
Abstract
The oviduct functions in the transportation of gametes to the site of fertilization (the ampulla) and is the site of early embryonic development. Alterations of this early developmental environment, such as the presence of sexually transmitted pathogens, may affect oviduct function leading to reduced fertilization rates and contribute to compromised embryonic development. In this study, sperm interactions, particle transport speed (PTS) and cilia beat frequency (CBF) in the ampulla following exposure to lipopolysaccharide (LPS), a constituent of the sexually transmitted pathogens Chlamydia trachomatis and Chlamydia abortus, was investigated. Three complementary experiments were performed to analyse; (1) bound sperm motility and cilia function (2) transport velocity in the oviduct and (3) the expression of genes related to immune function and inflammatory response (CASP3, CD14, MYD88, TLR4 and TRAF6). The motility of bound sperm was significantly lower in ampullae that were exposed to LPS. CBF and PTS significantly increased after treatment with LPS for 2 hours. Finally, gene expression analysis revealed that CASP3 and CD14 were significantly upregulated and TLR4 trended towards increased expression following treatment with LPS. These findings provide an insight on the impact of LPS on the oviduct sperm interaction, and have implications for both male and female fertility.
Similar content being viewed by others


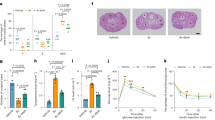
Introduction
Chlamydia trachomatis (C. trachomatis) is the most frequently sexually transmitted human disease worldwide1. C. trachomatis can cause infection of the upper genital tract without any symptoms and can lead to damage of the oviduct (uterine tubes, Fallopian tubes), increasing the risk of future ectopic pregnancy and tubal infertility; consequently C. trachomatis is estimated to be the most costly nonviral sexually transmitted infection. In ruminants, Chlamydia abortus (C. abortus) is endemic worldwide and is a major causative agent associated with abortion and foetal loss in sheep and cattle2,3. In addition, a large proportion, 80–90%, of bovine uteri are contaminated with bacteria following parturition, which may persist leading to subclinical endometritis and reduced conception rates4,5,6. It is recognised that infection of the endometrium by pathogenic bacteria and viruses is preceded by endometrial infection with Escherichia coli (E. coli)7. This large scale postpartum infection has vast economic consequence, contributing to female reproductive disorders and the associated infertility which is estimated to cost €1.4 billion in the EU and $650 million per year in the United States7,8,9. In human, aberrant immune function in relation to infection is associated with adverse outcomes in many aspects of fecundity, such as ovarian function, embryo implantation and pregnancy loss10,11.
For successful fertilization sperm must first migrate through the cervix, transit through the uterus, enter the narrow passageway into the oviduct (the uterotubal junction) and swim against the tubal fluid flow before reaching the oocyte in the ampulla. The ampulla is the middle region of the three anatomical segments into which the oviduct is divided: the infundibulum (proximal to the ovary), ampulla and isthmus (proximal to the uterotubal junction and uterine horn). It has a central role in the early stages of development as it is the location where gametes undergo their final maturation, fertilization occurs and early embryonic development takes place12,13.
Over 20 years ago it was suggested that binding of bovine sperm to oviduct epithelial cells prolongs sperm motility in order to maintain fertilisation capacity14. These findings were confirmed by further studies showing that sperm are retained in the isthmus in a storage reservoir15, which is essential for preserving sperm fertility, as sperm remain bound until they are slowly released around the time of ovulation16. Additionally, it has also been demonstrated that sperm binding is not affected by the anatomical region of the oviduct or the cycle stage of heifers17. More recently, it has been demonstrated that in vitro incubation of human sperm with known pathogenic (E. coli), and conditionally pathogenic bacterial strains (Staphylococcus haemolyticus, Bacteroides ureolyticus) results in cellular death, which may be due to both apoptosis and necrosis18. The authors concluded that bacteriospermia may be a direct cause of subfertility or contribute to additional negative factors that may deteriorate the prognosis of fertility in natural and assisted reproduction. However, a previous study investigating the incidence of bacteriospermia and elevated seminal leukocytes in a cohort of subfertile Canadian males reported that, although prevalent in subfertile men, bacteriospermia was not statistically associated with subfertility19. Nevertheless, these studies highlight the importance for further interrogation of the relationship between infection and fertility.
Sperm viability is not the only condition for successful reproduction; the biophysical environment of the female genital tract, such as transport speed and cilia beat frequency, plays a crucial role to facilitate sperm migration into the oviduct and to block the access of sexually transmitted pathogens20. In spite of these defensive mechanisms, infection of the oviduct is still possible as epithelial cells in the oviduct are exposed to pathogens and endotoxins from the uterus and peritoneal cavity21,22.
Lipopolysaccharide (LPS) is a major constituent of the cell wall in most gram negative bacteria. Gram-negative bacteria (e.g. C. trachomatis, C .abortus and E. coli) signify a major class of pathogens that are thought to infect all eukaryotes23. Therefore, LPS is a useful molecule for mimicking bacterial infections, both in vitro and ex vivo. Stimulation of TLR4, a member of the Toll-like receptor family expressed on the cell surface and endosomes of monocytes, macrophages, dendritic cells, the oviduct and intestinal epithelium, with the bacterial endotoxin LPS induces intracellular signalling (involving TRAF6 and MYD88) that results in the release of critical proinflammatory cytokines, which are required for triggering effective immune responses22,24,25,26,27. Transcriptomic effects, following challenge with LPS, have previously been investigated in vitro using epithelial cells of the bovine oviduct and endometrium5,28. These studies demonstrated an upregulation of inflammatory response genes (e.g. TRAF6, and CASP3) and immune and stress response genes (TLR4, MYD88 and CD14). However the response of the oviduct to LPS exposure, ex vivo, remains unknown.
The objective of the current investigation was to develop an ex vivo model of infection for studying sperm interactions in the oviduct. To this end, we developed a method using explants of the bovine oviduct and LPS to elucidate the physiological and molecular changes elicited by LPS on oviduct function and sperm-oviduct interactions. In respect of that, we combined previously established video microscopy imaging techniques29 with gene expression analysis to determine the effect of an LPS challenge on (1) bound sperm motility, (2) particle transport speed, (3) cilia beat frequency and (4) abundance of inflammatory, immune and stress response genes.
Results
Exposure of ampulla to LPS alters sperm motility
In the first experiment, uncapacitated sperm that were bound to sections of ampulla, which had been pre-incubated with 10 μg/ml LPS, were assessed for motility (Fig. 1). After addition of frozen thawed spermatozoa of bulls with proven fertility, it was observed that sperm rapidly attached to ciliated epithelial cells of the ampulla sections. Subsequently, the buffer was replaced after 30 and 60 min to remove unbound sperm and to ensure that sperm being captured at 90 min were bound. The motility of sperm, which were bound to the ampulla, was recorded in 5 defined regions of each video captured (Fig. 2Ai). Sperm were scored as either being motile or immotile and percentage motility was determined by comparing the ratio of bound motile to bound immotile sperm (Fig. 2Aii). The total number of individual bound sperm assessed under control and LPS challenge conditions were, CTL = 249 and LPS = 157. Although it appeared that less sperm were bound following exposure of the ampulla to LPS the difference was not statistically significant (P = 0.16). However, analysis of bound sperm motility revealed that it was significantly compromised in the LPS group (P < 0.05) with an average motility of 27%, about half the motility (58%) of bound sperm in the control group (Fig. 2B). Representative videos of sperm motility with and without LPS-challenge are located in the supplementary information (Supplementary videos 1 and 2).

Oviducts were isolated from slaughterhouse reproductive tracts (A) that were determined to be in the diestrus stage of the bovine reproductive cycle. Sections of the ampulla (B) were pinned or not pinned according to experiment being carried out. L = left ovary and R = right ovary. Black arrows show the location of the left and right uterine horns and the green box highlights the position of the right oviduct. With the exception of the cow schematic, this figure was drawn by AOD using photographs captured by MDF. The cow was adapted from the PPT Drawing Toolkits BIOLOGY Bundle, Motifolio // www.motifolio.com/.

Video microscopy was used to capture spermatozoa bound to the epithelium in sections of the bovine ampulla. (A) The motility of bound sperm was determined in 5 regions (represented by red boxes in (i)) using ImagePro software, bound sperm that were motile were labelled with a square and bound sperm that were immotile with a circle (ii). (B) The number of motile sperm (represented as a percentage of the total number of bound sperm counted) was significantly reduced following exposure to LPS (P < 0.05).
Increased particle transport speed following LPS challenge
Transit of gametes and early embryos through the oviduct is likely to be affected by the tubal fluid flow, which is managed by a) ciliary activity and b) smooth muscle contraction. In order to gain an insight into the physiological impact of LPS on oviduct function, transport speed was investigated in a second series of experiments (Figs 1 and 3A). Sections of ampulla that were exposed to 10 ug/ml LPS (n = 5) for 2 h were compared to control sections that were maintained in HEPES buffer (CTL n = 5). Six videos were recorded for all samples giving a total number of 30 for both CTL and LPS (Supplementary videos 3 and 4). After this relatively short exposure to LPS, the transport speed was significantly increased in LPS-exposed oviducts, CTL = 127 ± 5.5 vs LPS = 181.8 ± 7.2 μm/sec (mean ± SEM) (P ≤ 0.001) (Fig. 3B). A set of complementary PTS experiments were carried out, using ampulla sections from tracts similar to those described in supplementary file 1, to elucidate the directionality of tubal flow in the bovine oviduct (3 AOIs from two biological replicates, supplementary videos 5 and 6). These experiments demonstrated that particles transit in the direction of the uterus. Further investigation of videos recorded for PTS analysis revealed that the flow was in the same direction for each AOI in each replicate. Taken together, the direction of particle transport observed in both experiments suggests that tubal flow is unidirectional in these samples and is in accordance with what has previously been reported for orientation of tubal flow during diestrus, i.e. in the direction of the uterus30,31.

(A) Representative images of the method used to determine particle transport speed. (i) Raw image showing the presence of the particles (2.8 μM Dynabeads) in the oviduct. (ii) Removal of background and labelling of particles (red) Using ImagePro. (iii) Tracking of particles. (B) Particle transport speed (PTS) was significantly increased in ampulla sections that had been incubated in the presence of LPS for 2 h (P < 0.05). Data are presented as the mean ± standard deviation. Closed black circles represent the average PTS for each individual video used in the analysis (Total number of oviducts analysed, n = 5, Total number of PTS measurements, CTL n = 30 and LPS n = 29).
Does LPS affect cilia function?
Subsequent to identifying that transport speed was significantly increased after treatment with LPS we hypothesised that increased PTS occurs as a result of elevated cilia beat frequency. To test this hypothesis we recorded videos in oviducts that were either (1) maintained in HEPES buffer (CTL) or (2) exposed to LPS. CBF was analysed for both groups, before and after 2 h incubation at 37 °C (Fig. 1). Videos were captured using the same method described for analysis of sperm (Fig. 2Ai). The method used to determine CBF is outlined in Fig. 4A (See supplementary videos 7 and 8 for representative videos). As expected, no significant differences were observed for CBF between the CTL and LPS samples at the beginning of the experiment prior to incubation (CTL-before = 17.5 ± 4.8 Hz vs LPS-before = 17.6 ± 4.6 Hz, P ≥ 0.05) (Fig. 4B). However, samples that were incubated in LPS for 2 h showed a significant increase in CBF, relative to those that were unexposed CTL-after = 20.1 ± 5.2 Hz vs LPS-after = 23.1 ± 5.1 Hz, P ≤ 0.05) (Fig. 4B). These observations are in agreement with what was observed for particle transport in response to LPS exposure. Additionally, in this model there appeared to be an increase in CBF frequency over time in both control and LPS-treated samples (P ≤ 0.05).

Cilia beat frequency (CBF) was analysed in the ampulla before and after exposure to LPS and compared to the CBF of ampulla sections that were unexposed (CTL) using ImageJ software. (A)(i) Videos were captured at 100 frames per second and single ciliated cells were selected (yellow boxes) as areas of interest (AOI) before a binary mask was applied using the convert to binary function (ii). The AOIs are represented by red boxes following masking. (B) No significant differences in CBF were observed in CBF before LPS treatment (P > 0.05). CBF was significantly higher in the LPS group after incubation with LPS for 2 h (P > 0.05). CTL and LPS before videos n = 25 each. CTL and LPS after videos n = 25 each. Total number of AOIs analysed; CTL-before n = 100, LPS-before n = 100, CTL-after n = 100 and LPS-after n = 100.
Effect of ex vivo LPS exposure on expression of inflammatory and immunological response genes
Gene expression analysis was performed to determine if LPS was exerting an effect on the epithelial cells of ex vivo sections of ampulla, relative to untreated control samples (Fig. 1). The gene expression profiles of a set of five target genes (CASP3, CD14, MYD88, TLR4 and TRAF6), which have been shown to be up regulated in inflammatory, immunological and stress responses in vitro28, were characterised using three separate, but complementary, methods. No significant changes (P > 0.05) in the expression of any of the target genes was observed in epithelial cells isolated from closed sections of oviduct that were exposed to 10 μg/ml of LPS for a period of 2 h (Fig. 5A). Therefore, expression of the target genes was assessed in a second set of samples in which the section of ampulla had been dissected longitudinally through the lumen prior to LPS exposure. In this experiment the expression of CASP3 was significantly higher (P < 0.05) in ampulla that was challenged with LPS group (Fig. 5B). In a final set of experiments the concentration of LPS was increased from 10 μg/ml to 100 μg/ml and the incubation time increased to 3 h. Again, CASP3 expression was significantly higher (P < 0.05) in the ampulla sections exposed to LPS (Fig. 5C). Additionally, CD14 expression was also higher (P < 0.05) in the LPS challenged sections and TLR4 had trended towards increased expression (Fig. 5C).

Calibrated normalised relative quantities (CNRQ) were plotted using the boxplot function in Mintab v16. Whiskers and boxes represent outliers and the interquartile range, respectively. No significant differences (P > 0.05) in expression were observed for any of the genes in epithelial cells isolated from intact sections of oviduct (A). Only CASP3 was significantly upregulated (P < 0.05) in epithelial cells isolated from longitudinally dissected pieces of oviduct exposed to 10 μg/ml LPS (B) however both CASP3 and CD14 were significantly upregulated when the concentration was increased to 100 μg/ml (C).
Discussion
The effects of clinical and sub-clinical endometritis on fertility have been extensively studied using the bovine model5. However, limited information is known about the impact of infection in the bovine oviduct on fertility. The current investigations provide novel information on the impact of LPS, a key constituent of E. coli and the sexually transmitted pathogens C. trachomatis and C. abortus, on bound sperm motility, transport speed, cilia function and inflammatory and immunological gene expression patterns in an ex vivo model of oviduct infection.
The elevated speed and frequency observed in particle transport and cilia beating, respectively, under LPS challenge may represent a host response mechanism to increase fluid flow during the early stages of infection, in which the oviduct is attempting to remain sterile by flushing foreign particles towards the uterus20. This hypothesis is supported by previous evidence demonstrating that the fluid flow is unidirectional30,31, and that it represents a defensive mechanism against the invasion of pathogens13. Indeed, elevated CBF during infection is not uncommon as it has been demonstrated that cilia beat frequency is increased in murine and human ex vivo mucus-free airway preparations, when challenged with LPS32. In the current study, particles were shown to move in the direction of the uterus, and LPS was shown to increase both CBF and PTS. Although LPS clearly has an impact on the oviduct through increasing cilia beating and transport speed, the consequences of LPS on sperm transit through these conditions, in vivo, remains less clear. Indeed, it has been demonstrated that sperm are capable of swimming against the oviductal fluid flow (a process known as positive rheotaxis)33. Additionally, using microfluidic modelling it has been shown that microgrooves and gentle fluid flows provide preferential pathways that facilitate sperm transport against fluid flow20. Therefore, it is possible that sperm are capable of transiting against the increased fluid flow in oviducts that have been exposed to LPS, however to what extent they can go against the flow remains undetermined. Irrespective of the impact of LPS on sperm transport through the oviduct, results of this study show that the motility of bound sperm is significantly reduced.
It was recently demonstrated that exposure of bovine oviduct epithelial cells to LPS resulted in elevated expression of inflammatory and immunological response genes within three hours in vitro28. The authors identified an immediate response of oviduct epithelial cells to LPS through upregulated expression of TLR4 and associated genes (MYD88 and CD14) downstream in its pathway. This demonstrates that canonical LPS-TLR4 mediated signalling is occurring as expected34,35,36. Expression patterns of a selected panel of inflammatory and immunological response genes were assessed in the current study to determine if a similar response occurred in our ex vivo model. Although expression of CD14 increased, no increase in TLR4 expression was observed following challenge of the oviduct with LPS, suggesting that in this model LPS is not stimulating the canonical TLR4 signalling pathway and that, instead, a TLR4-independent mechanism may be in place. TLR4-independent pathways have been described by several groups and suggest that LPS stimulates non-canonical inflammasome activation in a caspase-1/caspase-11 dependent manner in mice35,37,38; whereby caspase-11 binds directly to LPS resulting in caspase oligomerization and inflammasome activation39. This non-canonical inflammasome activation has also been observed in human, where caspase-4 is involved with mediating a noncanonical response against gram-negative bacterial pathogens40.
Additionally, it was demonstrated that the expression of CASP3, a biological indicator and known executioner of apoptosis41,42,43, was upregulated following exposure of the oviduct to LPS in the current study. Increased CASP3 activity could be indicative of an apoptotic response by the bovine oviduct, mediated through LPS. It has, however, been shown using in vivo and in vitro models of microglia activation and brain inflammation that LPS-mediated induction of caspases-3, -8, and -7 results in a non-apoptotic response, as major cell death was not observed44. The authors of this investigation concluded that these caspases have a pivotal role in inflammation of the central nervous system. Therefore, LPS-induced activation of CASP3 in the ampulla observed in this study does not necessarily indicate an apoptotic response to LPS, and may be indicative of a similar inflammatory response described in44.
Classically, the oviduct is defined as a sterile environment, however pathogens and endotoxins (LPS) can invade the mucosal surfaces of the oviduct via the uterus and peritoneal cavity22. It has also been shown that, in cattle, LPS is detectable in ovarian follicular fluid; isolated in vivo from animals with different severities of uterine disease21. In this study, the authors showed that animals with clinical disease had concentrations of LPS up to 0.8 ug/ml and normal animals did not have measurable concentrations of LPS. This provides evidence that animals suffering uterine infection contain LPS in their follicular fluid, which may be delivered directly into the oviduct following ovulation. However, whether sperm can transit through infected uteri of animals with uterine disease remains largely unknown, therefore LPS or bacteria associated with uterine infection may elicit an effect on sperm prior to their transition into the oviduct. Females suffering C. trachomatis or C. abortus infections are often asymptomatic and subfertile, as opposed to completely infertile1,45. It is very plausible that in these females sperm encounter oviducts that contain or have been exposed to LPS. Therefore, the current study best represents a potential environment in females suffering Chlamydia infections. It has been previously suggested that bacterial endotoxins (LPS) can act directly on sperm through activation of TLRs present on their membranes, resulting in reduced sperm motility, apoptosis and, possibly, impairment of fertilization potential46. Results of the current study are in direct agreement with the observation of Fujita et al.46, given that bound sperm motility was reduced by 50% following a short exposure to LPS. Therefore, LPS may be exerting and effect on not only the oviduct, but may be directly affecting the sperms ability to interact with the oviductal epithelium and may also be involved with reducing the motility of the sperm that are capable of interacting with the epithelium. The impact of the observed reduction in sperm motility may disrupt its ability to interact with the oocyte, once present in the ampulla. However, at the current time this is only a hypothesis and future research should focus on elucidating the significance of reduced sperm motility following exposure to LPS.
In summary, we provide the first insight into sperm behaviour, in real time, in ex vivo oviduct sections following exposure to LPS. Oviduct function was significantly altered at both the physiological and the genetic level subsequent to a relatively short exposure to the gram negative bacterial endotoxin, LPS. These results demonstrate a rapid host response of healthy oviducts to LPS, through elevated tubal flow and increased frequency of cilia movement. The motility of sperm interacting with the oviduct was significantly affected by exposure to LPS and suggests that even a short exposure of the oviduct to infection may have dire consequences on fertility. Finally, the findings of this investigation provide evidence demonstrating that the cow is an appropriate model for studying reproductive diseases and immunity.
Materials and Methods
Experimental Overview
The physiological, inflammatory and immunological response of the bovine oviduct to LPS treatment was investigated. Firstly, bound sperm motility, PTS and CBF were interrogated using live cell imaging microscopy to examine the physiological response of ampulla to LPS treatment, Secondly, to investigate the inflammatory and immunological response of the oviduct epithelial cells to LPS, genes known to respond to LPS exposure in vitro (CASP3, CD14, MYD88, TLR4 and TRAF6)28 were analysed by real time PCR. Experiments were performed as outlined in Fig. 1.
Sample preparation and LPS treatment
Female reproductive tracts from 5 separate recently slaughtered animals were collected at a local abattoir, transported to the laboratory on ice within an hour and determined to be in the diestrus stage of the oestrus cycle based on (1) presence of a large corpus luteum on one of the ovaries (2) closed cervix (3) absence of mucus in the uterine horn and cervix. Details of the tracts used in this study (Sperm binding, PTS, CBF and the first gene expression experiment) are outlined in supplementary file 1. Ten additional tracts were collected for qPCR experiments (Fig. 5B,C). Identical parameters were used to select oviducts for the second and third qPCR experiments.
Tracts were prevented from drying out with PBS while the ipsilateral oviduct was being dissected from the uterus, trimmed free from connective tissue and each ampulla was divided in four 5 mm and two 10 mm segments. Two 5 mm segments, one for control conditions and the other for challenge with LPS, were used to measure cilia beat frequency (CBF) and bound sperm motility; the other two 5 mm to measure particle transport speed (PTS) with and without exposure to LPS. The 10 mm segments, one maintained in HEPES and the other in HEPES + LPS, were used to isolate the RNA for gene expression profiling.
The four 5 mm segments were washed in PBS, opened longitudinally and pinned down, with 1 cm pins recovered from 26 G Sterican needles (Braun, GmbH, Germany), on Delta T dishes (Bioptechs, PA, USA) coated with a 1 mm Sylgard 184 (Dow Corning, MI, USA) layer. The two 10 mm segments were transferred in a four-well dish filled with 1 ml PBS. Pinned sections were resuspended in 1.5 ml PBS a couple of times until cell debris was fully removed and discarded, then they were submerged in 1.5 ml room temperature HEPES buffer solution (5.6 mM KCl, 136.4 mM NaCl, 1 mM MgCl2-6H2O, 2.2 mM CaCl2-2H2O, 11 mM glucose, 10 mM HEPES) with and without 10 μg/ml LPS (Cat. No. L6529, Sigma-Aldrich, Germany). Ampulla segments in the four-well dish were gently washed by flushing PBS through the lumen and were finally submerged in 1 ml lukewarm HEPES buffer solution with and without 10 μg/ml LPS. The concentration was selected based on previous studies. Ibrahim et al.28 used a minimal dose of 0.5 μg/ml LPS. Given that these conditions were used in an in vitro cell culture model (in which monolayers of cells are homogenously exposed to LPS) and that 0.5 μg/ml was described as a minimal dose, a concentration of 10 μg/ml was used in the current ex vivo model. This concentration has also been used in other in vivo and in vitro models of infection21,47,48.
PTS analysis
Two ampulla sections from the ipsilateral oviduct of each animal (n = 5) were incubated in Petri dishes as per CBF for 2 h in HEPES buffer solution at 37 °C. After incubation, the dishes were transferred to the Delta T stage adapter and maintained at 37 °C. 3 μl (equivalent to 90 μg) of 2.8 μm Dynabeads Protein G (Life Technologies, AS, Norway) were added to the dish and gently resuspended a few times. Videos were recorded, in duplicate, on three different regions at 12 frames per second (FPS) for 10 sec, 640 × 480 pixel resolution. Videos were converted to 8-bit grayscale and particles were tracked with an automatic tracking procedure in-built in ImagePro (MediaCybernetics, PA, USA). Only the particles that travelled for at least 10 frames were used for calculation.
CBF analysis
Two sections of ampulla from the oviducts (ipsilateral side) of each animal (n = 5) were equilibrated in HEPES buffer solution at 37 °C in incubator for 10 min. To reduce evaporation, Delta T dishes were transferred in two humidity chambers consisting in two 10 cm closed Petri dishes whose bottom was coated with two layers of moist filter paper. Following equilibration, dishes were transferred to a Delta T stage adapter (Bioptechs, PA, USA) where temperature was controlled and maintained at 37 °C. Ciliary beating was assessed with a pre-warmed 40 × W/0.70 water immersion objective (Olympus, Hamburg, Germany) maintained at 37 °C by an objective heater (Bioptechs, PA, USA). Images were captured with the StreamPix 7.0 (NorPix, Canada) software connected to a SUMIX Mx7 camera (100 FPS/10 sec, 640 × 480 pixel resolution) mounted on a BX51WI fixed-stage upright microscope (Olympus, Hamburg, Germany). Videos were recorded on five different areas of interest (AOI) per sample (Fig. 2Ai) the central area and the four corners of each section. After imaging, dishes were emptied and replenished with a suspension of 5 × 106 spermatozoa in 1.5 ml HEPES buffer solution for the control and with 1.5 ml HEPES buffer solution supplemented with LPS for the treatment. The dishes were then incubated at 37 °C for 2 h during which videos were recorded every 30 min as previously described and buffer solutions were swapped with fresh ones before transferring the dishes back in the incubator.
Cilia beat was measured on 5 ciliated cells per AOI. The videos were converted to black and white by applying a binary mask using ImageJ (National Institutes of Health). Ciliated cells were selected within regions of approximately 40 × 40 μm, and 400 frames were analysed. Average values and frames were exported to AutoSignal (Systat Software GmbH) and frequency was calculated (Hz) using Fast Fourier Transformation (FFT).
RNA extraction and cDNA synthesis
Oviduct sections were subjected to three separate methods of exposure to LPS prior to harvesting for RNA extraction; all incubations were carried out at 37.5 °C. Firstly, oviduct epithelial cells were isolated from intact longitudinal 1 cm sections of ampulla, which had been challenged with 10 μg/ml LPS for 2 h, using non-serrated forceps in a stripping motion. Secondly, 1 cm sections of ampulla were dissected longitudinally prior to being challenged with 10 μg/ml LPS for 2 h. Cells were recovered with non-serrated forceps as above. Finally, longitudinally dissected sections of ampulla were challenged with 100 μg/ml LPS for 3 h. Epithelial cells were harvested by gently scrapping the sections with a glass slide. The buffers were changed after 1 h (1.5 h for the final method) and cells were immediately snap frozen and stored at −80 °C. All experiments had five independent biological replicates of oviduct sections exposed to LPS and control sections that were not exposed to LPS. Total RNA was extracted using an RNeasy micro kit (Qiagen) as outlined previously49. cDNA was synthesised from 200 ng of total RNA in 20 μl reactions, using the high-capacity cDNA reverse transcription kit (Applied Biosystems Inc), as follows; 1 × RT buffer, 5 μm random primers, 4 mM dNTPs, 40 U RNase inhibitor and 50 U Multiscribe RT. Reverse transcription reactions were incubated at 25 °C for 10 min, 37 °C for 120 min and 85 °C for 5 min.
Real Time PCR
Information on the genes analysed in this study has been published previously50 and the sequences of the primers that were used are outlined in Table 1. The expression of five candidate genes (CASP3, CD14, MYD88, TLR4 and TRAF6) was examined in all control and LPS-exposed oviduct samples. The geNorm function in qBasePLUS (Biogazelle, Zwijnaarde, Belgium)51 was used to analyse the stability of the candidate reference genes across all samples. The most stable reference genes, RPL19 and PPIA, were used for normalization. Real Time PCR was performed using the 7500 Fast Real-Time PCR system (Applied Biosystems®, USA).
Preparation of sperm
Sperm straws (Cryopreserved in BioxCell extender) from the same bull sire were thawed at 39 °C for 10 sec, resuspended 1 ml HEPES buffer solution and washed at 200 rcf/5 min. Supernatant was discarded and pellet was resuspended in 100 μl non-capacitating HEPES buffer solution (5.6 mM KCl, 136.4 mM NaCl, 1 mM MgCl2-6H2O, 2.2 mM CaCl2-2H2O, 11 mM Glucose, 10 mM HEPES). Sperm motility was visually assessed before and after wash using a standard phase contrast microscope. Only sperm with motility ≥70% were used for experiments. Spermatozoa were then quantified using a Neubauer chamber prior to addition to oviduct sections. Uncapacitated sperm were used in the experiments, as capacitation has been shown to reduce sperm binding to oviduct epithelial cells52,53.
Analysis of Bound Sperm Motility
Following an initial incubation of the ampulla for 2 h, in either HEPES buffer or LPS, 5 × 106 sperm were added to each section of ampulla. Five AOIs per sample were captured (n = 5 biological replicates) 90 min following addition of the sperm to the oviduct. Preliminary examination of the ampulla was performed to confirm that spermatozoa were interacting with the surface of the ampulla. The total number of motile and immotile sperm in each AOI was documented using Image Pro (MediaCybernetics). Motility was calculated by determining the ratio of motile to immotile bound sperm and is represented as a percentage.
Statistics Analysis
Bound sperm motility and PTS analysis were performed using a two sample t-test and CBF analysis was performed using an ANOVA test with ad-hoc tukeys in Mintab (Minitab Inc.). Statistical analysis of gene expression was performed using the calibrated normalised relative quantities, based on normalisation factors generated for the two reference genes (PPIA and RPL19), for each target gene using the Mann-Whitney function in the qbase + package (Biogazelle).
Additional Information
How to cite this article: O’Doherty, A. M. et al. Lipopolysaccharide (LPS) disrupts particle transport, cilia function and sperm motility in an ex vivo oviduct model. Sci. Rep. 6, 24583; doi: 10.1038/srep24583 (2016).
References
Newman, L. et al. Global Estimates of the Prevalence and Incidence of Four Curable Sexually Transmitted Infections in 2012 Based on Systematic Review and Global Reporting. PLos One 10, e0143304, 10.1371/journal.pone.0143304 (2015).
Longbottom, D. & Coulter, L. J. Animal chlamydioses and zoonotic implications. J Comp Pathol 128, 217–244 (2003).
Doull, L. et al. Late production of CXCL8 in ruminant oro-nasal turbinate cells in response to Chlamydia abortus infection. Vet Immunol Immunopathol 168, 97–102, 10.1016/j.vetimm.2015.08.011 (2015).
Foldi, J. et al. Bacterial complications of postpartum uterine involution in cattle. Anim Reprod Sci 96, 265–281, 10.1016/j.anireprosci.2006.08.006 (2006).
Swangchan-Uthai, T., Lavender, C. R., Cheng, Z., Fouladi-Nashta, A. A. & Wathes, D. C. Time course of defense mechanisms in bovine endometrium in response to lipopolysaccharide. Biol Reprod 87, 135, 10.1095/biolreprod.112.102376 (2012).
Sheldon, I. M., Williams, E. J., Miller, A. N., Nash, D. M. & Herath, S. Uterine diseases in cattle after parturition. Veterinary journal 176, 115–121, 10.1016/j.tvjl.2007.12.031 (2008).
Sheldon, I. M., Noakes, D. E., Rycroft, A. N., Pfeiffer, D. U. & Dobson, H. Influence of uterine bacterial contamination after parturition on ovarian dominant follicle selection and follicle growth and function in cattle. Reproduction 123, 837–845 (2002).
Cronin, J. G., Turner, M. L., Goetze, L., Bryant, C. E. & Sheldon, I. M. Toll-like receptor 4 and MYD88-dependent signaling mechanisms of the innate immune system are essential for the response to lipopolysaccharide by epithelial and stromal cells of the bovine endometrium. Biol Reprod 86, 51, 10.1095/biolreprod.111.092718 (2012).
Sheldon, I. M., Cronin, J., Goetze, L., Donofrio, G. & Schuberth, H. J. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol Reprod 81, 1025–1032, 10.1095/biolreprod.109.077370 (2009).
Carp, H. J., Selmi, C. & Shoenfeld, Y. The autoimmune bases of infertility and pregnancy loss. J Autoimmun 38, J266–274, 10.1016/j.jaut.2011.11.016 (2012).
Sen, A., Kushnir, V. A., Barad, D. H. & Gleicher, N. Endocrine autoimmune diseases and female infertility. Nat Rev Endocrinol 10, 37–50, 10.1038/nrendo.2013.212 (2014).
Lonergan, P. & Fair, T. In vitro-produced bovine embryos: dealing with the warts. Theriogenology 69, 17–22, 10.1016/j.theriogenology.2007.09.007 (2008).
Suarez, S. S. Mammalian sperm interactions with the female reproductive tract. Cell Tissue Res. 10.1007/s00441-015-2244-2 (2015).
Pollard, J. W. et al. Fertilizing capacity of bovine sperm may be maintained by binding of oviductal epithelial cells. Biol Reprod 44, 102–107 (1991).
Li, E., Beard, C. & Jaenisch, R. Role for DNA methylation in genomic imprinting. Nature 366, 362–365, 10.1038/366362a0 (1993).
Market-Velker, B. A., Zhang, L., Magri, L. S., Bonvissuto, A. C. & Mann, M. R. Dual effects of superovulation: loss of maternal and paternal imprinted methylation in a dose-dependent manner. Human molecular genetics 19, 36–51, 10.1093/hmg/ddp465 (2010).
Suarez, S. S. The oviductal sperm reservoir in mammals: mechanisms of formation. Biol Reprod 58, 1105–1107 (1998).
Katz-Jaffe, M. G., McCallie, B. R., Preis, K. A., Filipovits, J. & Gardner, D. K. Transcriptome analysis of in vivo and in vitro matured bovine MII oocytes. Theriogenology 71, 939–946, 10.1016/j.theriogenology.2008.10.024 (2009).
Domes, T. et al. The incidence and effect of bacteriospermia and elevated seminal leukocytes on semen parameters. Fertil Steril 97, 1050–1055, 10.1016/j.fertnstert.2012.01.124 (2012).
Tung, C. K. et al. Microgrooves and fluid flows provide preferential passageways for sperm over pathogen Tritrichomonas foetus. Proc Natl Acad Sci USA 112, 5431–5436, 10.1073/pnas.1500541112 (2015).
Herath, S. et al. Ovarian follicular cells have innate immune capabilities that modulate their endocrine function. Reproduction 134, 683–693, 10.1530/REP-07-0229 (2007).
Kowsar, R. et al. Regulation of innate immune function in bovine oviduct epithelial cells in culture: the homeostatic role of epithelial cells in balancing Th1/Th2 response. J Reprod Dev 59, 470–478 (2013).
Neyen, C. & Lemaitre, B. Sensing Gram-negative bacteria: a phylogenetic perspective. Curr Opin Immunol 38, 8–17, 10.1016/j.coi.2015.10.007 (2015).
Hori, N. et al. Aberrant CpG methylation of the imprinting control region KvDMR1 detected in assisted reproductive technology-produced calves and pathogenesis of large offspring syndrome. Anim Reprod Sci 122, 303–312, 10.1016/j.anireprosci.2010.09.008 (2010).
Ling, G. S. et al. Integrin CD11b positively regulates TLR4-induced signalling pathways in dendritic cells but not in macrophages. Nat Commun 5, 3039, 10.1038/ncomms4039 (2014).
Sabroe, I., Jones, E. C., Usher, L. R., Whyte, M. K. & Dower, S. K. Toll-like receptor (TLR)2 and TLR4 in human peripheral blood granulocytes: a critical role for monocytes in leukocyte lipopolysaccharide responses. J Immunol 168, 4701–4710 (2002).
Yang, X. et al. Gene body methylation can alter gene expression and is a therapeutic target in cancer. Cancer cell 26, 577–590, 10.1016/j.ccr.2014.07.028 (2014).
Ibrahim, S. et al. Expression pattern of inflammatory response genes and their regulatory micrornas in bovine oviductal cells in response to lipopolysaccharide: implication for early embryonic development. PLos One 10, e0119388, 10.1371/journal.pone.0119388 (2015).
Kolle, S. et al. Ciliary transport, gamete interaction, and effects of the early embryo in the oviduct: ex vivo analyses using a new digital videomicroscopic system in the cow. Biol Reprod 81, 267–274, 10.1095/biolreprod.108.073874 (2009).
Shi, D., Komatsu, K., Uemura, T. & Fujimori, T. Analysis of ciliary beat frequency and ovum transport ability in the mouse oviduct. Genes to cells: devoted to molecular & cellular mechanisms 16, 282–290, 10.1111/j.1365-2443.2011.01484.x (2011).
Shi, D. et al. Celsr1 is required for the generation of polarity at multiple levels of the mouse oviduct. Development 141, 4558–4568, 10.1242/dev.115659 (2014).
Bermbach, S. et al. Mechanisms of cilia-driven transport in the airways in the absence of mucus. Am J Respir Cell Mol Biol 51, 56–67, 10.1165/rcmb.2012-0530OC (2014).
Miki, K. & Clapham, D. E. Rheotaxis guides mammalian sperm. Current biology: CB 23, 443–452, 10.1016/j.cub.2013.02.007 (2013).
Kawai, T. & Akira, S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol 11, 373–384, 10.1038/ni.1863 (2010).
Molloy, S. Host response: LPS goes non-canonical. Nat Rev Microbiol 11, 599, 10.1038/nrmicro3106 (2013).
Akira, S., Uematsu, S. & Takeuchi, O. Pathogen recognition and innate immunity. Cell 124, 783–801, 10.1016/j.cell.2006.02.015 (2006).
Kayagaki, N. et al. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science 341, 1246–1249, 10.1126/science.1240248 (2013).
Hagar, J. A., Powell, D. A., Aachoui, Y., Ernst, R. K. & Miao, E. A. Cytoplasmic LPS activates caspase-11: implications in TLR4-independent endotoxic shock. Science 341, 1250–1253, 10.1126/science.1240988 (2013).
Shi, J. et al. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 514, 187–192, 10.1038/nature13683 (2014).
Brad, A. M. et al. Glutathione and adenosine triphosphate content of in vivo and in vitro matured porcine oocytes. Molecular reproduction and development 64, 492–498, 10.1002/mrd.10254 (2003).
Nicholson, D. W. et al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 376, 37–43, 10.1038/376037a0 (1995).
Cohen, G. M. Caspases: the executioners of apoptosis. Biochem J 326 (Pt 1), 1–16 (1997).
Green, D. & Kroemer, G. The central executioners of apoptosis: caspases or mitochondria? Trends Cell Biol 8, 267–271 (1998).
Burguillos, M. A. et al. Caspase signalling controls microglia activation and neurotoxicity. Nature 472, 319–324, 10.1038/nature09788 (2011).
DeGraves, F. J. et al. Reinfection with Chlamydophila abortus by uterine and indirect cohort routes reduces fertility in cattle preexposed to Chlamydophila. Infect Immun 72, 2538–2545 (2004).
Fujita, Y. et al. Toll-like receptors (TLR) 2 and 4 on human sperm recognize bacterial endotoxins and mediate apoptosis. Hum Reprod 26, 2799–2806, 10.1093/humrep/der234 (2011).
Richer, E. et al. N-ethyl-N-nitrosourea-induced mutation in ubiquitin-specific peptidase 18 causes hyperactivation of IFN-alphass signaling and suppresses STAT4-induced IFN-gamma production, resulting in increased susceptibility to Salmonella typhimurium. J Immunol 185, 3593–3601, 10.4049/jimmunol.1000890 (2010).
Williams, E. J. et al. The effect of Escherichia coli lipopolysaccharide and tumour necrosis factor alpha on ovarian function. American journal of reproductive immunology 60, 462–473 (2008).
O’Doherty, A. M., O’Shea, L. C. & Fair, T. Bovine DNA methylation imprints are established in an oocyte size-specific manner, which are coordinated with the expression of the DNMT3 family proteins. Biol Reprod 86, 67, biolreprod.111.094946 [pii] 10.1095/biolreprod.111. 094946 (2012).
Taraktsogloua, M. et al. Transcriptional profiling of immune genes in bovine monocyte-derived macrophages exposed to bacterial antigens. Vet Immunol Immunopathol 140, 130 –9, 10.1016/j.vetimm.2010.12.002 (2007).
Hellemans, J., Mortier, G., De Paepe, A., Speleman, F. & Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome biology 8, R19, 10.1186/gb-2007-8-2-r19 (2007).
Suarez, S. S. Regulation of sperm storage and movement in the mammalian oviduct. Int J Dev Biol 52, 455–462, 10.1387/ijdb.072527ss (2008).
Teijeiro, J. M., Dapino, D. G. & Marini, P. E. Porcine oviduct sperm binding glycoprotein and its deleterious effect on sperm: a mechanism for negative selection of sperm? Biological research 44, 329–337, /S0716-97602011000400003 (2011).
Acknowledgements
The authors would like to thank the staff, especially Paul Delaney, at a local abattoir for assistance with recovery of the female reproductive tracts used in this investigation. We would also like to thank Prof. David McHugh, Dr. David Magee and Dr. John Brown (University College Dublin) for providing primer and access to qBasePLUS software (Biogazelle) and Dr. Erin Williams (University of Edinburgh) for advice on manuscript preparation.
Author information
Authors and Affiliations
Contributions
A.M.O. conceived the study. A.M.O. and M.D.F. designed the experiments, carried out the experiments, performed analysis and prepared the manuscript. S.K. provided Live Cell Imaging equipment, funding and comments for manuscript preparation.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
O’Doherty, A., Di Fenza, M. & Kölle, S. Lipopolysaccharide (LPS) disrupts particle transport, cilia function and sperm motility in an ex vivo oviduct model. Sci Rep 6, 24583 (2016). https://doi.org/10.1038/srep24583
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep24583
This article is cited by
-
The complex microbiome from native semen to embryo culture environment in human in vitro fertilization procedure
Reproductive Biology and Endocrinology (2020)
-
Bovine sperm-oviduct interactions are characterized by specific sperm behaviour, ultrastructure and tubal reactions which are impacted by sex sorting
Scientific Reports (2020)
-
Growth kinetics of Chlamydia trachomatis in primary human Sertoli cells
Scientific Reports (2019)
-
Salpingitis Impairs Bovine Tubal Function and Sperm-Oviduct Interaction
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
