ).
Berland, R. & Wortis, H.H. Origins and functions of B-1 cells with notes on the role of CD5. Annu. Rev. Immunol. 20, 253–300 (2002).
Cyster, J.G. et al. Follicular stromal cells and lymphocyte homing to follicles. Immunol. Rev. 176, 181–193 (2000).
Forster, R., Mattis, A.E., Kremmer, E., Wolf, E., Brem, G. & Lipp, M. A putative chemokine receptor, BLR1, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell 87, 1037–1047 (1996).
Allen, C.D. et al. Germinal center dark and light zone organization is mediated by CXCR4 and CXCR5. Nat. Immunol. 5, 943–952 (2004).
Sebastiani, S. et al. Chemokine receptor expression and function in CD4+ T lymphocytes with regulatory activity. J. Immunol. 166, 996–1002 (2001).
Lundy, S.K., Berlin, A.A., Martens, T.F. & Lukacs, N.W. Deficiency of regulatory B cells increases allergic airway inflammation. Inflamm. Res. 54, 514–521 (2005).
Cariappa, A. et al. The follicular versus marginal zone B lymphocyte cell fate decision is regulated by Aiolos, Btk, and CD21. Immunity 14, 603–615 (2001).
Youinou, P., Jamin, C. & Lydyard, P.M. CD5 expression in human B-cell populations. Immunol. Today 20, 312–316 (1999).
Lemoine, S., Morva, A., Youinou, P. & Jamin, C. Human T cells induce their own regulation through activation of B cells. J. Autoimmun. 36, 228–238 (2011).
Yanaba, K., Bouaziz, J.-D., Haas, K.M., Poe, J.C., Fujimoto, M. & Tedder, T.F. A regulatory B cell subset with a unique CD1dhiCD5+ phenotype controls T cell-dependent inflammatory responses. Immunity 28, 639–650 (2008).
Bouaziz, J.D., Yanaba, K. & Tedder, T.F. Regulatory B cells as inhibitors of immune responses and inflammation. Immunol. Rev. 224, 201–214 (2008).
Mangan, N.E., Fallon, R.E., Smith, P., van Rooijen, N., McKenzie, A.N. & Fallon, P.G. Helminth infection protects mice from anaphylaxis via IL-10-producing B cells. J. Immunol. 173, 6346–6356 (2004).
Amu, S., Saunders, S.P., Kronenberg, M., Mangan, N.E., Atzberger, A. & Fallon, P.G. Regulatory B cells prevent and reverse allergic airway inflammation via FoxP3-positive T regulatory cells in a murine model. J. Allergy Clin. Immunol. 125, 1114–1124 (2010).
Wilson, M.S. et al. Helminth-induced CD19+CD23hi B cells modulate experimental allergic and autoimmune inflammation. Eur. J. Immunol. 40, 1682–1696 (2010).
Noh, J. et al. Characterisation of allergen-specific responses of IL-10-producing regulatory B cells (Br1) in cow milk allergy. Cell Immunol. 264, 143–149 (2010).
Lee, J.H. et al. Allergen-specific transforming growth factor-β-producing CD19+CD5+ regulatory B-cell (Br3) responses in human late eczematous allergic reactions to cow's milk. J. Interferon Cytokine Res. 31, 441–449 (2011).
Carson, W.F. IV, Guernsey, L.A., Singh, A., Vella, A.T., Schramm, C.M. & Thrall, R.S. Accumulation of regulatory T cells in the local draining lymph nodes of the lung correlates with the spontaneous resolution of a chronic model of murine asthma. Int. Arch. Allergy Immunol. 145, 231–243 (2008).
Khanna, K.M., Blair, D.A., Vella, A.T., McSorley, S.J., Datta, S.K. & Lefrançois, L. T cell and APC dynamics in situ control the outcome of vaccination. J. Immunol. 185, 239–252 (2010).
Acknowledgements
This work was supported by NIH grants R21–AI079533 and R01–HL43573.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Supplementary information
Rights and permissions
About this article
Cite this article
Natarajan, P., Singh, A., McNamara, J. et al. Regulatory B cells from hilar lymph nodes of tolerant mice in a murine model of allergic airway disease are CD5+, express TGF-β, and co-localize with CD4+Foxp3+ T cells. Mucosal Immunol 5, 691–701 (2012). https://doi.org/10.1038/mi.2012.42
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2012.42
This article is cited by
-
B cells oppose Mycoplasma pneumoniae vaccine enhanced disease and limit bacterial colonization of the lungs
npj Vaccines (2022)
-
Regulatory B cells: the cutting edge of immune tolerance in kidney transplantation
Cell Death & Disease (2018)
-
Early-life antibiotics attenuate regulatory T cell generation and increase the severity of murine house dust mite-induced asthma
Pediatric Research (2018)
-
Regulatory B and T lymphocytes in multiple sclerosis: friends or foes?
Autoimmunity Highlights (2018)
-
Activation of human B cells negatively regulates TGF-β1 production
Journal of Neuroinflammation (2017)
