
Abstract
Serpentinization of peridotites in Earth’s mantle is associated with the generation of hydrogen and low molecular weight organics that could support subsurface life. Studies of microbial metabolisms in peridotite-hosted environments have focused primarily on methanogenesis, yet DNA sequences, isotopic composition of sulfides and thermodynamic calculations suggest there is potential for microbial sulfate reduction too. Here, we use a sulfate radiotracer-based method to quantify microbial sulfate reduction rates in serpentinization fluids recovered from boreholes in the Samail Ophiolite, Oman and the California Coast Range Ophiolite, USA. We find that low levels of sulfate reduction occur at pH up to 12.3. These low levels could not be stimulated by addition of hydrogen, methane or small organic acids, which indicates that this metabolism is limited by factors other than substrate availability. Cellular activity drops at pH > 10.5 which suggests that high fluid pH exerts a strong control on sulfate-reducing organisms in peridotites.
Similar content being viewed by others


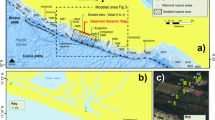
Introduction
The last two decades of planetary exploration have identified a number of environments beyond Earth that may contain liquid water, including the Martian subsurface1 and the sub-ice oceans of outer solar system satellites2,3. The presence of liquid water is necessary but not sufficient to create habitable conditions, and attention has now begun to focus on the potential for reaction of water with rocks to supply the energy and raw materials that are also required by life4,5.
Particular attention has focused on serpentinization—the aqueous alteration of ultramafic (Mg/Fe-rich) rocks (peridotite) to yield serpentine and other mineral end products, along with H2, methane, and low molecular weight organic acids6,7,8. H2 and low molecular weight organics are widely utilized as substrates by microorganisms on Earth. By generating these substrates in ongoing fashion, serpentinization could provide a sustained feedstock for life independently of photosynthesis, with important implications for the potential for life on the early Earth and elsewhere in our solar system. Moreover, the alkaline and H2-rich conditions created during serpentinization are proposed to have supported life’s origin by providing sustained sources of energy and reducing power for abiotic carbon fixation7,9,10.
As a major component of the chondritic materials from which the solar system formed, ultramafic rocks that undergo serpentinization are thought to be prevalent in the rocky bodies of our solar system. Both olivine and serpentine—reactant and product, respectively, in the serpentinization process—have been identified on the surface of Mars11,12. And, while the mineralogy of the interiors of the icy satellites remains unknown, observations made by the Cassini spacecraft at Enceladus reveal an alkaline and H2- and CH4-rich ocean that is consistent with a chemistry influenced by serpentinization13,14.
Microbial methanogenesis has long been seen as a model metabolism for photosynthesis-independent life in serpentinizing systems and a number of studies have sought to document its occurrence in such environments on Earth15,16,17. However, the H2 and organic acids that are associated with serpentinization can also be used by sulfate-reducing bacteria (SRB) when sulfate, an energetically more favorable oxidant than carbon dioxide, is present18,19. Dissimilatory sulfate reduction is among the oldest known microbial processes on Earth. Its occurrence is geochemically traceable to at least 2.7 Ga, shortly after the first appearance of sulfate on Earth, and concentrations of sulfate capable of enabling microbial sulfate reduction were present in the oceans by the early Achaean20,21,22. On Earth, sulfate reduction (SR) is the predominant anaerobic microbial terminal oxidation process in sulfur-rich marine sediments, where it outcompetes methanogenesis for the products of organic matter fermentation23, and it is also documented to occur in lakes and lake sediments24,25,26, the terrestrial rocky subsurface27, and subsurface lithoautotrophic ecosystems, where it is driven by radiolytically produced H2 and sulfate28. The presence of microbial sulfate reduction in low temperature serpentinizing systems has previously been inferred from the presence of sulfide minerals with a light isotopic signature (i.e., in serpentinites of the Iberian Margin with δ34S of −20 to −45‰) that is consistent with a microbial origin29,30. Findings of genetic material related to SRB also suggested the presence of sulfate reduction in this environment16,31,32. Active sulfate reduction was furthermore demonstrated in vents fluids and microbial mats collected from carbonate chimneys at the Lost City vent field (Mid-Atlantic Ridge) where sulfate reducers may benefit from energetic substrates generated by serpentinization in the underlying mantle33. However, direct quantification of microbial sulfate reduction rates (SRR) and their variation across the range of conditions that occur inside serpentinizing systems has not been reported.
Sulfate appears to be prevalent in environments beyond Earth where serpentinization is envisioned to occur. Orbital and lander observations made at Mars show geographically widespread sulfates in sedimentary deposits dating to the late Noachian and Hesperian epochs34, following which there is little evidence of water-related minerals. Thus, sulfate-rich fluids would have been an increasingly prominent habitat during the late stages of any surface biosphere on Mars and likely remain so in subsurface aquifers35. Near-infrared spectra of the surface of Jupiter’s moon Europa indicate the presence of sulfates, potentially as magnesium sulfate and sulfuric acid36,37. The prevalence of these features on the trailing hemisphere suggests that these deposits may result from deposition and radiation processing of sulfur efflux from the neighboring moon Io38, and ice overturn could deliver substantial quantities of this material into the ocean39. Some models of fluid chemical evolution for Europa also predict that sulfate could be an important endogenous component of ocean chemistry40,41. Given the seeming prevalence of sulfate in these environments, constraining the potential for microbial SR particularly under conditions that intersect with active serpentinization is important for understanding their habitability.
On Earth, active serpentinization and associated microbial activity is most easily studied in ophiolites—tectonic emplacements of ocean crust onto land that expose ultramafic rocks. We sought to determine whether biological SR occurs, and over what range of chemical conditions, in two such settings—the Samail Ophiolite, Oman, and the California Coast Range Ophiolite, USA. Both systems contain moderate levels of sulfate and sequences of SRB-associated genetic material but, to our knowledge, active SR has not been demonstrated within these serpentinizing systems.
Results and discussion
Active microbial sulfate reduction in peridotite fluids
We employed a sulfate radiotracer-based method to quantify microbial SRR in fluids from 17 different wells located in ophiolites on two different continents (Samail Ophiolite: 6 wells, Coast Range Ophiolite: 11 wells). The wells were previously drilled into differing host rocks (peridotite and peridotite/gabbro contact) and their current fluid chemistries span a range in pH (7.4–12.3) and concentrations of sulfate (40–1100 µmol L−1), H2 (below detection –21 µmol L−1), and CH4 (0.01–1751 µmol L−1) (Supplementary Data 1). The well fluids also contain low molecular weight organic acids (formate, acetate, propionate, and butyrate) in the low micromolar range31,42. We classified the fluids according to pH and well lithology as hyperalkaline peridotite fluids (pH > 10.5), alkaline peridotite fluids (pH 8–10.5), meteoric/shallow water fluids (Coast Range Ophiolite only), and peridotite/gabbro contact fluids (Samail Ophiolite only) (Supplementary Data 1). Broadly, fluid chemistry increasingly influenced by ongoing serpentinization, as indicated by higher pH and H2 concentrations, was accompanied by lower sulfate concentrations (Fig. 1).

pH, H2, and sulfate concentrations of the well samples from Samail Ophiolite (a–c) and Coast Range Ophiolite (d–f). Wells are grouped according to their classification by pH and fluid origin (light red background: gabbro/peridotite contact fluids; light blue background: alkaline peridotite fluids, dark blue background: hyperalkaline peridotite fluids, green background: meteoric/surface water).
To quantify SRR, we amended anoxically preserved fluid samples in the laboratory with trace amounts of 35S-sodium sulfate. Following anoxic incubation of the fluids for 10 days at near in situ temperatures (35° and 16 °C, respectively, for Samail Ophiolite and Coast Range Ophiolite, Supplementary Data 1), we measured radioactivity in the reduced sulfur fraction (all sulfur species less oxidized than sulfate) chemically separated from the incubated samples. SRR were then calculated based on the fraction of the radiotracer turned over during the runtime of the incubation experiment (see Method section for details). The use of radio-isotopically labeled trace compounds is an extremely sensitive method by which to quantify the rate of substrate metabolism. We employed a method based on established protocols43,44 that was optimized to increase sensitivity by increasing the radioactivity of the substrate tracer45. Reduction of 35S-sulfate occurred in all fluids at rates ranging between <10−3 and 2.1 pmol mL−1 d−1 (Fig. 2 and Supplementary Data 2 and 3) with only few samples below detection (mean detection limit (MDL) = 5 × 10−4 pmol mL−1 d−1, see Methods section). The results clearly show that SR is active in this environmental setting across all wells and fluid chemistries. Our findings represent the first direct measurement of such activity within serpentinizing rocks, significantly augmenting indirect evidence that sulfate reducers are present in serpentinizing systems31,32,42. To our knowledge, this is also the first report of active biological SR in an environmental system at pH above 10.65 (Lake Tanatar I, Siberia46), here observed in 5 of the 17 wells surveyed. Measured SRR are 3–4 orders of magnitude lower than those commonly observed in sulfate-containing shallow marine sediments47,48 and, at most, comparable with lowest rates found in the deepest samples thus far recovered from subseafloor sediments45. The very low SRR suggest that factors other than the availability of sulfate may be limiting to activity, given that SR is known to operate at sulfate concentrations well below those measured in any of the fluids included in this study (to as low as 10 µmol L−1 47,49,50).

Measured sulfate reduction rates (SRR) in incubations of fluids from Samail Ophiolite (a, b) and Coast Range Ophiolite (c, d). a, c Colors represent amendment of electron donors to the incubations (light blue: no substrate, orange: H2 amendment, yellow: acetate amendment, gray: CH4 amendment, dark blue: formate amendment); error bars represent standard deviation (±1σ) of three replicates. b, d Average SRR calculated from all treatments; error bars represent standard deviation (±1σ) of all replicates and amendments. Wells are grouped according to their classification by pH and fluid origin (light red background: gabbro/peridotite contact fluids; light blue background: alkaline peridotite fluids, dark blue background: hyperalkaline peridotite fluids, green background: meteoric/surface water).
To evaluate whether the availability of electron donors is limiting to SRR, we incubated a subset of samples with amendments of H2, CH4, and organic acids (the latter, as formate and acetate, were only used with Samail Ophiolite fluids). Based on a maximum RSD observed from triplicate experiments (91%, Supplementary Data 2 and 3) and an increase of rates by >10σ in order to be considered a significant stimulation, SRR must increase by a factor of at least 10 to be considered as significantly stimulated (see Supplementary Methods 1.1. for more details). In Samail Ophiolite fluids, SRR were not significantly stimulated by the addition of any of these substrates (Fig. 2a, c), suggesting that the availability of electron donors is not limiting to rates in this system. Alternatively, the incubation time of 10 days might simply have been too short to allow the community to respond to the sudden changes in substrate availability. A delay in response may occur even more in microbial communities that are adapted to the strong oligotrophic conditions in the ophiolite and thus operate at extremely slow metabolic rates. This hypothesis could be tested in future long-term experiments that include an extended pre-incubation phase where microbes can adapt to the modified environmental conditions. In Coast Range Ophiolite fluids, a small stimulation of SRR (<11-fold) occurred with addition of H2 to well fluids that had ambient H2 concentrations near or below the detection limit (<0.15 nmol L−1), which comprise all of the wells influenced primarily by meteoric water and some of the alkaline peridotite wells. The lowest SRR were found in hyperalkaline fluids with pH above 10.5 (Fig. 2), suggesting that pH is a critical limiting factor.
High pH and low sulfate concentrations as potential limitations for sulfate reduction
Our observations extend the range of observed microbial sulfate-reducing activity to higher pH. Alkaliphilic SRB isolated from alkaline soda lakes grow at pH up to 10.7 with an optimum between pH 9 and 1051,52. Microbial SRR in the range of several nmol mL−1 d−1, which are 103- to 106-fold higher than in the Samail Ophiolite and Coast Range Ophiolite fluids, have been measured in Mono and Searles Lakes (USA, pH 9.8)53,54, Big Soda Lake (USA, pH 9.7)55, a number of soda lakes in the Kulunda Steppe (Russia, pH 9.6–10.65)46, and in sediment pore waters of Lake Van (Turkey, pH 9.8)25. The much higher sulfate concentrations characteristic of these environments (tens to several hundreds of mmol L−1) relative to the Coast Range Ophiolite and Samail Ophiolite fluids could account for the difference in rates if sulfate concentration is a limiting factor. However, a 20-fold difference in rates remains even when compared to deeper sediments at Lake Van, where sulfate is depleted to about 1 mmol L−1 (equivalent to the sulfate concentration in WAB188 (Supplementary Data 1)), the highest observed in this study, although it must be noted that rates determined on sediments may include sediment particle-attached organisms, which is not the case in rates determined in fluids. Theoretically, the higher SRR observed in the soda lakes could also result from the higher concentrations and potential availability of organic carbon compared to the serpentinizing system. However, the lack of stimulation of SRR with addition of several potential electron donors would seem to rule out this possibility.
To our knowledge, only one other study has sought to find evidence for active microbial life in fluids with pH > 10.7 and high (20–30 mmol L−1) sulfate concentrations8. That study, conducted on fluids recovered from serpentinizing mud volcanoes at the Mariana Trench56, did not exhibit significant microbial abundance or activity. In that system, the high temperatures in the basement rocks sourcing the fluids (80–250 °C) combined with the very high pH (11–12.5) may have prevented microbial recolonization on the timescale associated with fluid ascent through the system. In the Samail Ophiolite and Coast Range Ophiolite systems, the evolution of fluid pH from lower to higher values as the extent of fluid–rock interaction increases allows the possibility that sulfate reducer populations are established under more moderate and permissive pH levels. Activity observed at the highest pH range could then result from cells that persist from lower pH fluids in a low activity state, rather than populations that have actively grown under such conditions. Activity of this type would not be observed in culture experiments, which typically focus on growth, but documenting its occurrence is important in mapping the boundaries of habitability in serpentinizing systems, and a first step toward understanding the adaptations that make life possible in these environments.
With the available sample set, it is challenging to disentangle the effects of high pH versus low sulfate concentrations on the SRR; however, some trends are evident (Fig. 3). SRR are consistently lowest in fluids with pH > 10.7, equivalent to the upper limit for growth of sulfate-reducing isolates from soda lakes. All but one of these fluids also fall into the lowest range of observed sulfate concentrations (<0.1 mmol L−1), so that either high pH or low sulfate, or the combination, could be limiting to rates. However, the set of well fluids that have sulfate concentrations <0.1 mmol L−1 includes seven wells with pH < 10.7 and rates are higher and more variable within this pool, suggesting that low sulfate concentrations alone do not impose the primary limitation on activity. This is consistent with the observation that microbial sulfate reduction in marine sediments (which have circumneutral pH) remains active with sulfate concentrations as low as 0.01 mmol L−1. Whether low rates at pH > 10.7 result from a combination of low sulfate and high pH, or from high pH alone, remains uncertain because we lack fluid samples having a combination of high pH and high sulfate.

Measured sulfate reduction rates (SRR) (average of all replicates and amendments) in fluids from Samail Ophiolite and Coast Range Ophiolite in the pH-sulfate concentration space. SRR is indicated by the coloring of the dots ranging from dark blue (0 pmol mL−1 d−1) to yellow (0.2 pmol mL−1 d−1).
Consistent with the rate data, cell numbers and community richness are significantly lower above pH 10.5 at both Coast Range Ophiolite31 and Samail Ophiolite42, as shown in Fig. 4a, c. This supports the notion that the very high pH in these fluids may represent the primary limitation for microbial life, limiting the community to a few species capable of survival in this challenging environment. Interestingly, previous work at Coast Range Ophiolite showed differences between the transcription of sulfate reduction-related genes and their abundance in metagenomes showing that despite a potentially low abundance of SRB, they may be highly active32.

Cell abundance (gray bars) and relative abundance (yellow dots) of sulfate-reducing bacteria (SRB) (a, c) and cell-specific sulfate reduction rates (SRR, mean of all treatments) (b, d) in Samail Ophiolite and Coast Range Ophiolite fluids. Wells are grouped according to their classification by pH and fluid origin (light red background: gabbro/peridotite contact fluids; light blue background: alkaline peridotite fluids, dark blue background: hyperalkaline peridotite fluids, green background: meteoric/surface water).
Cell-specific sulfate turnover rates
We normalized SRR to total cell abundance in order to determine the extent to which low rates at high pH result from lower cell numbers, and to enable comparison with cell-specific SRR determined in other environments. When normalized to total cell numbers, SRR are 2 × 10−3 to 1 fmol cell−1 year−1 (Fig. 4b, e). These rates fall in the range of SRR reported for deep marine sediments (1.5 × 10−5 to 3.3 fmol cell−1 year−1) and are several orders of magnitude lower than cell-specific SRR measured in marine surface sediments and in pure cultures of SRB in the laboratory57,58,59. The cell-specific SRR in fluids from hyperalkaline peridotite in Samail Ophiolite is about 200-fold lower than in alkaline peridotite fluids from Samail Ophiolite, whereas this difference is only about 20-fold in Coast Range Ophiolite fluids. These observations indicate that the lower bulk SRR in hyperalkaline vs. alkaline fluids is not simply a result of a decrease in total microbial abundance but may be related to a decrease in metabolic activity of the sulfate-reducing cells under very high pH conditions.
SRB associated with the families Thermodesulfovibrionaceae and Desulforudaceae were identified in Samail Ophiolite and Coast Range Ophiolite fluids and additionally Desulfovibrionaceae and Desulfitobacteriaceae in Coast Range Ophiolite fluids (Supplementary Tables 1 and 2). Using strict criteria for linking taxonomy to function (see Methods section and Supplementary Methods 1.2. and 1.3.), SRB constitute up to 38.59% of the total population for Samail Ophiolite and Coast Range Ophiolite, respectively (Fig. 4a, c and Supplementary Data 2 and 3). In the Samail Ophiolite, SRB comprise 21.3–38.5% of cells in hyperalkaline fluids but only 0.5–2.5% in alkaline fluids. In the Coast Range Ophiolite, SRB were only found in one of four the hyperalkaline fluids (CSW13, 1.87% of total population). In the other three hyperalkaline fluids of Coast Range Ophiolite, we could still detect sequences that are potentially SRB, given that metagenomic analyses have pointed to the widespread occurrence of genes associated with sulfate reduction, even in taxa with uncultured representatives60. However, we excluded them from the SRB abundance calculation due to our strict filtering criteria. On this basis, SRB abundance in alkaline Coast Range Ophiolite fluids was between 0.07 and 36.22%. For comparison, in sulfate-containing marine sediments, SRB typically comprise 1–20% of the microbial population58,61,62. The contribution of SRB in the hyperalkaline fluids from Oman is in the higher part of this range or even distinctly higher. The high relative abundance of SRB in the serpentinized fluids is remarkable given the very low SRR we measured. A possible explanation is that SRB utilize sulfur intermediates such as thiosulfate or other species produced by the reoxidation of sulfide, the potential for which is suggested by metagenomic analysis32. Those metabolic pathways are not captured in our rate assay and may additionally contribute to the high abundance of SRB in the fluids. Regardless of the specific metabolic processes they mediate, the high abundance of SRB at alkaline and hyperalkaline pH, where overall community richness diminishes, suggests adaptation to alkaline conditions and indicates that they are an important part of the serpentinite microbiome.
High pH is a challenge for microbial metabolism because cells must expend energy to maintain cytoplasmic pH homeostasis, and also because it is difficult to maintain a high proton motive force across the cell membrane to generate ATP at the exceptionally low proton concentrations in hyperalkaline fluids63,64. Some organisms overcome the latter problem and adapt to alkaline conditions by utilizing sodium ion gradients instead of proton gradients in chemiosmotic ATP production65,66. A search for genes that encode components of sodium ion-based energy metabolism in sulfate-reducing and other organisms from serpentinizing systems could serve to determine whether such an adaptation is present in the communities that inhabit serpentinizing systems. The challenge of energy expenditure to maintain pH homeostasis would remain even if such adaptations are present, and the low cell-specific SRR we measured, corresponding to low rates of metabolic energy production, would seem to compound this challenge.
Co-occurrence of sulfate reduction and methanogenesis
The presence and activity of SRB does not appear to preclude methanogenesis in the Samail Ophiolite system. SRB typically prevail in direct competition with methanogens for common substrates (e.g., H2) when sulfate is present at concentrations comparable to those in the well fluids studied here19,67,68. However, recent studies have shown that such zonation is not universal, with methanogenesis observed in the sulfate reduction zone and vice versa50,69,70. This appears to be true in the case of the Samail Ophiolite. In a recent study of methanogenesis in the same well fluids from Samail Ophiolite that we study here, Fones et al.16 demonstrated the potential for microbial reduction of CO2 to CH4 at rates of 10 to >1000 pmol mL−1 d−1. “Potential” acknowledges the possibility that rates were stimulated by addition of the substrate bicarbonate at higher than in situ levels and, for this reason, the rates are not directly comparable to those reported here for SR. However, the results of Fones et al. combined with the present study demonstrate that methanogenesis and sulfate reduction are simultaneously viable in the Samail Ophiolite fluids.
Conclusive summary
We demonstrated the active occurrence of microbial sulfate reduction across a range of conditions in fluids from the actively serpentinizing Samail Ophiolite and Coast Range Ophiolite systems. Bulk and cell-specific SRR were in the range of <10−3–2.1 pmol mL−1 d−1 and 2 × 10−3–1 fmol cell−1 year−1, respectively, and are comparable to the lower end of the range observed in deep subsurface environments. SRR do not appear to be limited by availability of suitable electron donors (H2, CH4, small organic acids) but rather by high pH either alone or in combination with low sulfate concentrations. Both bulk and cell-specific rates are lowest in the hyperalkaline wells, where pH is highest. The decrease in rates at these high pH values accompanies a drop in both overall cell abundance and community richness, although it is notable that in the Samail Ophiolite fluids SRB comprise a larger fraction of the hyperalkaline community than at lower pH.
To our knowledge, this is the first documentation of active microbial sulfate reduction in serpentinizing systems and at pH > 10.7. These observations bear directly on the potential for long-term habitability in subsurface environments—including on Mars, Europa, Enceladus, and other “ocean worlds”—where sunlight or the chemical products of photosynthesis are not available as an energy source for life. Serpentinizing systems have long been considered a model habitat in these environments, owing to both the likely prevalence of ultramafic minerals in the rocky bodies of our solar system and their reaction to produce H2 and other potential microbial substrates. The unique chemistry of these systems poses challenges to life in the form of both high pH and potentially limited inorganic carbon availability. However, our results now demonstrate that sulfate, which is observed to be abundant in surface deposits on Mars and Europa, can be utilized as an oxidant to fuel microbial metabolism in two geographically distinct serpentinizing systems whose fluid chemistry spans a range in both pH and sulfate availability.
Methods
Site description and sampling
California Coast Range Ophiolite, United States: the California Coast Range stretches north of San Francisco for about 600 km, roughly parallel to the Pacific Coast line and bordered on the east side by the Great Valley of California. The Coast Range Ophiolite comprises weathering ultramafic bedrocks (mainly olivine and pyroxene minerals in peridotite) from Jurassic oceanic lithosphere tectonically displaced on land71. Parts of the Coast Range Ophiolite are undergoing low temperature serpentinization by reaction with trapped Cretaceous seawater72,73. The Coast Range Ophiolite Microbial Observatory comprises an array of wells drilled and cased to varying depth into the subterranean aquifers in the ophiolite and provides access to deeply sourced serpentinizing fluids71. We collected samples from 11 wells (Supplementary Data 1). Fluids were collected with a bladder pump and directly passed through a flow-through cell equipped with temperature and pH sensors before sample collection by 60 mL disposable syringes. Subsamples of fluids for rate measurements and sulfate analysis were immediately transferred by syringe into combusted (5 h at 450 °C), sealed, and N2-filled 120 mL serum vials and stored at 4 °C. Fluid samples for analysis of dissolved gases (typically 50 mL) were collected in a 60-mL syringe and extracted with 10 mL N2 gas. The gas phase was transferred by syringe into sealed glass vials (Labco exetainer) that had been completely filled with a saturated sodium chloride solution. Some of this solution was displaced through a vent needle as the gas phase was added, with the remaining solution serving as a barrier to gas diffusion when the vials were stored in an inverted position. Gas concentrations were typically determined within 1 day after sampling. Fluids collected to extract DNA from microbial biomass were filtered through Sterivex filters (pore size 0.2 µm), and flash frozen in liquid nitrogen in the field.
Samail Ophiolite, Oman: the Samail Ophiolite in the Sultanate of Oman is composed of mafic and ultramafic rocks (gabbros and peridotites) exposed within a cross-section of several km of oceanic crust and underlying upper mantle. It is the largest accessible ophiolite on Earth and hosts >15,000 km3 of peridotite that is locally undergoing low temperature seprentinization (<60 °C)74 representing the largest accessible ophiolite on Earth. Ongoing serpentinization produces alkaline and hyperalkaline waters with high concentrations of dissolved hydrogen and methane75. Detailed descriptions of the wells sampled in this study can be found in Rempfert et al.42 and the geological and hydrological features of the Samail Ophiolite have been described previously76. We sampled six wells previously drilled by the water ministry of the Sultanate of Oman spanning the crust-mantle transition in the Tayin massif. Fluids were collected with a Grundfos submersible pump at the specified depth (Supplementary Data 1) and stored in combusted (5 h at 450 °C) and N2-filled serum bottles at room temperature. Cell abundance, microbial community composition, and well fluid chemistry at the Samail Ophiolite have been reported previously16,42.
Sulfate reduction rate determination
We determined SRR by incubation with radioactively labeled (35S) sulfate according to published protocols44,77. We injected 5-mL fluid samples with a syringe and needle via a septum into a sealed, combusted (450 °C, 5 h) 20 mL glass vial, which contained oxygen-free N2 atmosphere. In one set of samples, we exchanged the headspace gas with H2 and in a second set with CH4. For Samail Ophiolite fluids (only), a third and fourth set of samples were amended separately with formate and acetate to a final concentration of 100 µmol L−1. Subsequently we injected 3 MBq (approx. 15 µL) of carrier-free 35SO42− into each vial and incubated the samples at approximate in situ temperate (15 °C for Coast Range Ophiolite, 35 °C for Samail Ophiolite) for 10 days. All incubations were prepared in triplicates. The high radioactivity utilized in these incubations was designed to yield increased sensitivity in measured rates45. Following the incubation, we transferred the content of the vials into 50 mL Falcon tubes containing 10 mL of zinc acetate solution (20%, v:v) and stored the vials frozen until further analysis.
Separation of the reduced sulfur species from the incubated samples was done by a cold-chromium distillation procedure according to published protocols43,44,45. To determine the total radioactivity of samples, we withdrew a 100 µL subsample prior to the distillation and mixed it with 5 mL zinc acetate (5%, v:v) and 15 mL scintillation fluid (Ecoscint XR, National Diagnostics, USA) in a 20 mL scintillation vial. The remaining sample was transferred into a 100 mL round flask and mixed with 10 mL DMSO. Addition of 8 mL of a 6 N HCl and 15 mL of CrCl2 solution in 2 N HCl serve to drive reduction of all species more reduced than sulfate to hydrogen sulfide. Bubbling with an N2 gas stream transferred the evolving H2S gas through a citric acid wash and into 5 mL of zinc acetate solution (5%, v:v), where it is quantitatively trapped as zinc sulfide. After a 1.5 h distillation, the solution containing the precipitated zinc sulfide was mixed with 15 mL of scintillation fluid. Radioactivity in both, total sample (aTOT) and total reduced inorganic sulfur fraction (aTRIS), was measured as counts per minute (CPM) on a scintillation counter (Tri-Carb, Perkin Elmer).
SRR per sample were calculated according to Eq. (1):
where [SO42−] is the concentration of sulfate in the sample, t is the incubation time, and 1.06 represents an empirically determined fractionation factor between 35SO42− and 32SO42− that corrects for the slightly slower turnover of 35SO42− 44,77. MDL of the method was estimated from CPM data measured in “killed control” (blank) samples of 35S-sulfate tracer added to samples of each well immediately preserved in zinc acetate solution (20%, v.v), incubated with the regular samples and subjected to the distillation procedure. These blank samples had CPM values of 30 ± 1.4 (1ϭ) and MDLCPM was determined by the mean of n (=25) samples according to MDLCPM = mean(CPM) + t × ϭ, where t is the Student t-factor for n–1 degrees of freedom at 95% confidence. MDLCPM was 33.5 CPM and samples with lower CPM values were reported as below detection. The MDLCPM translates to an MDL of 5 × 10−4 pmol mL−1 d−1 at the incubated amount of radioactivity and 200 µmol L−1 sulfate concentration. Tracer blanks were determined in triplicates by subjecting 3-MBq aliquots of each tracer batch to the separation procedure to determine trace amounts of reduced 35S species in the sulfate tracer and used to correct the sample measurements.
Chemical species analysis
Sulfate analysis was done on a Dionex ICS 2000 ion chromatograph (Thermo Fisher) equipped with a KOH eluent generator on an AS14 analytical column (4 mm). KOH concentration was raised from 20 to 32 mmol L−1 after 15 min of runtime. Hydrogen and methane were measured in the gas phase extracts by gas chromatography using, respectively, a Peak Performer with a reduced compound photometer (Peak Laboratories, USA) and an SRI Model 8610 with flame ionization detector.
Cell abundance
Cell enumeration was performed as previously described15,17. Unfiltered well fluid was mixed with 3.7% formaldehyde at a ratio of 9:1 in the field and then refrigerated. Preserved fluids were filtered through black polycarbonate filters with 0.2 µm pore size (Millipore, Billerica, MA, USA) and stained with 1 µg mL−1 of 4’,6-diamidino-2-phenylindole and counted on an epifluorescence microscope78.
Abundance of SRB
To estimate the percentage of the microbial community that SRB constitute, we manually screened inferred OTU taxonomy from 16S rRNA amplicon sequencing of DNA extracted from fluid biomass for previously published sulfate-reducing organisms (see Supplementary for sequencing and processing methods). Taxa were considered sulfate-reducing if all members of the taxonomic rank an OTU most specifically classified to, as identified by the Genome Taxonomy Database (GTDB release 5), possess dsrAB genes. The relative abundance of OTUs identified as likely sulfate reducers was summed and reported as %SRB (Supplementary Tables 1 and 2). This value represents a likely underestimate as (1) there are taxa detected where only some members contain complete pathways for dissimilatory sulfate reduction and (2) there is growing evidence for the presence of uncultivated lineages that contain sulfate reduction genes.
Data availability
All data generated in this study are available at the PANGAEA database under the collection title: “Activity and abundance of sulfate reducing bacteria and environmental parameter in serpentinizing fluids of Coast Range ophiolite (USA) and Samail ophiolite (Oman)” and are additionally provided as supplementary data files.
References
Orosei, R. et al. Radar evidence of subglacial liquid water on Mars. Science 361, 490–493 (2018).
Hsu, H.-W. et al. Ongoing hydrothermal activities within Enceladus. Nature 519, 207–210 (2015).
Kivelson, M. G. et al. Galileo magnetometer measurements: a stronger case for a subsurface ocean at Europa. Science 289, 1340–1343 (2000).
Parkinson, C. D., Liang, M.-C., Yung, Y. L. & Kirschivnk, J. L. Habitability of Enceladus: planetary conditions for life. Orig. Life Evol. Biospheres 38, 355–369 (2008).
Hand, K. P., Carlson, R. W. & Chyba, C. F. Energy, chemical disequilibrium, and geological constraints on Europa. Astrobiology 7, 1006–1022 (2007).
Neubeck, A., Duc, N. T., Bastviken, D., Crill, P. & Holm, N. G. Formation of H2 and CH4 by weathering of olivine at temperatures between 30 and 70 °C. Geochem. Trans. 12, 6 (2011).
Vance, S. D. & Melwani Daswani, M. Serpentinite and the search for life beyond Earth. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 378, 20180421 (2020).
Eickenbusch, P. et al. Origin of short-chain organic acids in serpentinite mud volcanoes of the Mariana convergent margin. Front. Microbiol. 10, 1729 (2019).
Schrenk, M. O., Brazelton, W. J. & Lang, S. Q. Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 75, 575–606 (2013).
Schulte, M., Blake, D., Hoehler, T. & McCollom, T. Serpentinization and its implications for life on the early Earth and Mars. Astrobiology 6, 364–376 (2006).
Hoefen, T. M. et al. Discovery of Olivine in the Nili Fossae Region of Mars. Science 302, 627–630 (2003).
Ehlmann, B. L., Mustard, J. F. & Murchie, S. L. Geologic setting of serpentine deposits on Mars. Geophys. Res. Lett. 37, L06201 (2010).
Waite, J. H. et al. Cassini finds molecular hydrogen in the Enceladus plume: evidence for hydrothermal processes. Science 356, 155–159 (2017).
Waite, J. H. et al. Cassini ion and neutral mass spectrometer: Enceladus plume composition and structure. Science 311, 1419–1422 (2006).
Brazelton, W. J. et al. Metagenomic identification of active methanogens and methanotrophs in serpentinite springs of the Voltri Massif, Italy. PeerJ 5, e2945 (2017).
Fones, E. M. et al. Physiological adaptations to serpentinization in the Samail Ophiolite, Oman. ISME J 13, 1750–1762 (2019).
Crespo-Medina, M. et al. Methane dynamics in a tropical serpentinizing environment: the Santa Elena Ophiolite, Costa Rica. Front. Microbiol. 8, 916 (2017).
Hoehler, T. M. et al. Comparative ecology of H2 cycling in sedimentary and phototrophic ecosystems. Antonie Van Leeuwenhoek 81, 575–585 (2002).
Hoehler, T. M., Alperin, M. J., Albert, D. B. & Martens, C. S. Thermodynamic control on hydrogen concentrations in anoxic sediments. Geochim. Cosmochim. Acta 62, 1745–1756 (1998).
Canfield, D. E. & Raiswell, R. The evolution of the sulfur cycle. Am. J. Sci. 299, 697–723 (1999).
Ono, S. et al. New insights into Archean sulfur cycle from mass-independent sulfur isotope records from the Hamersley Basin, Australia. Earth Planet. Sci. Lett. 213, 15–30 (2003).
Cameron, E. M. Sulphate and sulphate reduction in early Precambrian oceans. Nature 296, 145–148 (1982).
Jørgensen, B. B. & Fenchel, T. The sulfur cycle of a marine sediment model system. Mar. Biol. 24, 189–201 (1974).
Lovley, D. R. & Klug, M. J. Sulfate reducers can outcompete methanogens at freshwater sulfate concentrations. Appl. Environ. Microbiol. 45, 187–192 (1983).
Glombitza, C., Stockhecke, M., Schubert, C. J., Vetter, A. & Kallmeyer, J. Sulfate reduction controlled by organic matter availability in deep sediment cores from the saline, alkaline Lake Van (Eastern Anatolia, Turkey). Front. Microbiol. 4, 209 (2013).
Holmer, M. & Storkholm, P. Sulphate reduction and sulphur cycling in lake sediments: a review. Freshw. Biol. 46, 431–451 (2001).
Bell, E. et al. Active sulfur cycling in the terrestrial deep subsurface. ISME J. 14, 1260–1272 (2020).
Lin, L.-H. et al. Long-term sustainability of a high-energy, low-diversity crustal biome. Science 314, 479–482 (2006).
Alt, J. C. & Shanks, W. C. Sulfur in serpentinized oceanic peridotites: serpentinization processes and microbial sulfate reduction. J. Geophys. Res. Solid Earth 103, 9917–9929 (1998).
Alt, J. C. et al. The role of serpentinites in cycling of carbon and sulfur: seafloor serpentinization and subduction metamorphism. Lithos 178, 40–54 (2013).
Twing, K. I. et al. Serpentinization-influenced groundwater harbors extremely low diversity microbial communities adapted to high pH. Front. Microbiol. 8, 308 (2017).
Sabuda, M. C. et al. A dynamic microbial sulfur cycle in a serpentinizing continental ophiolite. Environ. Microbiol. 22, 2329–2345 (2020).
Dulov, L., Lein, A., Dubinina, G. & Pimenov, N. Microbial processes at the Lost City Vent Field, Mid-Atlantic Ridge. Microbiology 74, 97–103 (2005).
Murchie, S. L. et al. A synthesis of Martian aqueous mineralogy after 1 Mars year of observations from the Mars Reconnaissance Orbiter. J. Geophys. Res. Planets 114, E00D06 (2009).
Marlow, J. J., LaRowe, D. E., Ehlmann, B. L., Amend, J. P. & Orphan, V. J. The potential for biologically catalyzed anaerobic methane oxidation on ancient Mars. Astrobiology 14, 292–307 (2014).
McCord, T. B. et al. Salts on Europa’s surface detected by Galileo’s near infrared mapping spectrometer. Science 280, 1242–1245 (1998).
Dalton, J. B. Spectral behavior of hydrated sulfate salts: implications for Europa mission spectrometer design. Astrobiology 3, 771–784 (2003).
Brown, M. E. & Hand, K. P. Salts and radiation products on the surface of Europa. Astron. J. 145, 110 (2013).
Kattenhorn, S. A. & Prockter, L. M. Evidence for subduction in the ice shell of Europa. Nat. Geosci. 7, 762–767 (2014).
Zolotov, M. Y. & Shock, E. L. Energy for biologic sulfate reduction in a hydrothermally formed ocean on Europa. J. Geophys. Res. Planets 108, E4, 5022 (2003).
McKinnon, W. B. & Zolensky, M. E. Sulfate content of Europa’s ocean and shell: evolutionary considerations and some geological and astrobiological implications. Astrobiology 3, 879–897 (2003).
Rempfert, K. R. et al. Geological and geochemical controls on subsurface microbial life in the Samail Ophiolite, Oman. Front. Microbiol. 8, 56 (2017).
Kallmeyer, J., Ferdelman, T. G., Weber, A., Fossing, H. & Jørgensen, B. B. A cold chromium distillation procedure for radiolabeled sulfide applied to sulfate reduction measurements. Limnol. Oceanogr. Methods 2, 171–180 (2004).
Røy, H., Weber, H. S., Tarpgaard, I. H., Ferdelman, T. G. & Jørgensen, B. B. Determination of dissimilatory sulfate reduction rates in marine sediment via radioactive 35S tracer. Limnol. Oceanogr. Methods 12, 196–211 (2014).
Glombitza, C. et al. Microbial sulfate reduction potential in coal-bearing sediments down to ∼2.5 km below the seafloor off Shimokita Peninsula, Japan. Front. Microbiol. 7, 1576 (2016).
Foti, M. et al. Diversity, activity, and abundance of sulfate-reducing bacteria in saline and hypersaline soda lakes. Appl. Environ. Microbiol. 73, 2093–2100 (2007).
Jørgensen, B. B., Findlay, A. J. & Pellerin, A. The biogeochemical sulfur cycle of marine sediments. Front. Microbiol. 10, 849 (2019).
Beulig, F., Røy, H., Glombitza, C. & Jørgensen, B. B. Control on rate and pathway of anaerobic organic carbon degradation in the seabed. Proc. Natl. Acad. Sci. USA. 115, 367–372 (2018).
Pellerin, A. et al. The sulfur cycle below the sulfate-methane transition of marine sediments. Geochim. Cosmochim. Acta 239, 74–89 (2018).
Glombitza, C., Egger, M., Røy, H. & Jørgensen, B. B. Controls on volatile fatty acid concentrations in marine sediments (Baltic Sea). Geochim. Cosmochim. Acta 258, 226–241 (2019).
Pikuta, E. V. et al. Desulfonatronum thiodismutans sp. nov., a novel alkaliphilic, sulfate-reducing bacterium capable of lithoautotrophic growth. J. Med. Microbiol. 53, 1327–1332 (2003).
Pérez-Bernal, M. F. et al. Desulfobotulus mexicanus sp. nov., a novel sulfate-reducing bacterium isolated from the sediment of an alkaline crater lake in Mexico. Int. J. Syst. Evol. Microbiol. 70, 3219–3225 (2020).
Oremland, R. S. et al. Bacterial dissimilatory reduction of arsenate and sulfate in meromictic Mono Lake, California. Geochim. Cosmochim. Acta 64, 3073–3084 (2000).
Kulp, T. R. et al. Dissimilatory arsenate and sulfate reduction in sediments of two hypersaline, arsenic-rich soda lakes: Mono and Searles Lakes, California. Appl. Environ. Microbiol. 72, 6514–6526 (2006).
Smith, R. L. & Oremland, R. S. Big Soda Lake (Nevada). 2. Pelagic sulfate reduction. Limnol. Oceanogr. 32, 794–803 (1987).
Fryer, P. et al. Mariana serpentinite mud volcanism exhumes subducted seamount materials: implications for the origin of life. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 378, 20180425 (2020).
D’Hondt, S., Rutherford, S. & Spivack, A. J. Metabolic activity of subsurface life in deep-sea sediments. Science 295, 2067–2070 (2002).
Ravenschlag, K., Sahm, K., Knoblauch, C., Jørgensen, B. B. & Amann, R. Community structure, cellular rRNA content, and activity of sulfate-reducing bacteria in marine Arctic sediments. Appl. Environ. Microbiol. 66, 3592–3602 (2000).
Knoblauch, C., Jørgensen, B. B. & Harder, J. Community size and metabolic rates of psychrophilic sulfate-reducing bacteria in Arctic marine sediments. Appl. Environ. Microbiol. 65, 4230–4233 (1999).
Anantharaman, K. et al. Expanded diversity of microbial groups that shape the dissimilatory sulfur cycle. ISME J. 12, 1715–1728 (2018).
Petro, C. et al. Marine deep biosphere microbial communities assemble in near-surface sediments in Aarhus Bay. Front. Microbiol. 10, 758 (2019).
Jochum, L. M. et al. Depth distribution and assembly of sulfate-reducing microbial communities in marine sediments of Aarhus Bay. Appl. Environ. Microbiol. 83, e01547-17 (2017).
Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biochim. Biophys. Acta BBA Bioenerg. 1807, 1507–1538 (2011).
Mulkidjanian, A. Y., Dibrov, P. & Galperin, M. Y. The past and present of sodium energetics: May the sodium-motive force be with you. Biochim. Biophys. Acta BBA Bioenerg. 1777, 985–992 (2008).
Skulachev, V. P. The sodium cycle: a novel type of bacterial energetics. J. Bioenerg. Biomembr. 21, 635–647 (1989).
Krulwich, T. A. Alkaliphiles:‘basic’ molecular problems of pH tolerance and bioenergetics. Mol. Microbiol. 15, 403–410 (1995).
Froelich, P. N. et al. Early oxidation of organic matter in pelagic sediments of the eastern equatorial Atlantic: suboxic diagenesis. Geochim. Cosmochim. Acta 43, 1075–1090 (1979).
Haveman, S. A. & Pedersen, K. Distribution of culturable microorganisms in Fennoscandian Shield groundwater. FEMS Microbiol. Ecol. 39, 129–137 (2002).
Leloup, J., Quillet, L., Berthe, T. & Petit, F. Diversity of the dsrAB (dissimilatory sulfite reductase) gene sequences retrieved from two contrasting mudflats of the Seine estuary, France. FEMS Microbiol. Ecol. 55, 230–238 (2006).
Xiao, K.-Q., Beulig, F., Røy, H., Jørgensen, B. B. & Risgaard‐Petersen, N. Methylotrophic methanogenesis fuels cryptic methane cycling in marine surface sediment. Limnol. Oceanogr. 63, 1519–1527 (2018).
Cardace, D. et al. Establishment of the Coast Range Ophiolite Microbial Observatory (CROMO): drilling objectives and preliminary outcomes. Sci. Drill. 16, 45–55 (2013).
Shervais, J. W., Murchey, B. L., Kimbrough, D. L., Renne, P. R. & Hanan, B. Radioisotopic and biostratigraphic age relations in the Coast Range Ophiolite, northern California: implications for the tectonic evolution of the Western Cordillera. GSA Bull. 117, 633–653 (2005).
Peters, E. K. D-18O enriched waters of the Coast Range Mountains, northern California: connate and ore-forming fluids. Geochim. Cosmochim. Acta 57, 1093–1104 (1993).
Barnes, I., O’Neil, J. R. & Trescases, J. J. Present day serpentinization in New Caledonia, Oman and Yugoslavia. Geochim. Cosmochim. Acta 42, 144–145 (1978).
Neal, C. & Stanger, G. Hydrogen generation from mantle source rocks in Oman. Earth Planet. Sci. Lett. 66, 315–320 (1983).
Kelemen, P. et al. Scientific drilling and related research in the samail ophiolite. Sultanate of Oman. Sci. Drill. 15, 64–71 (2013).
Jørgensen, B. B. A comparison of methods for the quantification of bacterial sulfate reduction in coastal marine sediments. Geomicrobiol. J. 1, 11–27 (1978).
Schrenk, M. O., Kelley, D. S., Delaney, J. R. & Baross, J. A. Incidence and diversity of microorganisms within the walls of an active deep-sea sulfide chimney. Appl. Environ. Microbiol. 69, 3580–3592 (2003).
Acknowledgements
This study was supported by funding from the NASA Astrobiology Institute to the Rock Powered Life Team (NNA15BB02A) and the NASA Habitable Worlds program (17-HW17_2-0036). C.G. was funded by the NASA Postdoctoral Program (NPP). We thank Juerg Matter and the Oman Ministry of Regional Municipalities and Water Resources for help with accessing the field sites in Oman and permitting sample export and Cathy Koehler and Paul Aigner for providing access to and maintaining the McLaughlin Natural Reserve and field station. We also thank Dawn Cardace, Emily Kraus, Elizabeth Fones, Daniel Nothaft, Eric Boyd, Laura Bueter, and Eric Ellison for help with sample collection, and Emily Kraus for extraction and library preparation of Samail Ophiolite DNA. We thank Esther Schwarzenbach and Tim Ferdelman for their constructive reviews.
Author information
Authors and Affiliations
Contributions
C.G. and T.M.H. deigned the study; A.S.T., C.G., K.R.R., L.I.P. and M.D.K. collected the samples; C.G. measured sulfate reduction rates; M.D.K. measured H2 and sulfate concentrations; K.R.R., L.I.P. and M.O.S. analyzed DNA; C.G. wrote the manuscript; A.S.T., M.O.S. and T.M.H. edited the manuscript. All co-authors critically evaluated the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Primary handling editor: Joe Aslin.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Glombitza, C., Putman, L.I., Rempfert, K.R. et al. Active microbial sulfate reduction in fluids of serpentinizing peridotites of the continental subsurface. Commun Earth Environ 2, 84 (2021). https://doi.org/10.1038/s43247-021-00157-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-021-00157-z
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
