

Key Points
- The upper esophageal sphincter (UES) closure muscles include the cervical esophagus, cricopharyngeus, and inferior pharyngeal constrictor, but primarily the cricopharyngeus.
- The main functions of UES closure are to prevent esophageal air insufflation during negative intrathoracic pressure events, for example, inspiration, and to prevent esophagopharyngeal/laryngeal reflux during esophageal peristalsis.
- The UES is opened intermittently by relaxation of its sphincteric muscles, contraction of its distracting muscles, and bolus pulsion.
- The function of intermittent UES opening is to allow transphincteric flow of fluid or gas during orthograde events, for example, swallowing, or antegrade events, for example, emesis.
- The UES opening muscles include anteriorly the superior and inferior hyoid muscles and posteriorly the superior pharyngeal muscles.
- The UES motor and sensory functions are controlled by branches of the glossopharyngeal and vagus nerves.
- The nucleus ambiguus is the primary motor nucleus of the UES, and the nucleus tractus solitarius is the primary termination site of UES afferents.
- The specific actions of individual muscles of the UES depend on the particular function, that is, tone generation, swallowing, belching, retching, or vomiting.
Definition of the Upper Esophageal Sphincter
The upper esophageal sphincter (UES) is that area of the upper digestive tract that forms a barrier between the esophagus and the pharynx, but that intermittently opens and closes to allow passage of contents during various physiologic events. This definition implies three responses: tone generation, phasic response activity, and sphincter opening. Closing of the UES can be considered a phasic function of the UES. The upper esophageal area encompassing the tone generation function of the UES is sometimes referred to as the upper esophageal high-pressure zone (UEHPZ). Although the physiologic concepts of UES and UEHPZ are clear, the anatomic components of these concepts are not.
Upper Esophageal High-Pressure Zone
The UEHPZ is recorded using intraluminal manometric devices, and, based on combined manometric and radiographic techniques, investigators have found that the UEHPZ is associated with three muscles (Figure 1): cervical esophagus, cricopharyngeus (CP), and inferior pharyngeal constrictor (IPC) [or thyropharyngeus (TP) in animals]; therefore, all three muscles probably contribute to the UEHPZ.1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 The peak pressure of the UEHPZ1, 2, 12 was found at the caudal edge of the IPC (Figure 2), but the preponderance of evidence suggests that the primary muscle of the UEHPZ is the cricopharyngeus.
Figure 1: Anatomy of the closing and some opening muscles of the upper esophageal sphincter (UES).
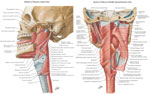
(Source: Netter medical illustration with permission of Elsevier. All rights reserved.)
Figure 2: Relationship of upper esophageal high-pressure zone (UEHPZ) to the pharyngoesophageal muscles.
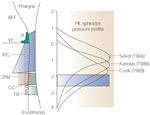
Data compiled from three different studies of the position of the UEHPZ with respect to individual pharyngeal muscles based on combined manometric and videofluoroscopic studies. Note that the peak pressures of the UEHPZ in humans at rest with the head fixed in all three studies coincides with the lower border of the IPC. A, arytenoid; AEF, aryepiglottic fold; CC, cricoid cartilage; ESO, esophagus; IPC, inferior pharyngeal constrictor; PE, pharyngoesophageal; TR, tracheal ring; VC, vocal cord. (Source: Adapted from Goyal et al.,12 with permission of Springer Science and Business Media.)
The pressure recorded intraluminally is the sum of all external forces and is related to active and passive forces of the muscles as well as the compliance of the surrounding structures. Therefore, although peak intraluminal pressure is recorded at the IPC, it is unknown which component of the region of the IPC accounted for this peak pressure. For example, the pressure of the UEHPZ may peak in the region of the IPC because of higher passive forces of the IPC and lower compliance of the IPC muscle or pharyngeal wall, rather than a stronger contraction of the IPC muscle itself. Therefore, the point of highest intraluminal pressure need not correspond with the most active muscle.
Another problem with the conclusion that the IPC is the main muscle of the UEHPZ is that all the investigations that associated peak pressure of the UEHPZ with location of specific muscles were recorded with the subject at rest in a fixed head position.2, 3, 4, 8 It has been found that UES tone13, 14, 15, 16 and CP electromyography (EMG)1 but not TP EMG1 are very variable and change greatly with the state of arousal, excitability, and head position (see Functions of the Upper Esophageal Sphincter, below). It is possible that the very relaxed conditions and the fixed position of the head in these experiments2, 3, 4, 8 resulted in very low states of activation of the CP compared to the TP. Thus, the association of peak pressure of the UEHPZ with location of the TP may hold only at very low levels of arousal and excitability and a fixed head position.
There are many situations when the tone of the UEHPZ increases much above basal conditions, and the reason for this is addressed below (see Functions of the Upper Esophageal Sphincter). Under conditions of high UES tone, the tone increase is usually due to activation of all of the UEHPZ muscles, that is, TP, CP, and cervical esophagus, but primarily the CP.
Therefore, physiologic evidence suggests that although the IPC and cervical esophagus may contribute to the UEHPZ under certain conditions, at all levels of activity or arousal the main muscle contributing to the UEHPZ is the CP.
Upper Esophageal Sphincter
The definition of the UES includes not only tone generation, but also UES opening and phasic changes in UES tone. Phasic changes in UES tone occur during various reflexes and physiologic states (see Functions of the Upper Esophageal Sphincter, below), and in most situations these changes are due to changes in activation of the cervical esophagus, CP, or TP. The opening of the UES depends on three actions: the relaxation of the UES tone-generating muscles, the distraction of the larynx and esophagus away from each other, and bolus pulsion. The distraction forces can overcome the tone generating muscles of UES to open the sphincter2; therefore, these muscles are equally important for one of the major functions of the UES. The TP, CP, and cervical esophagus relax during UES opening, and the superior and inferior hyoid muscles as well as the posterior pharyngeal muscles provide the major distracting forces.
Regardless of reflex or function, the relative contribution of each muscle of the UES depends on the specific function (see Functions of the Upper Esophageal Sphincter, below); however, regardless of the function, the primary muscle of the UES is the CP.
Anatomy and Physiology of the Upper Esophageal Sphincter
Anatomy and Cellular Physiology
Upper Esophageal Sphincter Closure Muscles
Cricopharyngeus The CP is structurally, biochemically, and mechanically distinct from surrounding pharyngeal and esophageal muscles except for the inferior IPC (iIPC) as discussed below. The CP muscle attaches to the dorsolateral aspect of the lower part of the cricoid cartilage, forming a horizontal muscular band. In humans, two sets of muscle fibers have been identified: the horizontally oriented fibers (CPh), pars fundiformis; and an oblique band of fibers (CPo), pars obliqua, that extend from the lateral aspect of the cricoid cartilage to the posterior raphe where they blend superiorly with the IPC.17 Unlike the IPC, the pars fundiformis of the CP has no median raphe. In most animal species the CP forms a muscular band distinct from the TP, but contains a median raphe18, 19.
The CP is a striated muscle composed of variable-sized fibers about 25 to 35  m in diameter.17, 20, 21 Unlike most other striated muscles, the muscle fibers of the CP are not oriented in a strict parallel fashion and they insert onto connective tissue rather than the skeletal framework.22 Therefore, the CP forms a muscular network. The CP contains both slow- (type I, oxidative) and fast-twitch (type II, glycolytic) muscle fibers,17, 21, 22, 23, 24 but unlike surrounding pharyngeal muscles the predominant fiber type is slow twitch.17, 21, 22, 23, 24, 25 The CPh (76%) has more slow-twitch fibers than the CPo (69%), and both parts of the CP are composed of two histochemically different layers.24 In both layers the slow-twitch fibers predominate, but more so in the inner (mean of 84%) than the outer (mean of 59%) layer.24 The presence of both slow- and fast-twitch fibers provides an anatomic basis for the two functions of the CP or UES: maintaining a basal tone and rapidly relaxing or contracting during various functions and reflexes.
m in diameter.17, 20, 21 Unlike most other striated muscles, the muscle fibers of the CP are not oriented in a strict parallel fashion and they insert onto connective tissue rather than the skeletal framework.22 Therefore, the CP forms a muscular network. The CP contains both slow- (type I, oxidative) and fast-twitch (type II, glycolytic) muscle fibers,17, 21, 22, 23, 24 but unlike surrounding pharyngeal muscles the predominant fiber type is slow twitch.17, 21, 22, 23, 24, 25 The CPh (76%) has more slow-twitch fibers than the CPo (69%), and both parts of the CP are composed of two histochemically different layers.24 In both layers the slow-twitch fibers predominate, but more so in the inner (mean of 84%) than the outer (mean of 59%) layer.24 The presence of both slow- and fast-twitch fibers provides an anatomic basis for the two functions of the CP or UES: maintaining a basal tone and rapidly relaxing or contracting during various functions and reflexes.
The optimum length at which the CP reaches maximum active tension is about 1.7 times its basal (or in situ) length,26 whereas this tension in most striated muscles occurs at its resting length (Figure 3). The source of this elasticity may be attributed to the connective tissue, that is, collagen, elastin, sarcolemma, or the contractile proteins actin and myosin.27 The CP contains more elastic connective tissue and sarcolemma than most other striated muscles.22, 25 This high degree of elasticity as well as the network arrangement of muscle fibers contribute to some important and distinctive characteristics of the UES. The UES is capable of maintaining a basal tone without active muscular contraction. The active tension of the UES increases throughout the range of distention26 similar to Starling's law of the heart, allowing a greater force to be exerted by the UES behind a passing bolus. This characteristic would be important to propel larger boluses and to prevent reflux. In addition, the optimum length of the CP is larger than the maximum bolus volume likely to occur26, ensuring that the UES tension will always accommodate to the bolus. Finally the high degree of elasticity of the CP allows the UES to be opened by a bolus or distraction forces without relaxation of the CP.2
Figure 3: Strain-energy relationship of the cricopharyngeus (CP) muscle.
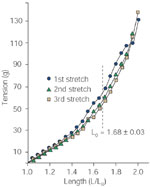
This graph shows that the optimum length (LO) of the CP is approximately 1.7 times its resting length (LR), whereas in other striated muscles LO = LR. This relationship illustrates the large degree of elasticity of the CP compared to other striated muscles. (Source: Medda et al.,26 with permission of the American Physiological Society.)
Inferior pharyngeal constrictor The IPC arises from the sides of the superior portion of the cricoid cartilage and the oblique line on the side of the thyroid cartilage and the fibers spread dorsally and medially to meet the opposite side of the muscle at a fibrous raphe in the posterior median line of the pharynx.28
The IPC is a functionally diverse muscle, as it contains rostral and caudal neuromuscular compartments with different amounts of muscle fiber types.28 The rostral compartment contains more (61%) fast-twitch fibers than the caudal compartment (30%), and each compartment is composed of two layers: an inner layer of slow-muscle fibers and an outer layer of fast-muscle fibers. The slow inner layer of the caudal compartment is twice as thick as its fast outer layer, and this organization is reversed for the rostral compartment. Therefore, the IPC may have two functions: a slower acting, perhaps tone-generating function of its caudal half, and a rapid contracting function of the rostral half. The muscle fibers of the most caudal portion of the IPC, the iIPC, are most similar to those of the more rostral portions of the CP, the CPo. They have similar distributions of slow- (84%) and fast-twitch fibers and both have similar thickness of inner and outer layers (2:1 ratio).
The anatomic and histochemical data suggest that the iIPC and CP function similarly, but no physiologic data are available to confirm this conjecture. Although the iIPC and CP have similar types of muscle fibers, it is possible that the iIPC may have less elasticity than the CP. This difference could result in lower compliance of the iIPC and therefore could account for the peak UEHPZ recorded in the vicinity of the iIPC. Further studies are needed to resolve this issue.
Cervical esophagus The cervical esophagus differs from the remainder of the esophagus in that the muscle fibers are arranged in a strictly horizontal fashion and the fibers are exclusively striated muscle fibers for first 1 to 5 cm.29 The muscle fibers are about the same size as the adjacent CP,30 and some authors have found that fibers from the CP contribute to the adjacent cranial portion of the cervical esophagus.31 The predominate muscle fiber type of the striated muscle esophagus in animals is fast twitch (type II),31, 32, 33 but this issue is disputed in human studies.29, 30, 31, 32 It has been suggested that this discrepancy is due to technical differences in tissue handling.33 Studies in animals33 but not humans30 have found regional differences in fiber type along the length of the esophagus. Although physiologic evidence suggests that the cervical esophagus may at times function as part of the UES, histochemical studies have not provided a biochemical basis for this function.
Upper Esophageal Sphincter Opening Muscles
The UES opening muscles are situated both anteriorly and posteriorly, where they act in concert and have similar anatomic and histochemical properties; therefore, they are discussed as groups of muscles rather than individually.
Anatomic location The anterior muscles include the superior and inferior hyoid muscles. The superior hyoid muscles include the geniohyoideus, mylohyoideus, stylohyoideus, hyoglossus, and anterior belly of the digastricus. These muscles arise from various structures superior to the hyoid bone and insert onto the superior aspect of the hyoid bone so that their contraction acts to move the hyoid bone superiorly and anteriorly.18, 19, 34 The inferior muscles include the thyrohyoideus, sternohyoideus, sternothyroideus, and omohyoideus. The thyrohyoideus arises from the thyroid cartilage, the sternohyoideus arises from the clavicle and manubrium, the sternothyroideus arises from the manubrium and upper vertebrae, the omohyoideus arises from the scapula, and all four muscles insert onto the inferior aspect of the hyoid bone.18, 19, 34 The action of these muscles is to pull the hyoid bone and thyroid cartilage inferior and anterior.
Although most of the action of these anterior UES opening muscles is on the hyoid bone, the thyrohyoideus forms the main connection between the hyoid and larynx. Therefore, the simultaneous contraction of the anterior muscles acts to move the hyoid bone and larynx anteriorly. The relative contribution of these anterior muscles to opening of the UES depends on the function of the UES, and the stronger and wider the opening needed, the more of these muscles are recruited.
The posterior muscles include the stylopharyngeus, palatopharyngeus, pterygopharyngeus, and perhaps other superiorly directed posterior pharyngeal muscles.18, 19, 34 The actions of these muscles are to elevate the pharynx and to stabilize the posterior wall of the pharynx by providing tension posteriorly.
Histology and histochemistry The UES opening muscles are histologically and histochemically similar to limb muscles and similar to each other.35, 36, 37, 38, 39, 40, 41 The muscle fibers are oriented in parallel fashion, the fibers are uniform size, and there is little connective tissue. Most of the fibers, about 60% to 85%, are composed of fast-twitch (type II) fibers and have a diameter of 20 to 40 m. The only opening muscle that differs to a significant degree is the thyrohyoid, which has more type I (40%) and highly oxidative muscle fibers41 than the other UES opening muscles. The increased fatigue resistance that results from these types of muscle fibers may contribute to the tonic functions of the thyrohyoid. The thyrohyoid maintains a constant distance between the thyroid and hyoid cartilages during many activities, and tonically contracts to provide the main link between the suprahyoid muscles and the larynx.
Neurophysiology of the Upper Esophageal Sphincter Muscles
Motor Innervation
Cricopharyngeus The CP receives innervation from the pharyngeal plexus, which is supplied by three major nerves: vagus nerve branches including the pharyngeal branch of the vagus nerve [also referred to as the pharyngoesophageal nerve (PEN), superior laryngeal nerve (SLN), and recurrent laryngeal nerve (RLN)]; the glossopharyngeal nerve (GPN); and sympathetic nerve fibers from the superior cervical ganglion. Although many nerves appear to project to the CP, the PEN provides the major motor innervation of the CP.23, 26, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51 Glycogen depletion studies in rats found that the primary motor nerve of the CP is the PEN, and the RLN provides no motor innervation.42 Although SLN stimulation depleted glycogen from the CP,42 electrical stimulation of the PEN resulted in a much greater (about five to six times greater) CP electrical response than stimulation of the SLN26 or GPN,49 and RLN stimulation had no effect on electrical43 or contractile activity46, 47 of the CP. Furthermore, transection of the PEN rather than the GPN50 or SLN46, 47 had profound long-term deficits on swallowing and resting pharyngeal pressure and produced denervation potentials in the CP, and transection of the RLN unilaterally had no effect on resting UES pressure in chronically instrumented dogs.51 In addition, transection of the PEN, but not SLN and RLN bilaterally, blocked the CP EMG responses during swallowing (Figure 4). Moreover, laryngeal paralysis in humans due to RLN paralysis is not associated with UES contractile dysfunction during swallowing.52 Although there is no prominent raphe of the CP, the CP is bilaterally innervated45 with each half of the CP acting as a distinct motor unit.26 The motor end plates of the CP form distinctive bands in each half of the CPh; but both CPo and CPh have end plates scattered throughout the muscle.23 The sympathetic nerves probably innervate the blood vessels and epithelial mucus glands, but they have no role in motor control of the CP.53
Figure 4: Effect of transection of various motor nerves on the motor responses of the UES closing muscles during swallowing.
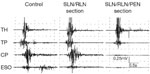
Each panel of this figure depicts the electromyography (EMG) responses of the muscles of the UES during swallowing activated by injection of 1 mL of water into the pharynx of a decerebrate cat. The headings of each panel indicate the nerves that were transected before the stimulus. Note that transection of the superior laryngeal nerve (SLN) and recurrent laryngeal nerve (RLN) bilaterally had no significant effect on the EMG responses of all muscles, but that the additional transection of the pharyngoesophageal nerve (PEN) bilaterally blocked responses of the cricopharyngeus (CP) and esophagus (ESO) 1 cm below CP, but had no significant effect on the thyrohyoideus (TH) or thyropharyngeus (TP).
Some have suggested that the RLN innervates the CP based on studies of the visual tracing of nerve fibers23 or the effects of electrical stimulation of the RLN on electrical responses recorded from the CP.53, 54, 55 The RLN connects with the pharyngeal plexus at the level of CP, and its fibers are continuous with the fibers of this plexus; therefore, it is not possible to know the source of fibers that branch from the RLN nerve and project to the CP based solely on visual inspection of fiber distribution. The electrical stimulation of the RLN causes electrical responses recorded from electrodes on the CP,53, 55 but these responses could have been due to spread of current from the underlying laryngeal muscles. Evidence for the possibility of spread of current from laryngeal muscles was found in these studies,55 as transection of all of the nerves innervating the CP did not eliminate the electrical responses from the CP due to electrical stimulation of the RLN, and RLN stimulation caused electrical responses on the IPC as well, even though prior evidence found no innervation of the IPC by the RLN.23, 28 Spread of current can also explain the results of the other electrical studies,53, 55 as the current needed to cause CP electrical responses to RLN stimulation was much greater than that for PEN stimulation, and the magnitude of the electrical responses recorded from the CP during RLN stimulation were much smaller than those activated by PEN stimulation55 or much smaller55 than that found in other studies when the response originated from the CP itself.26 In none of these electrical recording studies53, 54, 55 of the CP were experiments done to control for the possibility of spread of current from the laryngeal muscles. However, the spread of current has been observed in similar studies. The electrical stimulation of both the PEN and RLN caused electrical responses on the CP, but the electrical response to PEN stimulation was far greater (more than 10-fold) and only PEN stimulation caused contraction of the CP (Figure 5). One source of this current spread may be the posterior cricoarytenoideus, as the posterior cricoarytenoideus lies just below the anterior border of the CP (Figure 1) and the RLN innervates the posterior cricoarytenoideus. The primary location of CP electrical response to RLN stimulation was at the anterior portion of the CP,53 which is the closest part of the CP to the underlying posterior cricoarytenoideus. Therefore, the evidence supporting a role for the RLN in the motor innervation of the CP is lacking.
Figure 5: Effect of electrical nerve stimulation on electrical response and tension of UES closing muscles.

The effects of electrical nerve stimulation on electrical responses are indicated in the left panels and on tension in the right panels, whereas the effects of stimulation of the PEN are indicated in the top panels and of RLN stimulation in the bottom panels. Note that PEN stimulation had a large effect on both electrical response and tension of the CP, whereas RLN stimulation had a small effect on CP electrical response but no effect on CP tension. Therefore, electrical stimulation of the RLN can produce an electrical response on the CP that has no effect on muscle tension and is probably due to spread of electrical current from the RLN or laryngeal muscles underlying the CP.
Inferior pharyngeal constrictor The motor innervation of the IPC is supplied primarily by the pharyngeal branch of the vagus nerve through the pharyngeal plexus23, 28 ; however, no physiologic studies have been reported that have investigated this issue directly. Many studies have attempted to trace nerve fibers through the pharyngeal plexus to the IPC, but because this innervation is through a plexus of interconnecting nerves, it is not possible to definitively determine which branch supplies the IPC using these techniques. The iIPC is supplied by the same branch of the pharyngeal branch of the vagus nerve that supplies the CP.28 This implies similar control mechanisms, but no physiologic studies have been reported to confirm this conjecture. The entire IPC muscle has a single vertical band of motor end plates that corresponds to the location of the pharyngeal plexus.23
Cervical esophagus The cervical esophagus is innervated by the RLN.23, 45, 47 The terminal branches travel around the esophagus in a circular fashion and appear to cross the midline both anteriorly and posteriorly.23
Opening muscles The motor innervation of the UES opening muscles is listed in Table 1.56

Sensory innervation Sensory innervation of the UES involves both epithelial and muscular receptive pathways. The pharyngeal epithelium is innervated by branches of the GPN, SLN, and vagus nerves,57, 58, 59, 60, 61 the laryngeal epithelium by the internal branch of the SLN,60 and the esophageal epithelium by the vagus nerves and the SLN61 or RLN,55 depending on species. The SLN mediates various reflexes from the pharynx and esophagus including swallowing, the RLN mediates the afferent limb of esophago-UES contractile reflex (described below) from the cervical esophagus,47 and the GPN62 mediates the afferent limb of the pharyngo-UES contractile reflex (described below). The loss of sensory function from the UES may explain some of the swallowing deficits observed after transection of the GPN in dogs.49, 50 The CP is devoid of muscle spindles,22, 23 but a Golgi tendon organ-like structure has been found in the human CP that may be involved in providing muscle tension feedback to the pharyngeal motor neurons.63
Most of the UES opening muscles have muscle spindles and the afferent nerves transmitting this proprioceptive information to the central nervous system are the same nerves as the motor nerves.64, 65, 66, 67 The muscles that do not have muscle spindles and this sensory innervation include the digastric muscle,68 stylohyoideus,37 and stylopharyngeus.69, 70
Neurochemical control: Upper esophageal sphincter muscles The only neurotransmitter of any of the UES opening or closing muscles found to mediate contraction is acetylcholine acting through nicotinic cholinergic receptors. However, the following neuropeptides have been found in the CP: neuropeptide Y, calcitonin gene-related peptide (CGRP), tyrosine hydroxylase, substance P, vasoactive intestinal polypeptide, and galanin.71 Calcitonin gene-related peptide has also been found in the TP as well as the striated muscle esophagus, but there is less CGRP in the CP than TP.72, 73 The functional significance of these neuropeptides in the UES muscles is unknown, but they may simply represent the autonomic innervation of the muscle because substance P, VIP, and galanin are found in parasympathetic nerves and CGRP, neuropeptide Y, and tyrosine hydroxylase are found in sympathetic nerves. In general the sympathetic neuropeptides were more abundant than the parasympathetic neuropeptides. Clusters of neurons, that is, ganglia, have been found in the CP rather than TP, and these are mostly parasympathetic ganglia.71 The roles of either the sympathetic or parasympathetic innervation and peptides are unknown, but control of blood vessels, glands, and mucosa are the most likely functions.
Motor neurons: Upper esophageal sphincter closing muscles Most of the motoneurons of the muscles of the UES, that is, the CP, IPC, and cervical esophagus, are located in the nucleus ambiguus, and the nucleus ambiguus is topographically organized such that its more rostral portions contain the motoneurons to the more caudally located muscles,74, 75, 76, 77, 78, 79, 80, 81, 82, 83 although there is considerable overlap. A few motoneurons of the UES muscles are located in various brainstem nuclei outside of the nucleus ambiguus.74, 79, 80 The innervation of the IPC and the CP in those species in which the CP has a median raphe is mostly unilateral.74, 76, 80 The ipsilateral innervation of the pharynx by the nucleus ambiguus is consistent with swallowing studies that found that half of the brainstem controls the ipsilateral half of the pharynx during swallowing.84 In contrast, the reflex activation of the CP requires both halves of the brainstem (I.M. Lang, unpublished observations) and electrical stimulation of the nucleus tractus solitarius (NTS), the primary afferent nucleus of the vagus nerve, caused activation of the CP bilaterally. These results suggest that control of the UES differs with different functions. The neurons of the nucleus ambiguus have extensive dendritic arborization to the adjacent reticular formation,85 and ultrastructural studies indicate the synapses on these neurons are both excitatory and inhibitory.86 These findings provide an anatomic basis for the numerous excitatory and inhibitory responses of the UES and CP. Many pharyngeal motor neurons exhibit respiratory rhythm but none of them had a spontaneous background discharge, suggesting that the pharyngeal motor neurons are not the source of UES tone.87
Motor neurons: Upper esophageal sphincter opening muscles The motor nuclei of the UES opening muscles are listed in Table 1.
Sensory and premotor neurons: Upper esophageal sphincter closing muscles Pharyngeal vagal afferents have their cell bodies in the nodose ganglion74, 78 and terminate on premotor neurons found in the interstitial and intermediate subnuclei of the NTS.88, 89, 90, 91 However stimulation of the pharynx using a physiologic stimulus,92 that is, water injection, activated neurons in the interstitial, intermediate, as well as the ventromedial subnucleus of the NTS. The specific premotor neurons participating in each reflex response initiated by stimulation of the pharynx is unknown.
Esophageal afferents have their cell bodies in the nodose ganglion93, 94, 95 and cervical and thoracic dorsal root ganglia of the spinal cord,93, 94, 96, 97 and the nodose fibers93 terminate in premotor neurons88, 91, 98 in the central subnucleus of the NTS. No studies have been conducted to specifically examine the most rostral portion of the cervical esophagus that forms part of the UES. It is possible that this area of the esophagus has similar distribution of premotor neurons as the pharynx. However, stimulation of the esophagus using a physiologic stimulus92, that is, air injection, activated neurons in not only the central but also the ventral, dorsolateral, and ventrolateral subnuclei of the NTS. It is likely that the additional NTS subnuclei identified in this study are third- and higher order neurons. The function of each NTS subnucleus is unknown. It is possible that muscular and mucosal afferents terminate in different premotor subnuclei because although esophageal distention primarily activates neurons in the central subnucleus of the NTS, esophageal acidification primarily activates neurons in the interstitial subnucleus of the NTS (I.M. Lang, unpublished observations).
The stimulation of the pharynx or esophagus excites numerous reflexes, but the specific premotor neurons associated with each specific reflex response is unknown.
Sensory and premotor neurons: Upper esophageal sphincter opening muscles All of the sensory information from the UES opening muscles is from proprioceptors, and it projects to the associated central nervous system sensory nuclei listed in Table 1.
Functions of the Upper Esophageal Sphincter
Tone Generation
Tone refers to the constant generation of force within a sphincter. It is produced by both passive mechanical properties of the sphincter as well as active muscle contraction, and it is recorded intraluminally by manometric devices or externally by recording EMG from the muscles themselves. Manometric techniques, however, are subject to two problems: (1) they cannot distinguish between passive and active tension, and (2) they create passive tension themselves by distending the highly elastic CP muscle. The main limitation of EMG activity is that it can provide an index of active but not passive tension. Another important consideration is that the UES is composed of striated muscles that, unlike smooth muscles, have no intrinsic tone-generating mechanisms. All tone of the UES must arise from activation of the motor neurons.
Manometric studies have found a wide range, 35 to 200 mmHg, of resting UES pressure,99, 100 and this pressure falls to very low levels during sleep14 or anesthesia.16 In addition, pharyngeal motoneurons of decerebrate and paralyzed cats do not exhibit a spontaneous discharge.87 Therefore, evidence suggests that the UES has no active basal tone, although a residual intraluminal pressure may be recorded manometrically due to the passive elastic properties of the CP.
Although there is no basal tone of the UES, the tone of the UES is highly variable and can increase to very high levels. High UES pressures have been recorded during acute stress15, 101 (Figure 6) and other emotional states,1 and sharp increases in UES pressure have been recorded during waking.14 In addition, large increases in ongoing EMG activity of the CP in animals1 have been recorded during changes in posture, in certain head and body positions, and in response to stress or excitation (Figure 7). Therefore, although the UES has no active basal tone, there are many mechanisms for increasing tone above basal levels, and many of them involve reflexes, as discussed below.
Figure 6: Effect of stress on UES pressure.

This figure depicts the tracings of UES pressure, skin conductance, integrated frontalis EMG, and heart rate before and during a stressful listening task (at arrow). Note the anticipatory rise in UES pressure about 15 s before the stress begins. (Source: Cook et al.,15 with permission of the American Gastroenterological Association.)
Figure 7: Relationship of CP and TP EMG activities to UES pressure during rest and excitation.
![Figure 7 : Relationship of CP and TP|[emsp]|EMG activities to UES pressure during rest and excitation. Unfortunately we are unable to provide accessible alternative text for this. If you require assistance to access this image, or to obtain a text description, please contact npg@nature.com](/gimo/contents/pt1/thumbs/gimo12-f7.jpg)
a: Actual responses. b: Graph of the relationship between EMG and UES pressure. The UES pressure changed five times: one swallow (S) preceded and followed by two rises in UES pressure induced by whistling in an awake dog lying on its side. Note that UES pressure changes correlated well (p<.05) with integrated CP-EMG but not integrated TP-EMG. TP, thyropharyngeus: CP, cricopharyngeus. (Source: Lang et al.,1 with permission of the American Physiological Society.)
The specific muscles involved in active UES tone generation have been investigated by recording EMG from the TP, CP, and proximal esophagus in conscious, nonsedated, and nonrestrained animals. Under these conditions it was observed1 that the TP EMG changed little regardless of excitability, arousal, and head position, whereas the CP EMG changed greatly (Figure 7). In addition, correlation of basal TP EMG and CP EMG with UES pressure during changes in excitability1 revealed that CP but not TP EMG was significantly related to UES pressure (Figure 7). Yet, even in these studies it was found that peak pressure of the UEHPZ was observed radiographically at the distal end of the TP or proximal end of the CP.1 However, as with similar human studies these measurements were made with the animal relaxed and its head supported. Only under these conditions is TP tone more active than CP tone (Figure 8). Therefore, although the UEHPZ may occur in or close to the TP based on published studies, this finding only applies to the unusual conditions of the recording session and probably does not reflect the situation in a freely active individual. The UEHPZ may be higher in the vicinity of the TP under very low states of CP activation due to differences in passive mechanical factors of the muscles as the CP is much more compliant than the TP due to its high content of elastic tissue.
Figure 8: High variability of CP but not TP EMG.
![Figure 8 : High variability of CP but not TP|[emsp]|EMG. Unfortunately we are unable to provide accessible alternative text for this. If you require assistance to access this image, or to obtain a text description, please contact npg@nature.com](/gimo/contents/pt1/thumbs/gimo12-f8.jpg)
This figure depicts the EMG responses of the TP, CP, and diaphragm in a minimally restrained awake chronically instrumented dog. Throughout this recording the dog is lying prone with its head upright or resting down on its paws. Note that the CP but not TP EMG varies with head position, and when the dog is relaxed at rest with its head down the TP EMG exceeds the CP EMG. TP, thyropharyngeus; CP, cricopharyngeus; Dia-D, diaphragmatic dome fibers.
The above studies recorded TP EMG 1 to 2 cm above the CP in dogs, but recent evidence has suggested that the iIPC of humans has the similar histochemical properties and innervation pattern as the CP,28 and therefore may also participate in the UEHPZ and tone generation. However, no physiologic studies have been reported to date to confirm this conjecture.
Upper Esophageal Sphincter Opening and Closing
The UES opens during various physiologic states, for example, swallowing, vomiting, and belching, to allow the passage of luminal contents, but the specific manner of the opening and the specific muscles involved differ with the physiologic state. In all states UES opening begins with UES relaxation followed briefly by contraction of UES distracting muscles, but the specific responses differ as discussed below.
Swallowing
Opening of the UES during swallowing is a very complex maneuver. Simultaneous recordings of UES pressure and opening with EMG recording of UES muscles in chronically instrumented dogs found the following sequence of events during swallowing.1 First, the CP muscle relaxed; about 100 milliseconds (ms) later the UES pressure began to fall, that is, UES relaxation, and this was followed about 100 ms (150 ms in humans3) later by UES opening (Figures 9 and 10). Videofluoroscopic studies found that UES opening was associated with superior and anterior movement of the hyoid and larynx (Figure 10).1, 3 The onset of the superior movement of the hyoid and larynx coincided with the beginning of UES relaxation and preceded anterior movement of the larynx and hyoid by about 100 ms in dogs1 and over 200 ms in humans3 (Figure 10). The anterior movement just preceded (humans,3Figure 10) or was concomitant (dogs1) with UES opening. Therefore, the UES is first moved superiorly perhaps in order to position the UES to take maximum advantage of the subsequent contraction of the UES opening muscles. This sequential movement of hyoid superiorly and anteriorly probably accounts for the elliptical pattern of movement of the hyoid bone (Figure 11) observed during swallowing.106
Figure 9: Temporal relationship of UES pressure, opening and CP EMG during 4-mL barium swallow relative to hyoid movement.
![Figure 9 : Temporal relationship of UES pressure, opening and CP|[emsp]|EMG during 4-mL barium swallow relative to hyoid movement. Unfortunately we are unable to provide accessible alternative text for this. If you require assistance to access this image, or to obtain a text description, please contact npg@nature.com](/gimo/contents/pt1/thumbs/gimo12-f9.jpg)
The start and end of hyoid movement are indicated by vertical lines where the shaded areas indicate standard error (SE). Horizontal bars depict mean + SE onset and offset of measured variables relative to start of hyoid movement. Green bars depict data from noninstrumented dogs (n = 8), and blue bars depict data from instrumented dogs (n = 6). Note the electrode implantation significantly delayed time of UES closure. There is approximately a 100-ms difference between CP relaxation, UES relaxation, and UES opening. (Source: Lang et al.,1 with permission of the American Physiological Society.)
Figure 10: Temporal relationship among UES pressure, trans-UES flow, and movement of the hyoid and larynx during 5-mL barium swallow.

The data are pooled from eight subjects. The brown bar indicates the period of UES opening when trans-UES flow of barium occurred. Note that the superior movement of the larynx and hyoid precede the drop in UES pressure and the anterior movement of the larynx and hyoid precede UES opening. (Source: Cook et al.,3 with permission of the American Physiological Society.)
Figure 11: Movement of the hyoid bone during belching (a) and swallowing (b).

Open circles indicate UES opening observed by videofluoroscopy. Although hyoid bone movement during swallowing was invariably upward, forward, and counterclockwise, its movement during belching was mainly anterior and clockwise. The magnitude of hyoid bone movement during belching was significantly less than its movement during swallowing. Hyoid bone movement is an indication of the magnitude of the distraction forces that open the UES (p <.02). (Source: Shaker et al.,106 with permission of the American Physiological Society.)
The CP is the primary muscle that relaxes to permit UES opening while the TP muscle contracts during this time1, 102, 103 (Figure 12). During swallowing the CP relaxes for about 0.5 s in humans3 and about 0.3 s in dogs.1, 102, 103 Superior movement of the larynx and hyoid is probably caused by contraction of the superior hyoid muscles, as all of these muscles begin to be activated concomitant with CP relaxation, and this activation continues throughout CP relaxation.1, 103 It is also important to note that the inferior hyoid muscles completely relax during this time (Figure 13), allowing maximum superior movement of the hyoid, larynx and UES.1, 103 Anterior movement of the hyoid and larynx corresponds with maximal activation of the geniohyoideus, which suggests that the geniohyoideus is one of the primary suprahyoid muscles responsible for UES opening.1, 103 The stylopharyngeus is activated concomitant with the geniohyoideus; therefore, it is likely that maximum opening of the UES is due to activation not only of the geniohyoideus but also of the stylopharyngeus and other posterior pharyngeal muscles that act to stabilize the posterior wall of the pharynx.103
Figure 12: Response of UES muscles during swallowing.
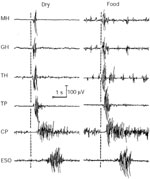
This figure illustrates the EMG responses of the three UES closure muscles, and many of the superior hyoid UES opening muscles during a dry swallow ad lib feeding of canned food in a chronically instrumented dog. Note that the three UES closing muscles respond quite differently during swallowing but the superior opening muscles are activate almost simultaneously. MH, mylohyoideus; GH, geniohyoideus; TH, thyrohyoideus; TP, thyropharyngeus; CP, cricopharyngeus; ESO, esophagus 2 cm below CP.
Figure 13: Responses of superior and inferior hyoid muscles during swallowing.
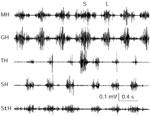
This figure illustrates the EMG responses of the superior (MH, GH) and inferior (TH, SH, StH) hyoid muscles during swallowing in a chronically instrumented dog. These muscles are capable of putting anterior traction on the larynx, thereby opening the UES. Note that during swallowing only the superior hyoid muscles are activated. S, swallowing: L, licking; MH, mylohyoideus; GH, geniohyoideus; TH, thyrohyoideus; SH, sternohyoideus; StH, sternothyroideus.
Belching
Belching is characterized by a prolonged, over 1 s, drop in UES pressure,105, 106 opening of the UES105, 106 (Figure 14), and relaxation of the UES muscles91 (Figures 15 and 16). Unlike during swallowing, both the TP and CP relax during belching (Figure 16), and the UES pressure drop during belching coincides with rather than precedes UES muscle relaxation (I.M. Lang, unpublished observations). In addition, during belching most of the distracting muscles are not active and the one active distracting muscle, the thyrohyoideus, is activated at a low but constant level107 (Figure 15). These small changes in UES distracting muscles probably account for the rather short anterior movement and lack of superior movement of the hyoid bone observed during belching105 (Figure 11). The role of infrahyoid and posterior pharyngeal muscles during belching remain to be studied, but given the small response of the anterior distracting muscles (and small anterior movement of the hyoid105), it is suggested that UES opening during belching may be related more to the distracting effects of the bolus than occurs during other functions such as swallowing. This effect may account for the observation that the delay from UES pressure drop to UES opening is longer during belching (about 0.4 s106) than swallowing (about 0.2 s3).
Figure 14: Temporal relationship among function of the glottis, UES, hyoid bone, and esophageal and stomach pressures during belching induced by rapid injection of 40 mL of air into the esophagus.
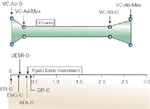
This figure is based on data obtained from concurrent recordings of videoendoscopic, videofluoroscopic, manometric, and electromyographic data. Belch begins with vocal cord adduction and ends with return to resting position. All other events, including UES opening and closure, occur while cords are fully adducted and thereby introitus to trachea is closed. VC-Ad-O, onset of vocal cord adduction; VC-Ab-Max, maximum onset of vocal cord adduction; VC-Ab-O, onset of vocal cord abduction; VC-Ab-Max, return of vocal cords to resting position; EPI-O, onset of increase in intraesophageal pressure; AEA-O, onset of approximation of arytenoids toward base of epiglottis; GPI-O, onset of increase in intragastric pressure; EMG-O, onset of EMG response from geniohyoid/mylohyoid muscle group; UESR-O, onset of UES relaxation. (Source: Shaker et al.,106 with permission of the American Physiological Society.)
Figure 15: Electromyography (EMG) responses of the opening and closing muscles of the UES during belching and swallowing activated by injection of 100 mL of air into the stomach of a chronically instrumented dog.

Note that the only anteriorly directed UES opening muscle activated during belching is the TH and the main UES opening muscle activated is the posteriorly directed StP. These muscle responses are consistent with the movement of the hyoid during belching (90). StP, stylopharyngeus, GH, geniohyoideus; TH, thyrohyoideus; CP, cricopharyngeus; ESO, esophagus 1 cm below the CP; SH, sternohyoideus.
Figure 16: Effects of belching on the UES closure muscles.

Belching was activated by the injection of air (50 mL) into the stomach of a chronically instrumented dog. Note that belching is associated with inhibition of both the TP and CP during the belch. TP, thyropharyngeus, CP, cricopharyngeus; ESO, esophagus 1 cm below CP; L, longitudinally oriented electrodes; C, circularly oriented electrodes.
Retching and Vomiting
The UES is involved in numerous functions during retching and vomiting, and all three UES muscles participate in one or more of these responses (Figure 17).
Figure 17: Role of UES closure and opening muscles during the three phases of vomiting.

Schematic illustration of typical pattern of activation of most pharyngeal, hyoid, and laryngeal muscles during vomiting derived from EMG data of chronically instrumented dogs stimulated to vomit by apomorphine. Note that during the pre-retch phase the tone of the CP and esophagus increase, significantly closing the UES. During retching the muscles pulling the UES open in a superior direction (superior hyoid muscles, HP and SP) are all activated in concert 180 degrees out of phase with the UES opening muscles that open the UES in an inferior direction (inferior hyoid muscles). However, the UES does not open during superior movement because all of the UES closure muscles are activated as well. During vomiting, the superior and inferior hyoid and pharyngeal UES opening muscles are activated concomitantly with relaxation of all of the UES closure muscles, thereby causing maximum opening of the UES. MH, mylohyoideus; GH, geniohyoideus; TH, thyrohyoideus; HG, hyoglossus; HP, hyopharyngeus; SP, stylopharyngeus; TP, thyropharyngeus; CP, cricopharyngeus; E#, esophagus number of centimeters from UES; SH, stenohyoideus; ST, sternothyroideus; CT, cricothyroideus; TA, thyroarytenoideus; CD, cricoarytenoideus dorsalis; DH, diaphragm hiatus. (Source: Lang et al.,103 with permission of the American Physiological Society.)
Pre-retch tone increase Just prior to retching the tone of the UES increases greatly in phase with the diaphragm, and this increase in tone involves the CP and cervical esophagus, but not the TP.103 The function of this increase in tone is to prevent air insufflation of the esophagus during the large negative thoracic pressure that is subsequently generated during the first very large inspiration of the first retch. This closing of the UES allows the lungs to fully inflate without allowing air into the esophagus or stomach. The LES is relaxed at this time, so without UES closure the stomach would fill with air.
Phasic changes in UES tone during retching After the first very large retch to fill the lungs, the diaphragm and glottis then shift phase so that the diaphragm contracts during glottal closure instead of glottal opening.103 This phase shift allows the diaphragm to be used as a pump to expel gastric contents, and the external intercostal muscles then become the sole generator of inspiration. Thus, at the end of the inspiratory phase of the first retch, the glottis closes as the UES opens, shifting the force of the diaphragmatic contraction to the stomach. The diaphragm then relaxes, the glottis opens, and the UES closes very strongly.103 The function of this closure of the UES during this phase of retching is to prevent aspiration, as this occurs during inspiration and gastric contents may be in the upper esophagus at this time.104 Prior studies have found that gastric contents can be expelled to just below the UES during retching104. This cycle continues until the end of retching. The UES during this phase of retching involves all three UES muscles, that is, cervical esophagus, CP, and TP.
Vomitus expulsion During vomitus expulsion all of the UES muscles relax and the UES is pulled open by contraction of not only the suprahyoid muscles but also the infrahyoid muscles as well as the posterior pharyngeal muscles.103 Therefore, maximum superior traction occurs as well as maximum anterior to posterior opening tension. The primary UES opening muscles during vomitus expulsion are the mylohyoideus, geniohyoideus, sternohyoideus, and stylopharyngeus.103 Although the trajectory of the hyoid bone has not been determined during vomitus expulsion, based on the muscle activation sequence (stylopharyngeus and hyopharyngeus, then mylohyoideus and geniohyoideus, then sternohyoideus) the hyoid bone probably forms an elliptical path moving in the following sequence: posteriorly, superiorly, anteriorly, inferiorly, and back to rest position. This movement pattern would be very different from that during swallowing or belching.
Upper Esophageal Sphincter Reflexes
Pharyngo-UES Contractile Reflex
Stimulation of the pharynx with light touch increases tone in the UES,108 and this response is due to activation of the TP and CP but primarily the CP.62 In some experiments the TP was found to be involved in the response but other times it was not. It is unknown whether the cervical esophagus is involved in this reflex response. The most sensitive area of the pharynx was the hypopharynx and nasopharynx.62 The afferent limb of this reflex is the glossopharyngeal nerve, and the efferent limb is the pharyngeal branch of the vagus nerve.62 This reflex may function to prevent air insufflation of the esophagus during strong inspiration as this reflex can be activated by air puffs and is strongest closest to the nasopharynx.108
Esophago-UES Contractile Reflex
The slow distention of the esophagus causes an increase in tone of the UES or CP,111, 112, 113 and this reflex involves all three UES muscles—CP, TP, and cervical esophagus—but primarily the CP (Figure 18). This reflex can be activated at all levels of the esophagus, but is most sensitive near the UES, and the receptors for this reflex are probably slowly adapting muscular mechanoreceptors.112 This reflex is stimulated not only by distention but also by contraction of the esophageal wall, and it is strongly activated by the propagating esophageal peristalsis.112 The role of intraluminal acid in activating this reflex is equivocal, as results have been contradictory.114, 115, 116, 117 The afferent limb of this reflex is the vagus nerve,109, 110, 112 and the efferent limb is the pharyngeal branch of the vagus nerve (I.M. Lang, unpublished observation). This reflex probably functions to prevent reflux of a bolus during its aborad propagation in esophageal peristalsis.
Figure 18: Effect of esophago-UES contractile reflex on the UES closure muscles.
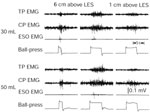
Distention of the esophagus of a chronically instrumented awake dog with a balloon caused an increase in EMG activity of the TP, CP, and cervical esophagus, but primarily the CP. The response magnitude was related to the magnitude of the stimulus, and the response was greater closer to the UES. TP, thyropharyngeus; CP, cricopharyngeus; ESO, esophagus 1 cm below CP; ball-press, balloon pressure; LES, lower esophageal sphincter.
Esophago-UES Relaxation Reflex
The rapid distention of the esophagus causes relaxation of the UES118, 119 (Figure 19) that involves the relaxation of the CP.112 It is unknown whether the TP or cervical esophagus are involved in this reflex, but it is likely that they are. This reflex is probably part of the belch response, as it is activated by the same stimuli, and the pharyngeal and laryngeal motor responses that accompany this reflex are identical to those activated during belching as discussed previously.112 The receptors for this reflex are probably rapidly adapting mucosal mechanoreceptors, and the afferent limb of this reflex is the vagus nerve.112
Figure 19: Role of UES closure muscles in esophago-UES relaxation reflex.
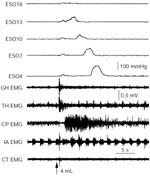
Rapid injection of air into the esophagus of a decerebrate cat caused the CP to relax concomitant with activation of UES opening muscles, i.e., GH and TH. ESO#, esophagus number of centimeters from LES; GH, geniohyoideus; TH, thyrohyoideus; CP, cricopharyngeus; IA, intraarytenoideus; CT, cricothyroideus. (Source: Lang et al.,112 with permission of the American Physiological Society.)
Lung-UES Contractile Reflex
The distention of the lungs above tidal volume or the rapid deflation of the lungs causes an increase in tone of the UES that is probably mediated by contraction of the TP and CP but primarily the CP (I.M. Lang, unpublished observations). The receptors for this reflex are probably rapidly adapting lung inflation receptors, and the afferent limb of this reflex is the vagus nerve (I.M. Lang, unpublished observations). This reflex may be partly responsible for the respiratory rhythm (Figure 20) that is often found on the UES1, 16 and this reflex may function to prevent esophageal insufflation during deep inspirations.
Vestibulo-UES Contractile Reflex
The UES tone changes greatly with change in head position, and this tone change is due to activation of both the TP and CP but primarily the CP1, 16 (Figure 8). It is likely that this reflex is due to activation of the vestibular apparatus and may function to prevent pharyngeal reflux of esophageal contents when gravity is against the direction of peristalsis.
Conclusion
The UES is not just one muscle but is composed of sets of opening and closing muscles; however, the primary muscle that governs all functions of the UES is the CP. The CP is not only unique in its actions, but also in its anatomy and physiology. Numerous nerves control the function of the UES, but the primary motor nerve of the CP is the pharyngeal branch of the vagus nerve. Tone is generated in the UES by various reflexes rather than specific tone-generating neurocircuitry. The primary functions of the UES are to prevent esophageal air insufflation during negative intrathoracic pressure and to prevent esophagopharyngeal/laryngeal reflux during esophageal peristalsis. The degree to which the UES is actively opened by distracting muscles is related to the volume of the bolus such that belching is associated with minimal and vomiting with maximal distracting forces.
In the past most research investigations have concentrated on the role of the cricopharyngeus in UES function, but in future studies the role of other sphincteric and distracting muscles involved with UES function will be investigated. In addition, future studies will investigate the possibility that individual UES muscles may have multiple functions. Better methods for recording EMG or force, possibly using magnetic resonance imaging techniques, from UES muscles in humans would greatly increase our knowledge of the role of the individual UES muscles in various functions.
