
Abstract
Tissue factor (TF) is important for initiation of coagulation and for the increased thrombin activity observed at sites of inflammation. Thrombin activity is induced by allergen challenge in asthmatic airways and is involved in the pathogenesis of asthma. A −603A → G polymorphism (rs1361600) in the promoter region of the TF gene has been associated with serum TF levels and with the development of cardiovascular diseases. The aim of this study was to determine whether the functional −603A → G polymorphism has genetic influences on the development of asthma. Case–control analysis was performed of the association between six common single-nucleotide polymorphisms (SNPs), including the −603A → G polymorphism, at the TF gene, and the development of asthma, using two unrelated Japanese populations. In the primary population (n=826), the GG genotype at the −603A → G polymorphism was associated with adult-onset asthma (onset at ⩾21 years of age) (odds ratio (OR) 2.886, P=0.0231). A second population showed a similar tendency (n=1654, OR 1.602, P=0.064). Transcriptional activity of promoters with −603A → G genotypes were examined using luciferase promoter assays. The −603G allele was associated with higher promoter activity (P<0.05). The association between the functional polymorphism (−603A → G) in the TF gene promoter and adult-onset asthma indicates that TF is a candidate gene contributing to asthma susceptibility.
Similar content being viewed by others


Introduction
Several previous reports suggest that activation of coagulation is involved in the pathogenesis of inflammatory pulmonary diseases,1, 2, 3, 4 although the precise role of coagulation abnormalities in pulmonary pathology is still unclear. It has been established that the coagulation system is activated in human asthma and that the levels of thrombin, the enzyme generated by the activation of coagulation, are consequently increased in the airways.5, 6 Thrombin has been shown to induce a variety of cell responses involved in the pathogenesis of asthma, such as mast cell degranulation7 and airway hyperreactivity8 and remodeling.9 Genetic polymorphisms of key coagulation factor genes are associated with an increased risk of adverse outcomes in illnesses such as deep venous thrombosis and stroke, and, in some cases, of acute lung injury and pulmonary fibrosis.10, 11, 12 Recently, we showed that polymorphisms in the plasminogen-activator inhibitor-1 gene, which has shown a significant association with cardiovascular diseases in several studies, are associated with the development of asthma.13 These results strongly suggest that activation of the coagulation system in the airway has a significant role in the pathogenesis of asthma.
Tissue factor (TF) is a 43-kDa transmembrane cell surface glycoprotein, which, after forming a complex with factor VIIa, activates the extrinsic pathway of the coagulation system. In response to tissue injury and stress, TF has an important role in the initiation of coagulation and in the increase in thrombin activity at the site of inflammation. In a murine model, when coagulation is blocked at the TF-activated factor VII level during experimental sepsis or acute respiratory distress syndrome, inflammatory responses in the lung are attenuated,14, 15, 16 indicating that activation of coagulation by TF is involved in the pathogenesis of inflammatory lung diseases, including asthma. The human TF gene is located on chromosome 1, spans 12.4 kb, and is organized into six exons separated by five introns. Polymorphisms with potential functional relevance have been identified in the TF promoter.10 Of these polymorphisms, the −603G allele has been analyzed most extensively and has been associated with increased TF gene mRNA expression in human monocytes17 and with higher serum levels of TF.10, 11 Further evidence of the functional importance of these variants lies in the significantly increased risk of myocardial infarction11 and venous thrombosis10 in the carriers of risk alleles.
Given the potential association of an activated coagulation system with airway inflammation, particularly the role of TF as an important initiator of the coagulation system, the TF gene is an excellent candidate for asthma susceptibility. In this study, using a case–control analysis, we evaluated the association of TF polymorphisms in correlation with the development of asthma. In particular, the −603A → G polymorphism, which has been associated with the development of cardiovascular diseases, was correlated to the development of asthma. To gain further insight into the possible molecular basis of this disease association, we also examined the functional consequences of this polymorphism at the regulatory region of the TF gene in vitro.
Materials and methods
Study populations
Two independent populations of Japanese asthmatic patients and control subjects were recruited. The demographic characteristics of the primary population (primary study population) and the replicate population (second study population) are presented in Table 1.
Primary study population
A total of 826 unrelated Japanese subjects, including 437 patients with asthma and 389 healthy volunteers, were recruited as the primary study population (Table 1). All patients with asthma were recruited from the pulmonary clinic of the First Department of Medicine, Hokkaido University Hospital. Asthmatic patients were defined as patients with recurrent episodes of at least two of three asthma symptoms (coughing, wheezing and dyspnea) and with a demonstrable reversible airflow limitation (15% variability in forced expiratory volume in 1 s, or in peak expiratory flow rate, either spontaneously or with an inhaled short-acting β2-agonist) and/or with increased airway responsiveness to methacholine as previously described.18, 19 Individuals who visited our clinic for annual routine physical examinations, as well as healthy students at the Hokkaido University School of Medicine, were recruited as non-asthmatic controls if they had no history of asthma or any other chronic pulmonary diseases. Total serum immunoglobulin-E (IgE) levels (IU ml−1) were measured in all subjects, and specific IgE responses to 10 common inhaled allergens, including Der f, grass pollens, animal danders and molds, were assessed. Atopic status was defined as a positive response to at least 1 of the 10 common inhaled allergens, as previously described.18, 19 An increase in specific IgE antibody levels (IgE CAP RAST >0.35 UA ml−1, or MAST >1.0 lumicount) was considered to be a positive response. All subjects were unrelated and were Japanese. Written informed consent was obtained from all the subjects for enrollment in the study. The Ethics Committee of the School of Medicine, Hokkaido University, approved the study.
Replicate study population
A total of 1654 unrelated Japanese subjects, including 932 patients with asthma and 722 healthy volunteers, were used as the replicate study population (Table 1). All subjects with asthma were diagnosed according to the criteria of the National Institute of Health as previously described.20, 21 The diagnosis of atopic asthma was based on one or more positive skin scratch test responses to seven common aeroallergens in the presence of a positive histamine control and a negative vehicle control. The seven aeroallergens were house dust, Felis domesticus dander (Feld), Canis familiaris dander, Dactylis glomerata, Ambrosia, Cryptomeria japonica and Alternaria alternate. A total of 722 healthy individuals, without any respiratory symptoms or history of asthma-related diseases, were recruited on the basis of a physician’s interview, in which they were asked whether they had been diagnosed as having asthma and/or atopy. All asthmatic patients were atopic and no information was obtained regarding the atopic status of healthy controls. No information regarding the smoking status or the total serum IgE level was obtained for this population. All subjects were unrelated Japanese who provided their written informed consent for enrollment in the study. The Ethics Committee of the SNP Research Center, The Institute of Physical and Chemical Research (RIKEN), approved the study.
Genotyping
We initially genotyped the −603A → G polymorphism in the primary population because of reports linking this polymorphism as functionally relevant to several cardiovascular diseases.10, 11, 17 We selected an additional five single-nucleotide polymorphisms (SNPs) for genotyping on the basis of the frequency and location of SNPs and the linkage disequilibrium structure in and around the TF gene. We initially obtained genotyping data for 17 HapMap SNPs (spanning 15 kb around the gene) from the International HapMap Project (available online at http://www.hapmap.org/). To select tagSNPs in this region, we used the multimarker predictor method implemented in the Tagger program.22 The Tag set was generated (using a threshold r2 of 0.8) using five common SNPs with a minor allele frequency of more than 0.1 in the Japanese population.
Genotypes of subjects were identified by an assay that combined kinetic real-time quantitative PCR with allele-specific amplification as previously described.18, 19 Real-time PCR was performed using the SYBR Green I Master Mix (Applied Biosystems, Foster City, CA, USA) and an ABI PRISM TM 7700 Sequence Detection System (Applied Biosystems). In addition, 25 samples were directly sequenced to confirm the accuracy of genotyping using allele-specific PCR. The sequences of the primers used for allele-specific PCR and direct sequencing are shown in Supplementary Tables S1 and S2. The −603A → G polymorphisms of the replicate population were genotyped using the TaqMan system (Applied Biosystems).
Cell culture
BEAS-2B cells were cultured in Dulbecco's modified Eagle's medium/F-12 medium supplemented with 10% heat-inactivated fetal bovine serum at 37 °C in 5% CO2-containing humidified air. Human umbilical vein endothelial cells were obtained from Cambrex (Walkersville, MD, USA) and were cultured in endothelial cell basal medium (EBM-2, Cambrex) maintained at 37 °C in 5% CO2 and supplemented with 5% fetal bovine serum, penicillin/streptomycin and endothelial cell growth supplement (SingleQuots, Cambrex).
Construction of TF reporter vectors
Reporter plasmids containing the TF −603A or −603G alleles were constructed as follows. A PCR product of 864 bp was generated from the −603A or −603G alleles using genomic DNA templates from individuals homozygous for either A or G at position −603. The PCR products, including the −603A → G polymorphism, were digested with KpnI/BglII, and then directionally cloned into the KpnI and BglII sites of the promoterless, enhancerless luciferase (luc) reporter plasmid, pGL4.10 (Promega, Madison, WI, USA), generating −603A-luc and −603G-luc, respectively. The pGL4.10 plasmid was denoted by pGL4.10basic. The constructs were verified by direct sequence analysis and prepared for transfection using the EndoFree Plasmid Maxi kit (Qiagen, Valencia, CA, USA).
Transfection and dual-luciferase assay
The cell lines BEAS-2B or human umbilical vein endothelial cells were individually plated in the wells of six-well plates. The plasmids −603A-luc, −603G-luc or pGL4.10 (1.0 μg) were transiently co-transfected into the same cell as the control Renilla luciferase reporter plasmid (pGL4.74, 1.0 μg; Promega) using Superfect (Qiagen). Cells were then incubated at 37 °C in 5% CO2. Following 24 h incubation in growth medium, luciferase activity was measured using the Dual-Luciferase Reporter Assay system (Promega) and a TD 20/20 luminometer (Promega). The results were normalized for Renilla activity and expressed as relative luciferase activity.
Electrophoretic mobility shift assay
Nuclear extracts were prepared from BEAS-2B cells using the Nuclear Extraction Kit (Chemicon, Temecula, CA, USA) and were frozen at −80 °C. The protein concentration of the extract was determined using the Bio-Rad protein assay kit (Bio-Rad, Hercules, CA, USA). Nuclear extract (10 μg protein) was mixed for 30 min at room temperature with biotin-labeled, double-stranded oligonucleotides using an EMSA kit (Panomics, Redwood, CA, USA) according to the manufacturer’s instructions. The oligonucleotides used corresponded to the promoter region of TF spanning −603A (5′-TCAAGAATACTTGGCCTGCC-3′ and 5′-GGCAGGCCAAGTATTCTTGA-3′) and −603G (5′-TCAAGAATACCTGGCCTGCC-3′ and 5′-GGCAGGCCAGGTATTCTTGA-3′), and were synthesized by Hokkaido System Science (Sapporo, Japan). Nuclear extract (10 μl) was subjected to 6% polyacrylamide gel electrophoresis at 120 V for 55 min in Tris-borate EDTA buffer (Bio-Rad), followed by transfer to a nylon transfer membrane (Pall biodyne B nylon membrane, Pall, East Hills, NY, USA) using the Trans-Blot SD semi-dry transfer cell system (Bio-Rad). The membranes were then ultraviolet cross-linked (CL-1000; Funakoshi, Tokyo, Japan) for 3 min. Detection was performed according to the manufacturer’s instructions after the membranes were exposed on Hyperfilm ECL (Amersham Biosciences, Chandler, AZ, USA). For the super-shift and competition assays, 2 μl of the antibodies E2F-1 or USF-1, E47 or NF-1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), or 2 μg of the respective non-labeled synthesized oligonucleotides were added to 10 ng of the biotin-labeled oligonucleotide (Hokkaido System Science).
Transcription factor binding site identification
Transcription factor binding sites were predicted using the TFSEARCH (http://www.cbrc.jp/research/db/TFSEARCH.html) and TESS (http://www.cbil.upenn.edu/cgi-bin/tess/tess) databases.
Statistical analyses
The χ2-test was used to compare qualitative risk factors (sex, smoking status and atopic status) among the cases and controls. One-way analysis of variance was used to compare the age of the subjects. The Haploview program23 was used to compare the observed number of genotypes with the number of genotypes expected from the Hardy–Weinberg equilibrium. Statistical analyses were based on the calculation of odds ratio (OR) to provide estimates of the relative risk of asthma. Logistic regression analysis was used to estimate OR adjusted for sex, age and smoking status (current, ex or never). The linkage disequilibrium structure was examined using the Haploview software (http://www.broad.mit.edu/mpg/haploview).23 For haplotype analysis, the Haplo.score program was used, which adjusts for covariates and calculates P-values for each haplotype.24 A study-wide analysis combining two case–control populations was performed using the Cochran–Mantel–Haenszel method. To further examine the relationship between age at the onset of asthma and the genetic effects of the −603A → G allele, we performed survival analyses with age at the onset of asthma as the primary outcome using all subjects. The subgroups stratified according to the TF −603A → G genotypes were analyzed for the time taken for development of asthma using the standard Kaplan–Meier method, which plots the proportion of the population that is asthma-free based on the subjects’ age at the time of evaluation. Relative promoter activity in the luciferase reporter assay was compared between the −603A → G genotypes using an unpaired t-test. These statistical analyses were performed on a personal computer with the statistical packages SYSTAT for Windows (version 10.2, Systat, Chicago, IL, USA) and SPSS for Windows (SPSS Japan, Tokyo, Japan).
Results
Analysis of the primary population
The demographic characteristics of the 389 healthy controls and 437 patients with asthma from the primary population are listed in Table 1. The median age of subjects with asthma was significantly higher compared with the healthy control subjects (P<0.0001). There were significantly more females in the asthma group than in the control group (P<0.0001). Over one half of the subjects with asthma were never smokers, which was a lower number than that for controls (55 vs 68%; χ2-test, P<0.01). More than 50% of the control subjects had atopic status, a result consistent with recent findings that the prevalence of atopy (as indicated by specific IgE against common inhaled allergens) among the Japanese population is increasing.25, 26
Supplementary Table S3 shows the genotype frequencies of six TF gene polymorphisms for all 826 subjects in the primary population. No significant deviation from Hardy–Weinberg equilibrium was observed in healthy control subjects (P>0.05). Of the SNPs examined, there was a significant association only of the −603A → G polymorphism with asthma (χ2-test, P<0.05, Supplementary Table S3). As these results were in accordance with the results of the initial screen of the association analysis and with several previous studies that showed an association of this polymorphism with cardiovascular diseases,10, 11 we therefore further analyzed the association of this polymorphism with asthma.
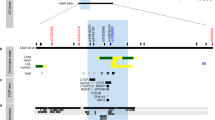
The linkage disequilibrium structure was examined using the Haploview software.23 Analysis of the data from the six SNPs using this program identified a haplotype block (Supplementary Figure S1) in the primary case–control population. Haplotype block I comprised three SNPs in the promoter and coding region (−1953AG [rs958587], −603A → G [rs1361600] and 1973AG [rs696619]). The frequency of the TF haplotype is shown in Figure 1. Haplotype analysis was performed in block I that contained the −603A → G polymorphism. However, none of the haplotypes in block I was associated with asthma (Supplementary Table S4).

Kaplan–Meier survival analysis by TF −603 genotype (primary population). The plots show the effect of the TF −603 genotype on age of asthma onset. (solid line: GG, dotted line: AA or AG). *P=0.033.
The normal aging process alters the blood coagulation system in humans.27 It is interesting that epidemiologic studies show that adult-onset asthma, age of onset ⩾21 years in particular, is associated with cardiovascular diseases,28 carotid atherosclerosis,29 coronary heart disease and stroke,28 in which TF has a significant role during disease development. Therefore, we analyzed the possible effects of the −603A → G genotypes on the age of asthma onset using the Kaplan–Meier method of estimation. Figure 1 shows a plot of the proportions of subjects that were asthma-free versus their age of asthma onset for control (TF −603GG) and −603 AG/AA genotypes. In the plot for the proportion of asthma-free subjects, the slope for the −603GG carriers becomes steeper around 20 years of age. Statistical analyses indicated a significant difference between the two estimated survival curves for the −603A → G genotypes (P<0.05). In light of the results of the Kaplan–Meier curve and the interest in the associations between cardiovascular diseases and adult-onset asthma (age of onset ⩾21 years), in this study, we divided the patients with asthma into two groups: child-onset asthma (age of onset <21 years, n=135) and adult-onset asthma (age of onset ⩾21 years, n=302). Patients with child-onset asthma were highly atopic as a population compared with healthy controls (Table 1). Compared with child-onset asthma, patients with adult-onset asthma had a lower level of mean total serum IgE and a lower frequency of atopy. The GG genotype was significantly associated with adult-onset asthma (OR 2.886; 95% confidence interval 1.156–7.202; P=0.023). In contrast, there were no significant differences in genotype distribution of the −603A → G polymorphism between healthy controls and patients with child-onset asthma (OR 1.72; P=0.377) (Table 2 and Supplementary Table S5).
Analysis of the replicate population
To confirm this association between the −603A → G polymorphism and adult-onset asthma, we determined whether we could replicate the association in an independent population. The demographic characteristics of the 722 healthy controls and 932 patients with asthma from the replicate population are shown in Table 1. The median age of subjects with asthma was significantly higher compared with healthy controls (P<0.0001). There were significantly more females in the asthma group than in the control group (P<0.0001). Genotype frequencies of the −603A → G polymorphism are presented in Supplementary Table S5. No significant deviation from the Hardy–Weinberg equilibrium was observed in healthy controls (P>0.05).
Patients with asthma were divided into two groups consisting of 196 child-onset and 736 adult-onset asthma as defined in the primary population (Table 1). Although it was not statistically significant, a similar tendency of association was observed in this second population, in which the frequency of individuals carrying the −603 GG genotype was higher in the adult-onset asthmatic group (logistic regression analysis, P=0.064) (Table 2 and Supplementary Table S5).
Analysis of the combined populations
Analysis of the combined populations by the Cochran–Mantel–Haenszel test showed that the OR for the GG genotype (AA or AG vs GG) was 1.595 (95% confidence interval 1.082–2.351; P=0.018) (Table 3). This genotype was consistently associated with adult-onset asthma in a pooled sample when the analysis was adjusted for age and sex using the logistic regression model. The OR for the GG homozygotes of the −603A → G polymorphism was 1.736 compared with the A allele (95% confidence interval 1.130–2.667; P=0.0117) (Table 3). In contrast, there were no significant differences in genotype distribution of the −603A → G polymorphism between healthy controls and patients with child-onset asthma (OR 1.207, P=0.559) (Table 3).
The −603A → G polymorphism modulates the transcription of the TF gene
To directly confirm the allele-specific effects of the TF −603A → G polymorphism on native promoter activity, two luciferase reporter gene constructs were generated, spanning 864 bp of the TF promoter region (736 bp of the TF 5′-flank and 128 bp of the 5UTR), with an A or G at the −603 polymorphic site (Figure 2a). The promoter region contained within these constructs has three known sequence variants other than −603A → G. Variances of all other known SNPs in the promoter region of TF were excluded from these constructs so that the data would reflect the functional impact of the −603A → G polymorphism. These constructs were transiently transfected into BEAS-2B and human umbilical vein endothelial cells. As shown in Figure 2b, the relative luciferase activities driven by the mutant G-allelic TF promoter were 30% of those driven by the A-allelic TF promoter in both of the cell lines examined (P<0.05). These results clearly indicated that the G-allelic TF promoter is associated with increased transcriptional activity.

Comparison of the luciferase activity of the A-allele and G-allele promoter-reporter constructs after transient transfection. (a) Schematic outline of the reporter constructs in the pGL4 vector (Luc, luciferase). (b) BEAS 2B (n=10) and human umbilical vein endothelial cells (n=8) were transfected with 1.0 μg of pGL4.74 and with an A-allele (A) or G-allele (G) promoter-reporter plasmid and were harvested 24 h later. The data are presented as the means fold increase ±s.d. relative to the luciferase activity of a Renilla luciferase reporter plasmid (pGL4.74). *P=0.032, **P=0.0015.
Allele-specific binding of nuclear proteins to the TF promoter
Electrophoretic mobility shift assay, using probes corresponding to the 20-bp sequence surrounding the −603A → G site, confirmed the binding of several transcription factors, and the varying affinity of A and G alleles for these factors, at this polymorphic site. Electrophoretic mobility shift assay was performed using nuclear extracts from BEAS-2B cells that were incubated with biotin-labeled oligonucleotide probes corresponding to the −603A or −603G allele. A specific DNA/nuclear extract complex was generated by both the A-allele and G-allele probes. Higher levels of this complex were observed with the −603G allele probe than with the −603A allele probe (Figure 3), suggesting that the −603G allele had a higher affinity for this binding factor and that the −603A → G transition might inhibit or eliminate this binding site. This DNA–protein complex was completely eliminated by 200-fold unlabeled −603A-allele or G-allele probes, indicating that this DNA–protein binding is likely specific for the −603A allele. As the factor does bind, although more weakly, to the −603A allele, this further suggests that the −603A → G transition might inhibit or eliminate this binding site (Figure 3). Two website databases (TESS and TFSEARCH) predicted that sequences surrounding the −603A → G polymorphic site in the promoter region of the TF gene could potentially bind the transcription factors E2F-1, E47, NF-1 or USF-1. However, none of the antibodies to these transcription factors revealed a supershifted complex in the presence of the −603A or −603G allele (data not shown).

Electrophoretic mobility shift assay of fragments of the −603A → G region of the TF gene with nuclear extracts from BEAS 2B. Genomic fragments containing −603A or −603G were synthesized and used as DNA probes. A 200-fold molar excess of cold oligonucleotides was added in lanes 3 and 4. Increased formation of a specific DNA–protein complex was observed with the G-allele when compared with the A-allele (black arrow). *A:A allele, G:G allele, **BEAS-2B cells.
Discussion
In this study, we showed that the −603A → G polymorphism at the promoter region of the TF gene is associated with the development of adult-onset asthma in a Japanese population. Further evidence of the importance of the −603G allele in the pathogenesis of asthma was provided by the observed significant association of this allele with increased transcriptional activity and strong binding of transcription factor(s) in vitro. Among several molecules involved in the coagulation system, TF has an important role in the initiation of coagulation and in the increase in thrombin activity at the site of inflammation.30, 31 TF binds activated factor VII, resulting in activation of factor IX and factor X and ultimately leading to thrombin and fibrin formation. In addition, TF has a role in tissue fibrosis through G-protein-coupled protease-activated receptors.32 In vivo activation of protease-activated receptors leads to proinflammatory responses,33, 34 bronchoconstriction35 and airway remodeling.36, 37 The level of TF in the sputum is higher in asthma patients than in healthy controls.5 The combined data suggest that TF might be involved in the pathogenesis of asthma through its participation in a number of different pathways. Our findings in this genetic study are in line with this contention.
Asthma is a phenotypically heterogeneous disorder that results from complex interactions between environmental and genetic factors.38 A precise definition of the asthma phenotype is now becoming more relevant not only to increase our understanding of pathophysiologic mechanisms but also to ascertain the specific genes associated with these phenotypes. The Kaplan–Meier analysis identified an age of 20 years at onset as the cutoff point for assigning subjects to a child-onset group and an adult-onset group. This approach made it possible to avoid prespecification of an arbitrary threshold and may be suitable for identification of homogeneous subsets for association studies. A cutoff age of 20 years is consistent with the observation that child-onset asthma and adult-onset asthma have distinct pathobiologic mechanisms. Patients with child-onset asthma were shown to have a significantly greater likelihood of allergic sensitization and clinical responses to triggers than patients with adult-onset asthma. Genetic factors related to atopy, including genes on chromosomes 5q31 and 11q13, are involved in child-onset asthma.39, 40, 41, 42 In contrast, asthma that develops during adulthood is more heterogeneous and a family history of asthma is often nonexistent. Although the role of genetic predisposition in adult-onset asthma is less clear than in atopic childhood-onset asthma, a specific host genetic factor might be important in the development of adult-onset asthma. It is noted that we previously reported a functional polymorphism in the CCL5/RANTES gene promoter that was associated with late-onset asthma, that is, starting after 40 years of age, in a Japanese population.43 This study may have identified a new genetic factor involved in the development of adult-onset asthma.
Several epidemiologic studies showed that adult-onset asthma (age of onset ⩾21 years) in particular is associated with cardiovascular diseases,28 carotid atherosclerosis,29 coronary heart disease or stroke.28 Asthma may predispose an individual to atherosclerosis through specific pathobiologic pathways, perhaps linked to the chronic inflammatory responses of this disorder. Alternatively, the association between asthma and atherosclerosis may be caused by an inherent joint susceptibility to both diseases through shared inflammatory pathways and genetic factors. It is noted that the −603G gain-of-function allele in the TF gene has been associated with an increased risk of myocardial infarction11 and venous thrombosis.10 These data, together with the data presented in this study, suggest that TF might be a common genetic factor for the development of both asthma and several cardiovascular diseases. Further studies that incorporate precise data regarding the presence of cardiovascular diseases will help to clarify this speculation.
Several environmental factors have been identified, such as strong inducers of TF gene expression, including viruses,44 bacteria,45, 46, 47 other microbial pathogens such as cytomegalovirus, Chlamydia pneumoniae, Streptococcus sanguis and Plasmodium falciparum,48, 49, 50 endotoxin51 and oxidative air pollutant particulate matter.52 In this study, we showed that the G allele in the −603 position is associated with a higher promoter activity than the A allele in vitro. Therefore, in individuals possessing the potentially functional −603GG genotype, TF may be expressed to a greater extent in response to an exogenous stimulus, ultimately leading to an exaggerated inflammation of the airways. Prolonged and repeated exposure to exogenous stimuli may be required for the −603GG genotype to manifest its genetic effects in airways, which may in part explain why the association of this allele was found only with adult-onset asthma but not with child-onset asthma. It has also been noted that the normal aging process alters the blood coagulation system in humans.27 The plasma concentration of several coagulation factors increases in healthy humans in parallel with the physiological aging process.53 In mice studies, stress-induced expression of TF in several tissues was substantially higher in aged mice than in young mice.54 Accordingly, TF expression in subjects with the −603GG genotype may be enhanced to a greater extent by aging when compared with the carriers of other genotypes, and adult-onset asthma subsequently develops in these individuals, which is an alternative explanation for the heterogeneity of the genetic effects of the TF gene according to age at onset of the disease.
Sequence analysis indicated that, at the site of the −603A → G polymorphism, the A allele creates a potential E2F-1 or NF-1 binding site that might function in suppression of transcription according to the TESS database, and that the G allele creates a potential E47 or USF-1 binding site that might function in enhancement of transcription according to the TFSEARCH database. A specific DNA/nuclear protein complex was formed by the A-allele and G-allele probes. Furthermore, the G-allele showed enhanced formation of this specific DNA–protein complex compared with the A-allele. However, no difference in specific binding of these nuclear factors to the −603A → G polymorphism could be observed. Further studies are required to identify the specific transcription factor(s) that bind to this region, which may enable a demonstration of different binding affinities and thereby explain the varying transcriptional activity induced by this SNP.
A similar trend for association of the −603A → G polymorphism in the TF gene and adult-onset asthma was observed in the second population, although it was not statistically significant (P=0.064). Differences in several confounding factors between the two populations, which were recruited from geographically different areas of Japan, might explain this observation, as asthma is a complex disease associated with gene–gene and gene–environmental factor interactions. The age and sex distribution, as well as the frequency of atopic subjects, differed between the two populations. Information concerning smoking status was also lacking in the second population. Therefore, given the genetic heterogeneity underlying the pathogenesis of asthma, especially for patients who develop the disease in their adulthood, the existence of such similar trends in both populations supports the result obtained in the primary population.
Case–control studies based on questionnaires are susceptible to recall bias. Definitions of physician-diagnosed asthma have been shown to be specific but somewhat lacking in sensitivity and probably exclude milder cases of asthma.55 In this study, to judge age at onset of asthma as accurately as possible, patients were asked about episodes of dyspnea, wheezing or coughing during childhood and puberty. In cases of uncertainty, the time of earliest respiratory symptoms was designated as age at onset of asthma symptoms, which was used for the calculation of the duration of asthma. A recent study of age at onset of asthma in 4335 Japanese adult patients with asthma found that 26.3% developed the disease before the age of 20 years, and 73.8% developed the disease at 20 years or older.56 This variation in age at onset of adult asthma is similar to that found in this study and is evidence that the self-reported data on age at onset of asthma obtained in this study are reliable.
In conclusion, the results of this case–control study and in vitro functional analysis, combined with the known important roles of the coagulation system in inflammatory lung diseases, suggest that TF is involved in the pathogenesis of asthma. Further study to determine the mechanism by which risk for adult-onset asthma is mediated by specific genetic variations in TF should improve our understanding of the molecular basis of asthma.
References
Idell, S., James, K. K., Levin, E. G., Schwartz, B. S., Manchanda, N., Maunder, R. J. et al. Local abnormalities in coagulation and fibrinolytic pathways predispose to alveolar fibrin deposition in the adult respiratory distress syndrome. J. Clin. Invest. 84, 695–705 (1989).
Welty-Wolf, K. E., Carraway, M. S., Ortel, T. L. & Piantadosi, C. A. Coagulation and inflammation in acute lung injury. Thromb. Haemost. 88, 17–25 (2002).
Kotani, I., Sato, A., Hayakawa, H., Urano, T., Takada, Y. & Takada, A. Increased procoagulant and antifibrinolytic activities in the lungs with idiopathic pulmonary fibrosis. Thromb. Res. 77, 493–504 (1995).
Günther, A., Mosavi, P., Heinemann, S., Ruppert, C., Muth, H., Markart, P. et al. Alveolar fibrin formation caused by enhanced procoagulant and depressed fibrinolytic capacities in severe pneumonia: comparison with the acute respiratory distress syndrome. Am. J. Respir. Crit. Care. Med. 161, 454–462 (2000).
Gabazza, E. C., Taguchi, O., Tamaki, S., Takeya, H., Kobayashi, H., Yasui, H. et al. Thrombin in the airways of asthmatic patients. Lung 177, 253–262 (1999).
Shinagawa, K., Martin, J. A., Ploplis, V. A. & Castellino, F. J. Coagulation factor Xa modulates airway remodeling in a murine model of asthma. Am. J. Respir. Crit. Care. Med. 175, 136–143 (2007).
Cirino, G., Cicala, C., Bucci, M. R., Sorrentino, L., Maraganore, J. M. & Stone, S. R. Thrombin functions as an inflammatory mediator through activation of its receptors. J. Exp. Med. 183, 821–827 (1996).
Wagers, S. S., Norton, R. J., Rinaldi, L. M., Bates, J. H., Sobel, B. E. & Irvin, C. G. Extravascular fibrin, plasminogen activator, plasminogen activator inhibitors, and airway hyperresponsiveness. J. Clin. Invest. 114, 104–111 (2004).
Terada, M., Kelly, E. A. & Jarjour, N. N. Increased thrombin activity after allergen challenge: a potential link to airway remodeling. Am. J. Respir. Crit. Care. Med. 169, 373–377 (2004).
Arnaud, E., Barbalat, V., Nicaud, V., Cambien, F., Evans, A., Morrison, C. et al. Polymorphisms in the 5′ regulatory region of the tissue factor gene and the risk of myocardial infarction and venous thromboembolism: the ECTIM and PATHROS studies. Etude Cas-Témoins de l’Infarctus du Myocarde. Paris Thrombosis case-control Study. Arterioscler. Thromb. Vasc. Biol. 20, 892–898 (2000).
Ott, I., Koch, W., von Beckerath, N., de Waha, R., Malawaniec, A., Mehilli, J. et al. Tissue factor promotor polymorphism −603 A/G is associated with myocardial infarction. Atherosclerosis 177, 189–191 (2004).
Russell, J. A. Genetics of coagulation factors in acute lung injury. Crit. Care. Med. 31, S243–S247 (2003).
Hizawa, N., Maeda, Y., Konno, S., Fukui, Y., Takahashi, D. & Nishimura, M. Genetic polymorphisms at FCER1B and PAI-1 and asthma susceptibility. Clin. Exp. Allergy 36, 872–876 (2006).
Miller, D. L., Welty-Wolf, K., Carraway, M. S., Ezban, M., Ghio, A., Suliman, H. et al. Extrinsic coagulation blockade attenuates lung injury and proinflammatory cytokine release after intratracheal lipopolysacharide. Am. J. Respir. Crit. Care. Med. 26, 650–658 (2002).
Enkhbaatar, P., Okajima, K., Murakami, K., Uchiba, M., Okabe, H., Okabe, K. et al. Recombinant tissue factor pathway inhibitor reduces lipopolysaccharide-induced pulmonary vascular injury by inhibiting leukocyte activation. Am. J. Respir. Crit. Care. Med. 162, 1752–1759 (2000).
Laterre, P. F., Wittebole, X. & Dhainaut, J. F. Anticoagulant therapy in acute lung injury. Crit. Care. Med. 31, S329–S336 (2003).
Reny, J. L., Laurendeau, I., Fontana, P., Bièche, I., Dupont, A., Remones, V. et al. The TF−603A/G gene promoter polymorphism and circulating monocyte tissue factor gene expression in healthy volunteers. Thromb. Haemost. 91, 248–254 (2004).
Hizawa, N., Yamaguchi, E., Takahashi, D., Nishihira, J. & Nishimura, M. Functional polymorphisms in the promoter region of macrophage migration inhibitory factor and atopy. Am. J. Respir. Crit. Care. Med. 169, 1014–1018 (2004).
Maeda, Y., Hizawa, N., Jinushi, E., Honda, A., Takahashi, D., Fukui, Y. et al. Polymorphisms in the muscarinic receptor 1 gene confer susceptibility to asthma in Japanese subjects. Am. J. Respir. Crit. Care. Med. 174, 1119–1124 (2006).
National Heart, Lung, and Blood Institute Guidelines for the diagnosis and management of asthma. National Heart, Lung, and Blood Institute. National Asthma Education Program. Expert Panel Report. J. Allergy. Clin. Immunol. 88, 425–534 (1991).
Hirota, T., Suzuki, Y., Hasegawa, K., Obara, K., Matsuda, A., Akahoshi, M. et al. Functional haplotypes of IL-12B are associated with childhood atopic asthma. J. Allergy. Clin. Immunol. 116, 789–795 (2005).
de Bakker, P. I., Yelensky, R., Pe’er, I., Gabriel, S. B., Daly, M. J. & Altshuler, D. Efficiency and power in genetic association studies. Nat. Genet. 37, 1217–1223 (2005).
Barrett, J. C., Fry, B., Maller, J. & Daly, M. J. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21, 263–265 (2005).
Schaid, D. J., Rowland, C. M., Tines, D. E., Jacobson, R. M. & Poland, G. A. Score tests for association between traits and haplotypes when linkage phase is ambiguous. Am. J. Hum. Genet. 70, 425–434 (2002).
Nakagomi, T., Itaya, H., Tominaga, T., Yamaki, M., Hisamatsu, S. & Nakagomi, O. Is atopy increasing? Lancet 343, 121–122 (1994).
Kusunoki, T., Hosoi, S., Asai, K., Harazaki, M. & Furusho, K. Relationships between atopy and lung function: results from a sample of one hundred medical students in Japan. Ann. Allergy Asthma Immunol. 83, 343–347 (1999).
Sagripanti, A. & Carpi, A. Natural anticoagulants, aging, and thromboembolism. Exp. Gerontol. 33, 891–896 (1998).
Onufrak, S. J., Abramson, J. L., Austin, H. D., Holguin, F., McClellan, W. M. & Vaccarino, L. V. Relation of adult-onset asthma to coronary heart disease and stroke. Am. J. Cardiol. 101, 1247–1252 (2008).
Onufrak, S., Abramson, J. & Vaccarino, V. Adult-onset asthma is associated with increased carotid atherosclerosis among women in the Atherosclerosis Risk in Communities (ARIC) study. Atherosclerosis 195, 129–137 (2007).
Pawlinski, R., Pedersen, B., Schabbauer, G., Tencati, M., Holscher, T., Boisvert, W. et al. Role of tissue factor and protease-activated receptors in a mouse model of endotoxemia. Blood 103, 1342–1347 (2004).
Chu, A. J. Tissue factor mediates inflammation. Arch. Biochem. Biophys. 440, 123–312 (2005).
Ruf, W., Dorfleutner, A. & Riewald, M. Specificity of coagulation factor signaling. J. Thromb. Haemost. 1, 1495–1503 (2003).
Schmidlin, F., Amadesi, S., Dabbagh, K., Lewis, D. E., Knott, P., Bunnett, N. W. et al. Protease-activated receptor 2 mediates eosinophil infiltration and hyperreactivity in allergic inflammation of the airway. J. Immunol. 169, 5315–5321 (2002).
Vergnolle, N. Proteinase-activated receptor-2-activating peptides induce leukocyte rolling, adhesion, and extravasation in vivo. J. Immunol. 163, 5064–5069 (1999).
Ricciardolo, F. L., Steinhoff, M., Amadesi, S., Guerrini, R., Tognetto, M., Trevisani, M. et al. Presence and bronchomotor activity of protease-activated receptor-2 in guinea pig airways. Am. J. Respir. Crit. Care. Med. 161, 1672–1680 (2000).
Blanc-Brude, O. P., Archer, F., Leoni, P., Derian, C., Bolsover, S., Laurent, G. J. et al. Factor Xa stimulates fibroblast procollagen production, proliferation, and calcium signaling via PAR1 activation. Exp. Cell. Res. 304, 16–27 (2005).
Chambers, R. C. & Laurent, G. J. Coagulation cascade proteases and tissue fibrosis. Biochem. Soc. Trans. 30, 194–200 (2002).
Vercelli, D. Genetics, epigenetics, and the environment: switching, buffering, releasing. J. Allergy. Clin. Immunol. 113, 381–386 (2004).
Hizawa, N., Freidhoff, L. R., Ehrlich, E., Chiu, Y. F., Duffy, D. L., Schou, C. et al. Genetic influences of chromosomes 5q31-q33 and 11q13 on specific IgE responsiveness to common inhaled allergens among African American families. Collaborative Study on the Genetics of Asthma (CSGA). J. Allergy. Clin. Immunol. 102, 449–453 (1998).
Postma, D. S., Bleecker, E. R., Amelung, P. J., Holroyd, K. J., Xu, J., Panhuysen, C. I. et al. Genetic susceptibility to asthma-bronchial hyperresponsiveness coinherited with a major gene for atopy. N. Engl. J. Med. 333, 894–900 (1995).
van Herwerden, L., Harrap, S. B., Wong, Z. Y., Abramson, M. J., Kutin, J. J., Forbes, A. B. et al. Linkage of high-affinity IgE receptor gene with bronchial hyperreactivity, even in absence of atopy. Lancet 346, 1262–1265 (1995).
Noguchi, E., Shibasaki, M., Arinami, T., Takeda, K., Maki, T., Miyamoto, T. et al. Evidence for linkage between asthma/atopy in childhood and chromosome 5q31-q33 in a Japanese population. Am. J. Respir. Crit. Care. Med. 156, 1390–1393 (1997).
Hizawa, N., Yamaguchi, E., Konno, S., Tanino, Y., Jinushi, E. & Nishimura, M. A functional polymorphism in the RANTES gene promoter is associated with the development of late-onset asthma. Am. J. Respir. Crit. Care. Med. 166, 686–690 (2002).
Vercellotti, G. M. Effects of viral activation of the vessel wall on inflammation and thrombosis. Blood Coagul. Fibrinolysis 9, S3–S6 (1998).
Drake, T. A. & Pang, M. Staphylococcus aureus induces tissue factor expression in cultured human cardiac valve endothelium. J. Infect. Dis. 157, 749–756 (1988).
Veltrop, M. H., Beekhuizen, H. & Thompson, J. Bacterial species- and strain-dependent induction of tissue factor in human vascular endothelial cells. Infect. Immun. 67, 6130–6138 (1999).
Heyderman, R. S., Klein, N. J., Daramola, O. A., Hammerschmidt, S., Frosch, M., Robertson, B. D. et al. Induction of human endothelial tissue factor expression by Neisseria meningitidis: the influence of bacterial killing and adherence to the endothelium. Microb. Pathog. 22, 265–274 (1997).
Bouwman, J. J., Visseren, F. L., Bosch, M. C., Bouter, K. P. & Diepersloot, R. J. Procoagulant and inflammatory response of virus-infected monocytes. Eur. J. Clin. Invest. 32, 759–766 (2002).
Bea, F., Puolakkainen, M. H., McMillen, T., Hudson, F. N., Mackman, N., Kuo, C. C. et al. Chlamydia pneumoniae induces tissue factor expression in mouse macrophages via activation of Egr-1 and the MEK-ERK1/2 pathway. Circ. Res. 92, 394–401 (2003).
Veltrop, M. H., Thompson, J. & Beekhuizen, H. Monocytes augment bacterial species- and strain-dependent induction of tissue factor activity in bacterium-infected human vascular endothelial cells. Infect. Immun. 69, 2797–2807 (2001).
Erlich, J. H., Fearns, C., Mathison, J., Ulevitch, R. J. & Mackman, N. Lipopolysaccharide induction of tissue factor expression in rabbits. Infect. Immun. 67, 2540–2546 (1999).
Kooter, I., Pennings, J., Opperhuizen, A. & Cassee, F. Gene expression pattern in spontaneously hypertensive rats exposed to urban particulate matter (EHC-93). Inhal. Toxicol. 17, 53–65 (2005).
Ofosu, F. A., Craven, S., Dewar, L., Anvari, N., Andrew, M. & Blajchman, M. A. Age-related changes in factor VII proteolysis in vivo. Br. J. Haematol. 94, 407–412 (1996).
Yamamoto, K., Shimokawa, T., Yi, H., Isobe, K., Kojima, T., Loskutoff, D. J. et al. Aging and obesity augment the stress-induced expression of tissue factor gene in the mouse. Blood 100, 4011–4018 (2002).
Toren, K., Brisman, J. & Jarvholm, B. Asthma and asthma-like symptoms in adults assessed by questionnaires: a literature review. Chest 104, 600–608 (1993).
Akiyama, K., Takahashi, K. & Yanagawa, H. Nation-wide survey of asthma patients in Japan [abstract]. Allergy Clin Immunol Int (Suppl 4), 87 (1997).
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on Journal of Human Genetics website (http://www.nature.com/jhg)
Rights and permissions
About this article
Cite this article
Isada, A., Konno, S., Hizawa, N. et al. A functional polymorphism (−603A → G) in the tissue factor gene promoter is associated with adult-onset asthma. J Hum Genet 55, 167–174 (2010). https://doi.org/10.1038/jhg.2010.4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/jhg.2010.4
Keywords
This article is cited by
-
Japanese-Specific Filaggrin Gene Mutations in Japanese Patients Suffering from Atopic Eczema and Asthma
Journal of Investigative Dermatology (2010)

{kind=link}